Effects of different flui replacements on serum HSP70 and lymphocyte DNA damage in college athletes during exercise at high ambient temperatures
2016-02-05HeeTaeRohSuYounChoWiYoungSoIlYoungPaikSangHoonSuh
Hee-Tae Roh,Su-Youn Cho,Wi-Young So,Il-Young Paik*,Sang-Hoon Suh*
aDepartment of Physical Education,Dong-A University,Busan 604-714,Republic of Korea
bExercise Physiology Laboratory,Department of Physical Education,Yonsei University,Seoul 120-749,Republic of Korea
cSports and Health Care Major,College of Humanities and Arts,Korea National University of Transportation,Chungju-si 380-702,Republic of Korea
Effects of different flui replacements on serum HSP70 and lymphocyte DNA damage in college athletes during exercise at high ambient temperatures
Hee-Tae Roha,†,Su-Youn Chob,†,Wi-Young Soc,Il-Young Paikb,*,Sang-Hoon Suhb,*
aDepartment of Physical Education,Dong-A University,Busan 604-714,Republic of Korea
bExercise Physiology Laboratory,Department of Physical Education,Yonsei University,Seoul 120-749,Republic of Korea
cSports and Health Care Major,College of Humanities and Arts,Korea National University of Transportation,Chungju-si 380-702,Republic of Korea
Purpose:The aim of this study was to investigate the effects of flui replacement by water or sports drinks on serum heat shock protein 70 (HSP70)levels and DNA damage during exercise at a high ambient temperature.
Methods:Ten male college athletes with an athletic career ranging from 6 to 11 years were recruited from Yonsei University.The subjects ran on a treadmill at 75%of heart rate reserve during 4 different trials:thermoneutral temperature at 18°C(T),high ambient temperature at 32°C without flui replacement(H),high ambient temperature at 32°C with water replacement(HW),and high ambient temperature at 32°C with sports drink replacement(HS).During each condition,blood samples were collected at the pre-exercise baseline(PEB),immediately after exercise(IAE),and 60 min post-exercise.
Results:Skin temperature significant y increased during exercise and was significant y higher in H compared to T and HS at IAE.Meanwhile, serum HSP70 was significant y increased in all conditions at IAE compared to PEB and was higher in H compared to T at the former time point. Significant y increased lymphocyte DNA damage(DNA in the tail,tail length,tail moment)was observed in all trials at IAE compared to PEB, and attenuated DNA damage(tail moment)was observed in HS compared to H at IAE.
Conclusion:Acute exercise elevates serum HSP70 and induces lymphocyte DNA damage.Fluid replacement by sports drink during exercise at high ambient temperature can attenuate HSP response and DNA damage by preventing dehydration and reducing thermal stress.
©2016 Production and hosting by Elsevier B.V.on behalf of Shanghai University of Sport.This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
Acute exercise;DNA damage;Fluid replacement;Heat stress;HSP70;Sports drink
1.Introduction
Increased external temperature can exert an influenc on cardiovascular,hormonal,and immune responses,and can also negatively affect performance during endurance exercise.1Hyperthermia and dehydration are also considered major limiting factors of exercise performance in the heat.2Previous studies have suggested that hyperthermia and dehydration,together with oxidative stress,are associated with heat-related exercise response,1,3and that the production of free radicals resulting from exercise and/or heat stress is the cause of leukocyte DNA damage after exercise.4,5DNA damage due to severe exercise stress in turn can lead to apoptosis and could partly account for exercise-related lymphopenia.6
The induction of heat shock proteins(HSPs)may function in leukocytes as a protective mechanism against exercise,heat,and oxidative stress.1Fehrenbach et al.1suggested that a cause-andeffect relation was present between exercise-induced oxidative stress and HSP response.The protective functions of HSPs include the protection of cellular homeostasis and immune function through chaperoning protein assembly,degradation,translocation,and stabilization.7HSPs can be classifie according to molecular weight(kDa)into the categories of 10 kDa,20–30 kDa,40 kDa,60 kDa,70 kDa,90 kDa,and 100 kDa proteins.8Of these,HSP70 showed the greatest variation in expression in response to exercise stimuli.8In addition,HSP70 has been found to play a crucial role in thermotolerance9and to be induced rapidly by physiological stress factors such as pH changes in the body or glycogen depletion.10Furthermore, HSP70 expression in response to stress has been shown to play an important protective role against apoptosis11and in the regulation of apoptotic cell signaling.12Nuclear translocation of HSP72,a member of the HSP70kDa family,was also reported to suppress the occurrence of apoptosis in DNA-damaged cells.13
On the other hand,flui replacement during exercise in the heat could attenuate the rise in body temperature.14Additionally,a previous study by Hillman et al.3suggested that exerciseinduced dehydration could increase oxidative stress but that flui replacement could decrease thermal and oxidative stress during prolonged exercise in the heat.Sports drinks have also been reported to be more effective than water in flui replacement for attenuating the increase in oxidative stress caused by dehydration.15However,to date,noin vivostudy has assessed the heat shock response and DNA damage in leukocytes in a comparison between different flui replacements.
Therefore,the present study aimed to investigate the effects of flui replacement by water or sports drinks on serum HSP70 and lymphocyte DNA damage during exercise at a high ambient temperature among college athletes.
