高寒草甸草地微斑块植物特征及其土壤性质的研究
2016-01-28石明明张永超张典业任运涛宗文杰傅华牛得草
石明明,张永超,张典业,任运涛,宗文杰,傅华,牛得草*
(1.草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2.甘肃省玛曲县畜牧兽医局,甘肃 甘南 747300)
高寒草甸草地微斑块植物特征及其土壤性质的研究
石明明1,张永超1,张典业1,任运涛1,宗文杰2,傅华1,牛得草1*
(1.草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2.甘肃省玛曲县畜牧兽医局,甘肃 甘南 747300)
摘要:植物群落的斑块化与维持是草地对外界干扰的响应,也是植物群落及其生物多样性持续发展的基础。分析了3个不同鼠洞数量梯度下斑块种类、数量、面积的变化和中度鼠洞数下5种主要斑块植物群落的结构及其土壤性质。结果表明,在中度鼠洞数下,斑块的种类和总数量最高;斑块的总面积随鼠害的加重呈增加趋势;从中度到重度梯度上,鹅绒委陵菜、黄帚橐吾和草玉梅斑块的数量和面积都增加,且面积增大剧烈,乳白香青和火绒草等斑块的数量和面积都减小,具有消失的趋势;在重度鼠洞数量下草地微斑块表现为由少数起主导作用的斑块组成,斑块格局变的较为简单。斑块中单一物种生物量的变异性增加是斑块最主要的特征,这种单一物种的大量繁殖影响着斑块中植物的功能群结构和物种的多度,从而通过不同形式对干扰产生缓冲作用。植物群落的斑块化影响着土壤的异质性,对不同斑块土壤性质进行主成分分析,除乳白香青和基质斑块,其余4种斑块土壤都具有较低的全氮、碱解氮、速效磷,且不同斑块间土壤全氮、碱解氮和速效磷的变异系数都较高,可见,土壤全氮、碱解氮和速效磷的含量以及空间异质性在响应植被的演替上较敏感,因此,高寒草甸生态系统中维持全氮和速效养分资源的供应对维持生态系统的稳定具有重要的意义。
关键词:微斑块;功能群;主成分分析;土壤异质性;鼠洞数量
Plant traits and soil properties in pasture mini-patches in an alpine meadow
SHI Ming-Ming1, ZHANG Yong-Chao1, ZHANG Dian-Ye1, REN Yun-Tao1, ZONG Wen-Jie2, FU Hua1, NIU De-Cao1