2.Methods
2.1.Subjects
Ten male college athletes(soccer,n=5;rugby,n=5)with athletic careers ranging from 6 to 11 years were recruited from Yonsei University.Subjects were non-smoking,non-drinking, and injury-and disease-free as determined by a health history questionnaire and physical examination.The baseline physical characteristics of the subjects are shown in Table 1.All subjects provided informed consent,and the study protocol wasapproved by the Institutional Ethics Review Board of the Department of Physical Education at Yonsei University.
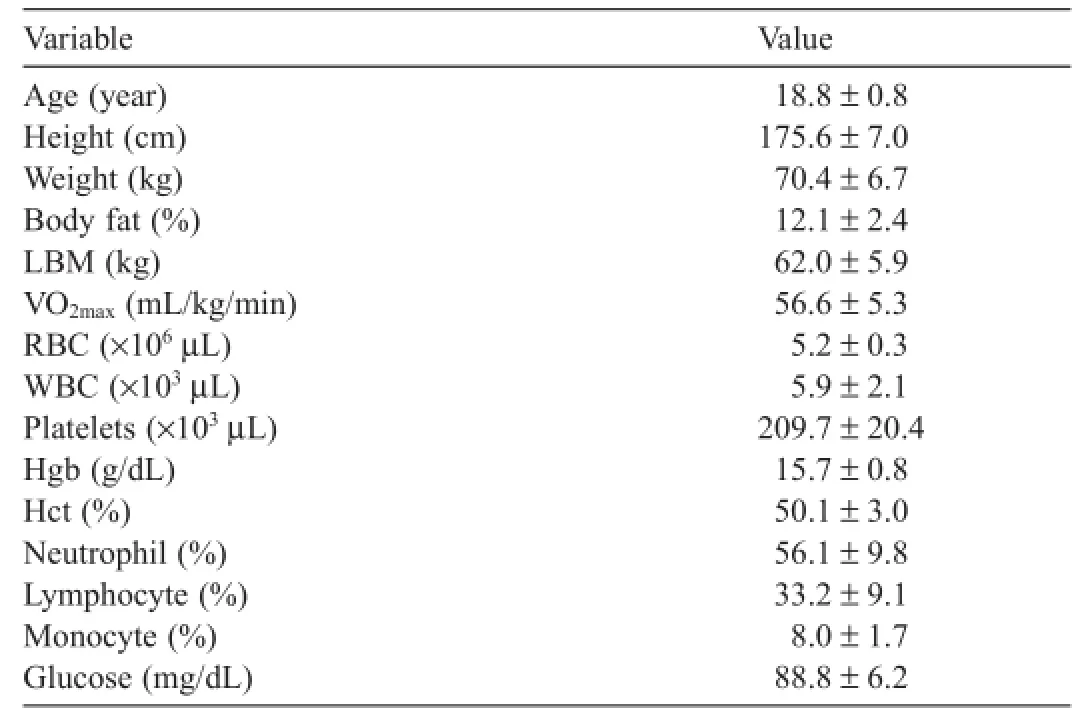
Table 1 Physical characteristics of the subjects at baseline(n=10,mean±SD).
2.2.Preliminary tests
Preliminary tests included measurements of height,body composition(body weight,fat mass,and fat free mass), maximal exercising capacities(maximal oxygen consumption (VO2max),maximal heart rate(HRmax))and hematologic parameters.Height was measured with a stadiometer(HD;STDK, Tokyo,Japan),and body composition was measured through bio-impedance body composition analysis with Inbody220 (Biospace,Seoul,Korea).The VO2maxmeasurement was performed on a treadmill(Q65;Quinton,Seattle,WA,USA) according to the Bruce protocol,in which an increase of 2%in the incline and 0.8 mph in the speed occurs every 3 min from an initial incline level of 10%and a speed of 1.7 mph.16Gas analyses were performed with MetaMax 3B(Cortex,Leipzig, Germany)every 10 s to determine the volume of single ventilation,oxygen consumption,carbon dioxide emission,respiratory exchange ratio,and breathing rate per minute.HRmaxwas measured during the VO2maxmeasurement with Polar a5(Polar, Kempele,Finland).The following hematologic parameters were accessed using an automatic hematology analyzer (Sysmex XE-2100;Sysmex,Kobe,Japan):red blood cell count, white blood cell count,platelet count,hemoglobin concentration,hematocrit value,and neutrophil,lymphocyte,and monocyte percentages.Serum glucose concentration was determined by a hexokinase method using a commercially available glucose/hexokinase assay kit(Pointe Scientific Canton,MI, USA).The absorbance was measured at 340 nm with an automatic analyzer(Hitachi,Tokyo,Japan).
2.3.General experimental design
The exercise intensity for the trials was set at 75%of heart rate reserve for each subject based on data obtained from screening tests.The subjects ran under 4 different conditions:a thermoneutral temperature at 18°C(T),high ambient temperature at 32°C without flui replacement(H),high ambient temperature at 32°C with water replacement(HW),and high ambient temperature at 32°C with sports drink replacement (HS).The exercise intensity and humidity(50%)were identical in all trials.The trials were performed in the order of trial T,H, HW,and HS,and were each separated by 7 days to avoid any transient effects on the physiological and psychological conditions of the subjects.
The experimental condition of 18°C and 50%relative humidity is considered to be thermoneutral for physical activity and to be a comfort zone,since a low heat stress index has been reported for this condition.17In contrast,the condition at 32°C and 50%relative humidity is considered to be adverse because it frequently results in heat cramps.Based on the wet-bulb globe temperature index,sports events conducted under this condition are subject to cancellation.17–19Both experimental conditions(a thermoneutral environment and a high ambient temperature)were set up using an E series walk-in temperature and humidity chamber(ESPEC,Osaka,Japan).
Skin temperature was calculated using the equation introduced by Watson et al.20based on the data collected from the chest,forearm,thigh,and calf with MSR12(MSR Electronics GmbH,Henggart,Switzerland).The measurement timings were 8 times during rest,every 10 min during the exercise trial,immediately after the exercise trial,and 60 min post-exercise.Dehydration levels were calculated by the difference in the body weight between pre-exercise and immediately after the trial in the H group.An amount of plain water(Evian,Danone,France)or sports drink(Gatorade;Pepsi Co.,Inc.,Barrington,IL,USA) equal to the body weight loss after trial H was given to the subjects for flui replacement,with multiple separations during exercise.Plain water and sports drink were prepared at 15°C,the temperature suggested providing the best absorption rate and sensation of refreshment.21In terms of nutritional information, the sports drink used in the current study contained 6%carbohydrate,20.9 mEq/L Na+,6.1 mEq/L K+,and 9.5 mEq/L Cl-.
2.4.Blood sampling and analyses
Blood samples were collected at the pre-exercise baseline (PEB),immediately after the exercise(IAE),and 60 min postexercise(60MPE)from the antecubital vein.For each of the blood sampling time points,serum pH levels,plasma lactate, HSP70,and lymphocyte DNA damage were determined.
2.4.1.Serum pH and plasma lactate analyses
Serum pH was detected with a Corning pH/Ion analyzer model 355(Corning Inc.,Corning,NY,USA).For analyzing the plasma concentration of lactate,potassium oxalate/sodium fluorid was added to the blood samples.The blood samples were subsequently centrifuged at 3000 rpm for 20 min,and the plasma was extracted for analysis.Ten microliters of the collected plasma samples was pipetted on an LAC DT slide using a DT pipette(Eastman Kodak,Rochester,NY,USA)with a microtip(Ortho-Clinical Diagnostics,Rochester,NY,USA). The absorbance was measured at 555 nm for detecting lactate using an Ektachem DT 60 clinical chemistry analyzer(Eastman Kodak).
2.4.2.Serum HSP70 analysis
The serum HSP70 concentrations were determined by an enzyme-linked immunosorbent assay(EIA)using a commercially available HSP70 high-sensitivity EIA kit(Assay Designs, Ann Arbor,MI,USA).The absorbance was measured at 450 nm with a microplate reader(Molecular Device,PaloAlto, CA,USA).
2.4.3.Lymphocyte DNA damage analyses
DNA damage was analyzed as described by Singh et al.22and Green et al.23For the Comet assay,whole blood was mixed with phosphate-buffered saline and poured gently over a peripheral blood lymphocyte separation solution(Histopaque-1077;Sigma Chemical Co.,St.Louis,MO,USA).After centrifugation at 1450 rpm for 25 min,peripheral blood lymphocytes were transferred into another tube,mixed with 0.7%low-melting agarose,and added to slides precoated with 0.5%agarose.The cells were then lysed for 1 h at 4°C in a lysis buffer containing 2.5 mol/L NaCl,100 mmol/L EDTA,1% Triton X-100,10%DMSO,and 10 mmol/L Tris,pH 10.After the lysis,the DNA was allowed to unwind for 40 min in an electrophoretic solution containing 300 mmol/L NaOH and 1 mmol/L EDTA(pH>13).Electrophoresis was carried out at 30 V for 30 min.The slides were then neutralized with 0.4 mol/LTris,pH 7.5,and fi ed with ethanol.Finally,the DNA was stained by adding 60 μL of ethidium bromide(20 μg/mL) to each slide.Measurements were made using image analysis software(Komet 5.0;Kinetic Imaging,Liverpool,UK)and a fluorescenc microscope(Leica,Wetzlar,Germany)equipped with an excitation filte of 515–560 nm and a barrier filte of 590 nm.Images from 50 cells were analyzed for each slide,and the parameters recorded were DNA in the tail(%),tail length (μm),and tail moment(%of DNA in tail×tail length).All steps were performed under dimmed light,and the electrophoresis tank was covered with black paper to avoid additional light-induced DNA damage.
2.5.Statistical analyses
Statistical analyses were performed with SPSS Version 17.0 for Windows(SPSS Inc.,Chicago,IL,USA).Data are presented as mean±SD unless otherwise stated.For identifying differences in normally distributed results,two-way repeated analysis of variance(ANOVA)was employed.When a signifi cant interaction was apparent,the simple main effects on measured variables were determined using one-way ANOVA. Tukey’spost hoctest was subsequently used to conservatively locate significan differences.Statistical significanc was set atp<0.05.
3.Results
3.1.Skin temperature
In trial T,the skin temperature was significant y increased from Ex.20 min(20 min of exercise)to IAE relative to that at Ex.10 min(p<0.05;Fig.1).Additionally,trials H,HW,and HS showed significant y increased temperatures from Ex.10 min to IAE in comparison to PEB(p<0.05).Further,the skin temperature was significant y higher from Ex.10 min to IAE in trials H,HW,and HS as compared to that in trial T(p<0.05). At IAE,the skin temperature was significant y higher in trial H compared to that of trials T and HS(p<0.05).However,there was no significan between trials H and HW(p>0.05).
3.2.Serum pH level and plasma lactate
The pH and plasma lactate levels were significant y decreased and increased,respectively,at IAE relative to those at PEB in all trials(p<0.05;Fig.2).Further,the pH levels were significant y lower at IAE in trial H as compared to trials T and HS(p<0.05);however,there was no significan difference between trials H and HW(p>0.05),while no significan differences in plasma lactate were observed between the trials at any of the blood sampling points.
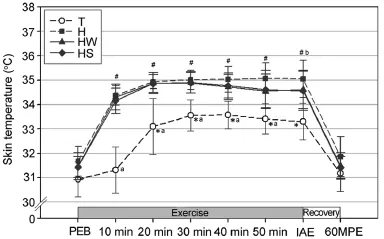
Fig.1.Changes in skin temperature(mean±SD).*Significant y different from the skin temperature at 10 min of exercise in T(p<0.05).#Significant y different from the skin temperature at PEB in H,HW,and HS(p<0.05).aSignificant y lower in T than H,HW,and HS(p<0.05).bSignificant y higher in H than T and HS(p<0.05).H=exercise at high ambient temperature; HS=exercise at high ambient temperature with flui replacement by sports drink;HW=exercise at high ambient temperature with flui replacement by water;IAE=immediately after exercise;PEB=pre-exercise baseline; T=exercise in a thermoneutral environment;60MPE=60 min post-exercise.
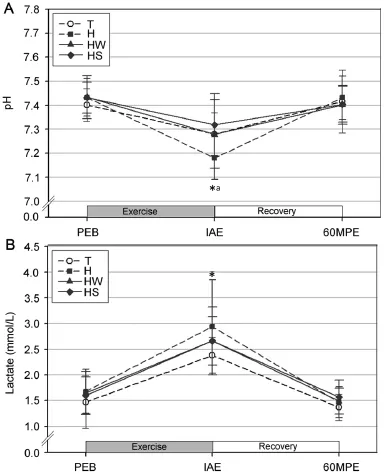
Fig.2.Changes in the serum pH level(A)and plasma lactate(B)(mean±SD). *Significant y different from PEB in all trials(p<0.05).aSignificant y lower in H than T and HS(p<0.05).H=exercise at high ambient temperature; HS=exercise at high ambient temperature with flui replacement by sports drink;HW=exercise at high ambient temperature with flui replacement by water;IAE=immediately afterexercise;PEB=pre-exercise baseline; T=exercise in a thermoneutral environment;60MPE=60 min post-exercise.
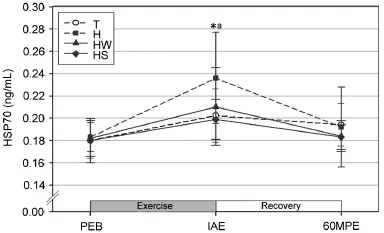
Fig.3.Changes in serum HSP70(mean±SD).*Significant y different from PEB in all trials(p<0.05).aSignificant y higher in H than HS(p<0.05). H=exercise at high ambient temperature;HS=exercise at high ambient temperature with flui replacement by sports drink;HW=exercise at high ambient temperature with flui replacement by water;IAE=immediately after exercise;PEB=pre-exercise baseline;T=exercise in a thermoneutral environment;60MPE=60 min post-exercise.
3.3.Serum HSP70
The serum HSP70 levels were significant y increased at IAE as compared to PEB in all trials(p<0.05;Fig.3).In addition, the serum HSP70 at IAE was significant y higher in trial H as compared to trial HS(p<0.05).
3.4.Lymphocyte DNA damage
Significant y increased DNA in the tail(Fig.4A),tail length (Fig.4B),and tail moment(Fig.4C)relative to the levels at rest were observed at IAE for all trials(p<0.05).Further,the tail moment was significant y higher in trial H at IAE as compared to trials T and HS(p<0.05);there was no significan difference between trials H and HW(p>0.05).However,no significan differences in DNA in the tail and tail length were observed between the trials.
4.Discussion
4.1.Skin temperature
In the present study,the skin temperature was significant y increased after exercise,with trials H,HW,and HS showing significant y higher values than trial T.These results are consistent with previous studies reporting that exercise at high ambient temperature and humidity as compared to a thermoneutral condition results in impaired heat-control capacity,thereby leading to higher body temperature and increased body flui loss.24,25We assume that the high ambient temperature condition(32°C,50%) set for the current study impaired heat control capacity and thus lead to impaired homeostasis of body temperature.In addition, the skin temperature appeared to be significant y lower immediately after exercise in trial HS compared to trial H.These finding support the results of a previous study which showed that a rise in body temperature caused by prolonged exercise could be attenuated more effectively by sports drinks containing carbohydratesand electrolytes than by water,at the time of flui replacement during exercise.26
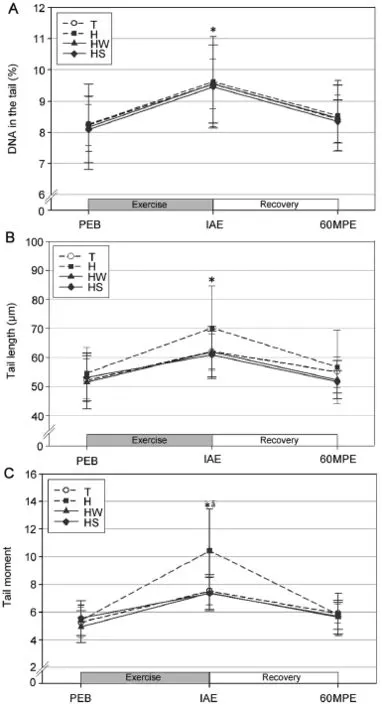
Fig.4.Changes in DNA in the tail(A),tail length(B),and tail moment(C) (mean±SD).*Significant y different from PEB in all trials(p<0.05).aSignificant y higher in H than T and HS(p<0.05).H=exercise at high ambient temperature;HS=exercise at high ambient temperature with flui replacement by sports drink;HW=exercise at high ambient temperature with flui replacement by water;IAE=immediately after exercise; PEB=pre-exercise baseline;T=exercise in a thermoneutral environment; 60MPE=60 min post-exercise.
Glucose administered with flui during exercise was previously reported to spare liver glycogen storage and to reduce the rate of gluconeogenesis,resulting in~6%reduction in heat production within the liver.27,28Moreover,the study of Shi and Gisolf29showed that the combination of increased osmotic pressure and unbalanced sodium levels results in a rise in body temperature,and that a drink containing glucose and electrolytes(e.g.,sodium)can attenuate an increase in osmotic pressure and maintain body flui volume by balancing electrolyte levels.Based on these previous findings the relatively attenuated rise in body temperatures in trial HS as compared to the other trials in the current study can be attributed to the consumption of glucose and electrolytes contained in the sports drink.
4.2.Serum pH and plasma lactate
Generally accepted physiological variables that impair exercising capacity include pH and lactate levels.30,31Lactate does not directly impair physiological functions;however,the increased hydrogen ion(H+)levels that occur with lactate accumulation contribute to acidification which can disrupt enzymatic activities associated with ATP production.Hence,the increased acidificatio can eventually induce fatigue and impair a number of physiological functions.30,32
In the current study,the pH was significant y decreased at IAE relative to PEB in all trials but was restored at 60MPE.The decreased pH induced by exercise eventually results from an accumulation of H+due to accelerated glycolysis.This decreased pH impairs enzymatic activities and cross-bridge formation during muscle contraction,resulting in the early onset of fatigue.At IAE,the pH in trial H was significant y lower than that in trials T and HS.Abbiss et al.33showed a linear inverse coefficien(r=-0.69)between core temperature and pH levels based on experiments involving 3 different conditions:hot(34°C),neutral(22°C),and cold(10°C).Consistent with this previous report,the current study showed that the lowest pH level after exercise occurred under the condition that induced the highest rise in body temperature.
An inefficien oxygen supply to the working muscles is well known to limit oxidative energy production and to increase the dependency on glycolysis,a shift that can result in high lactate levels.34,35In the present study,the plasma lactate levels were significant y increased at IAE in all trials relative to PEB and had decreased to baseline levels at 60MPE.Although no significan difference between the trials was observed at IAE, plasma lactate levels were highest to lowest in the order of trials H,HW,HS,and T.These results are consistent with the previous study of Hargreaves et al.36which reported that during submaximal exercise,a non-hydration group had higher glycogen usage for energy production compared to a hydration group.Furthermore,increased blood viscosity due to decreased blood plasma results in higher cardiac stress,leading to the impaired clearance of metabolic by-products,including lactate. In the current study,trials HW and HS had relatively lower plasma lactate than trial H,and thus support the finding of Ali et al.37In this previous study,fluireplacement during highintensity exercise prevented excessive plasma loss,attenuated cardiac stress,and prevented a substantial decrease in oxygen supply to working muscles,which resulted in a lower production of plasma lactate.Given that lactate production is known to increase with every rise in body temperature,37,38flui replacement during exercise is assumed to contribute to lower plasmalactate and delay fatigue through attenuating an elevation in body temperature.
Previous studies have reported that the administration of fast-absorptive drink containing electrolytes restores oxygen supply to the working muscles by increasing cardiac output and subsequently the peripheral blood supply,as well as delaying acidification39,40Therefore,these previous studies and the current one support that administering sports drink during dehydrating conditions is more effective in attenuating acidifi cation and delaying the onset of fatigue than administering water.
4.3.Serum HSP70
In the present study,serum HSP70 appeared to be signifi cantly higher at IAE in all trials compared to PEB,consistent with the finding of previous studies that acute exercise increases serum HSP70.41,42According to Puntschart et al.,4230 min of treadmill running significant y increased serum HSP70 within skeletal muscle cells.Meanwhile,Suzuki et al.41reported significant y increased serum HSP70 after a triathlon race.Of interest in the current study,the serum HSP70 in trial HS appeared to be significant y lower than that in trial H. According to Sandström et al.,43monocyte serum HSP70 is positively associated with body temperature with a moderate coefficien(r=0.44).Moreover,Lovell et al.44reported higher serum HSP70 within peripheral blood mononuclear cells when heat shock was administered and suggested that serum HSP70 is temperature dependent.Given these reports,the lower serum HSP70 at IAE in trial HS as compared to trial H of the current study is likely attributable to the relatively lower rise in body temperature.Other studies support this discussion that HSP70 expression is induced during increasing oxidative stress.9An increase in oxidative stress by exercise-induced dehydration was determined to be independent of environmental conditions (e.g.,atmospheric temperature)and was able to be attenuated through the maintenance of euhydration by flui replacement during exercise.4A previous study has reported that after excessive dehydration(3%body weight),the intake of sports drinks rather than water was more effective for flui replacement in the attenuation of oxidative stress.15In addition,serum HSP70 was reported to be influence not only by thermal stress but also by pH45and glucose levels.46Taking these results together,the lower serum HSP70 in trial HS as compared to trial H can be attributed to rehydration attenuating a rise in body temperature and oxidative stress and to electrolytes and glucose in the sports drink possibly delaying the acidificatio of working muscles and increasing the efficien y of energy utilization.
4.4.Lymphocyte DNA damage
All 3 parameters for lymphocyte DNA damage(DNA in the tail,tail length,and tail moment)were significant y increased at IAE compared to PEB in all trials.Based on previous reports,the lymphocyte DNA damage likely resulted from the increased production of reactive oxygen species (ROS),particularly by the superoxide radicals generated by the high oxygen supply during high-intensity exercise.47,48Previous studies have reported that acute exercise increases the parameters for oxidative stress.49–51Moreover,some studies have reported that increased DNA damage,as assessed by the Comet assay,follows acute exercise,15,52,53supporting the results of the current study.In particular,Paik et al.15reported increased DNA in the tail,DNA tail length,and DNA tail moment in lymphocytes at the completion of high-intensity treadmill running at80% VO2maxuntil exhaustion.Meanwhile, Mastaloudis et al.53reported a significant y increased DNA in the tail of leukocytes during a 50 km ultra-marathon for 7 h.In addition,Hartmann et al.52reported a significant y increased tail moment at the completion of a triathlon.
In the current study,the tail moment in trial H was significantly higher than that in trials T and HS at IAE.Pereira Ede et al.54suggested that dehydration is the most deleterious factor for oxidative stress based on their study results,which showed a 10-fold increase in oxidative stress after dehydration in yeast cells.Zuo et al.55showed higher ROS production in the diaphragm of rats under heat stress conditions(42°C)as compared to the control condition(37°C).In this previous study,a greater elevation in the core temperature was observed under the heat stress condition,suggesting a potential association between core temperature and ROS production.Given the finding of these previous reports,the elevated body temperatures and the subsequent dehydration possibly contributed to the increased lymphocyte DNA damage in the present study.By contrast,the flui replacement by water or sports drinks may have attenuated lymphocyte DNA damage by reducing the 2 stressors of dehydration and exercise.Sports drinks were reported to be 4-fold more efficien in preserving plasma volume as compared to water,since sports drinks control osmotic pressure by replenishing electrolyte loss and enhancing the re-absorption of electrolytes by supplying glucose.Glucose functions by acting as a co-carrier for water and sodium in the proximal tubule of the glomerulus.56,57Therefore,in the current study,attenuated DNA damage with flui replacement by sports drinks during exercise in high ambient temperature is thought to be the result of a reduction in ROS production through the prevention of hypoxia and elevated body temperatures during exercise.
5.Conclusion
These results suggest that(a)acute exercise elevates serum HSP70 and induces lymphocyte DNA damage;and(b)flui replacement by sports drinks during exercise at high ambient temperature can attenuate HSP response and DNA damage by preventing dehydration and reducing thermal stress.
Acknowledgment
This work was supported by the Dong-A University research fund.This article is a partly condensed form of the firs author’s doctoral thesis(Dr.Hee-Tae Roh)from Yonsei University.
Authors’contributions
HTR and SYC participated in study design,subject recruitment,data collection,data processing,data analysis,anddrafted the manuscript;WYS participated in data analysis,and drafted the manuscript;IYP and SHS conceived of the study, and participated in its design and coordination and helped to draft the manuscript.All authors have read and approved the fina version of the manuscript,and agree with the order of presentation of the authors.
Competing interests
None of the authors declare competing financia interests.
1.Fehrenbach E,Veith R,Schmid M,Dickhuth HH,Northoff H,Niess AM. Inverse response of leukocyte heat shock proteins and DNA damage to exercise and heat.Free Radic Res2003;37:975–82.
2.Cheuvront SN,KeneficRW,Montain SJ,Sawka MN.Mechanisms of aerobic performance impairment with heat stress and dehydration.J Appl Physiol2010;109:1989–95.
3.Hillman AR,Vince RV,Taylor L,McNaughton L,Mitchell N,Siegler J. Exercise-induced dehydration with and without environmental heat stress results in increased oxidative stress.Appl Physiol Nutr Metab2011;36:698–706.
4.Niess AM,Baumann M,Roecker K,Horstmann T,Mayer F,Dickhuth HH. Effects of intensive endurance exercise on DNA damage in leukocytes.J Sports Med Phys Fitness1997;38:111–5.
5.Niess AM,Dickhuth HH,Northoff H,Fehrenbach E.Free radicals and oxidative stress in exercise-immunological aspects.Exerc Immunol Rev1999;5:22–56.
6.Mars M,Govender S,Weston A,Naicker V,Chuturgoon A.High intensity exercise:a cause of lymphocyte apoptosis?Biochem Biophys Res Commun1998;249:366–70.
7.Morimoto RI,Tissieres A,Georgopoulos C.Progress and perspectives on the biology of heat shock proteins and molecular chaperones.In:Morimoto RI,Tissieres A,Georgopoulos C,editors.The biology of heat shock proteins and molecular chaperones.New York,NY:Cold Spring Harbor Laboratory Press;1994.p.1–30.
8.Noble EG,Milne KJ,Melling CW.Heat shock proteins and exercise:a primer.Appl Physiol Nutr Metab2008;33:1050–65.
9.Piper P.Induction of heat shock proteins and thermotolerance.Methods Mol Biol1996;53:313–7.
10.Morton JP,Holloway K,Woods P,Cable NT,Burniston J,Evans L,et al. Exercise training-induced gender-specifi heat shock protein adaptations in human skeletal muscle.Muscle Nerve2009;39:230–3.
11.Siu PM,Bryner RW,Martyn JK,Alway SE.Apoptotic adaptations from exercise training in skeletal and cardiac muscles.FASEB J2004;18:1150–2.
12.Creagh EM,Sheehan D,Cotter TG.Heat shock proteins—modulators of apoptosis in tumour cells.Leukemia2000;14:1161–73.
13.Kato K,Yamanaka K,Hasegawa A,Okada S.Dimethylarsinic acid exposure causes accumulation of Hsp72 in cell nuclei and suppresses apoptosis in human alveolar cultured(L-132)cells.Biol Pharm Bull1999;22:1185–8.
14.Horswill CA.Effective fluireplacement.Int J Sport Nutr1998;8:175–95.
15.Paik IY,Jeong MH,Jin HE,Kim YI,Suh AR,Cho SY,et al.Fluid replacement following dehydration reduces oxidative stress during recovery.Biochem Biophys Res Commun2009;383:103–7.
16.Bruce RA,Blackmon JR,Jones JW,Strait G.Exercising testing in adult normal subjects and cardiac patients.Pediatrics1963;32:742–56.
17.McArdle WD,Katch FI,Katch VL.Essentials of exercise physiology.3rd ed.Philadelphia,PA:Lippincott Williams&Wilkins;2005.
18.American College of Sports Medicine.Prevention of thermal injuries during distance running.Med J Aust1984;141:876–9.
19.American College of Sports Medicine;Armstrong LE,Casa DJ, Millard-Stafford M,Moran DS,Pyne SW,et al.American College of Sports Medicine position stand.Exertional heat illness during training and competition.Med Sci Sports Exerc2007;39:556–72.
20.Watson P,Shirreffs SM,Maughan RJ.Blood-brain barrier integrity may be threatened by exercise in a warm environment.Am J Physiol Regul Integr Comp Physiol2005;288:1689–94.
21.American College of Sports Medicine;Sawka MN,Burke LM,Eichner ER,Maughan RJ,Montain SJ,et al.American College of Sports Medicine position stand.Exercise and flui replacement.Med Sci Sports Exerc2007;39:377–90.
22.Singh NP,McCoy MT,Tice RR,Schneider EL.A simple technique for quantitation of low levels of DNA damage in individual cells.Exp Cell Res1988;175:184–91.
23.Green MH,Lowe JE,Harcourt SA,Akinluyi P,Rowe T,Cole J,et al.UV-C sensitivity of unstimulated and stimulated human lymphocytes from normal and xeroderma pigmentosum donors in the comet assay:a potential diagnostic technique.Mutat Res1992;273:137–44.
24.Powers SK,Howley ET,Cox R.Blood lactate concentrations during submaximal work under differing environmental conditions.J Sports Med Phys Fitness1985;25:84–9.
25.Rasch W,Samson P,Cote J,Cabanac M.Heat loss from the human head during exercise.J Appl Physiol1991;71:590–5.
26.Bergeron MF,Waller JL,Marinik EL.Voluntary flui intake and core temperature responses in adolescent tennis players:sports beverage versus water.Br J Sports Med2006;40:406–10.
27.Hamilton MT,Gonzalez-Alonso J,Montain SJ,Coyle EF.Fluid replacement and glucose infusion during exercise prevent cardiovascular drift.J Appl Physiol1991;71:871–7.
28.Nielsen B,Savard G,Richter EA,Hargreaves M,Saltin B.Muscle blood fl w and muscle metabolism during exercise and heat stress.JAppl Physiol1990;69:1040–6.
29.Shi X,Gisolf CV.Fluid and carbohydrate replacement during intermittent exercise.Sports Med1998;25:157–72.
30.Finsterer J.Biomarkers of peripheral muscle fatigue during exercise.BMC Musculoskelet Disord2012;13:218.doi:10.1186/1471-2474-13-218
31.Juel C.Changes in interstitial K+and pH during exercise:implications for blood fl w regulation.Appl Physiol Nutr Metab2007;32:846–51.
32.Parkhouse WS.The effects of ATP,inorganic phosphate,protons,and lactate on isolated myofibrilla ATPase activity.Can J Physiol Pharmacol1992;70:1175–81.
33.Abbiss CR, Nosaka K, Laursen PB. Hyperthermic-induced hyperventilation and associated respiratory alkalosis in humans.Eur JAppl Physiol2007;100:63–9.
34.Chwalbinski-Moneta J, Krysztofia H, Ziemba A, Nazar K, Kaciuba-Uscilko H.Threshold increases in plasma hormone in relation to plasma catecholamine and blood lactate concentrations during progressive exercise in endurance-trained athletes.Eur J Appl Physiol Occup Physiol1996;73:117–20.
35.Moquin A,Mazzeo RS.Effect of mild dehydration on the lactate threshold in women.Med Sci Sports Exerc2000;32:396–402.
36.Hargreaves M,Dillo P,Angus D,Febbraio M.Effect of flui ingestion on muscle metabolism during prolonged exercise.J Appl Physiol1996;80:363–6.
37.Ali A,Gardiner R,Foskett A,Gant N.Fluid balance,thermoregulation and sprint and passing skill performance in female soccer players.Scand J Med Sci Sports2011;21:437–45.
38.Cooper KE.Some responses of the cardiovascular system to heat and fever.Can J Cardiol1994;10:444–8.
39.Bell AW,Hales JR,King RB,Fawcett AA.Influenc of heat stress on exercise-induced changes in regional blood fl w in sheep.J Appl Physiol1983;55:1916–23.
40.Hultman E,Sahlin K.Acid–base balance during exercise.Exerc Sport Sci Rev1980;8:41–128.
41.Suzuki K,Peake J,Nosaka K,Okutsu M,Abbiss CR,Surriano R,et al. Changes in markers of muscle damage,inflammatio and HSP70 after an Ironman Triathlon race.Eur J Appl Physiol2006;98:525–34.
42.Puntschart A,Vogt M,Widmer HR,Hoppeler H,Billeter R.Hsp70 expression in human skeletal muscle after exercise.Acta Physiol Scand1996;157:411–7.
43.Sandström ME,Madden LA,Taylor L,Siegler JC,Lovell RJ,Midgley A, et al.Variation in basal heat shock protein 70 is correlated to core temperature in human subjects.Amino Acids2009;37:279–84.
44.Lovell R,Madden L,Carroll S,McNaughton L.The time-profil of the PBMC HSP70 response toinvitroheat shock appears temperature-dependent.Amino Acids2007;33:137–44.
45.Milne KJ,Noble EG.Exercise-induced elevation of HSP70 is intensity dependent.J Appl Physiol2002;93:561–8.
46.Yamada P,Amorim F,Moseley P,Schneider S.Heat shock protein 72 response to exercise in humans.Sports Med2008;38:715–33.
47.Niess AM,Simon P.Response and adaptation of skeletal muscle to exercise-the role of reactive oxygen species.Front Biosci2007;12:4826–38.
48.Radak Z.Free radicals in exercise and aging.Champaign,IL:Human Kinetics;2000.
49.Bloomer RJ,Davis PG,Consitt LA,Wideman L.Plasma protein carbonyl response to increasing exercise duration in aerobically trained men and women.Int J Sports Med2007;28:21–5.
50.Miyata M,Kasai H,Kawai K,Yamada N,Tokudome M,Ichikawa H,et al. Changes of urinary 8-hydroxydeoxyguanosine levels during a two-day ultramarathon race period in Japanese non-professional runners.Int J Sports Med2008;29:27–33.
51.Sureda A,Ferrer MD,Tauler P,Romaguera D,Drobnic F,Pujol P,et al. Effects of exercise intensity on lymphocyte H2O2production and antioxidant defences in soccer players.Br J Sports Med2009;43:186–90.
52.Hartmann A,Pfuhler S,Dennog C,Germadnik D,Pilger A,Speit G. Exercise-induced DNA effects in human leukocytes are not accompanied by increased formation of 8-hydroxy-2′-deoxyguanosine or induction of micronuclei.Free Radic Biol Med1998;24:245–51.
53.Mastaloudis A,Tian WY,Obert P,O’donnell FB,Roderick H,Dashwood RH,et al.Endurance exercise results in DNA damage as detected by the COMET ASSAY.Free Radic Biol Med2004;36:966–75.
54.Pereira Ede J,Panek AD,Eleutherio EC.Protection against oxidation during dehydration of yeast.Cell Stress Chaperones2003;8:120–4.
55.Zuo L,Christof FL,Wright VP,Liu CY,Merola AJ,Berliner LJ,et al. Intra-and extracellular measurement of reactive oxygen species produced during heat stress in diaphragm muscle.Am J Physiol Cell Physiol2000;279:1058–66.
56.Madara JL,Pappenheimer JR.Structural basis for physiological regulation of paracellular pathways in intestinal epithelia.J Membr Biol1987;100:149–64.
57.Nose H,Mack GW,Shi XR,Nadel ER.Involvement of sodium retention hormones during rehydration in humans.J Appl Physiol1988;65:332–6.
Received 30 October 2014;revised 3 January 2015;accepted 13 May 2015
Available online 25 September 2015
Peer review under responsibility of Shanghai University of Sport.
*Corresponding authors.
E-mail addresses:ppaik@yonsei.ac.kr(I.-Y.Paik),ssh@yonsei.ac.kr (S.-H.Suh).
†The firs two authors(Hee-Tae Roh and Su-Youn Cho)contributed equally to this work.
http://dx.doi.org/10.1016/j.jshs.2015.09.007
2095-2546/©2016 Production and hosting by Elsevier B.V.on behalf of Shanghai University of Sport.This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
杂志排行

Journal of Sport and Health Science的其它文章
- Biomarker-guided classificatio scheme of neurodegenerative diseases
- Mechanism of neurodegeneration through tau and therapy for Alzheimer’s disease
- Examining the relationship between sport and health among USA women: An analysis of the Behavioral Risk Factor Surveillance System
- The microbiome,microbial-generated proinflammato y neurotoxins, and Alzheimer’s disease
- Longitudinal trajectories of physical activity in women using latent class growth analysis:The WIN Study
- Evidence of a conservative gait strategy in athletes with a history of concussions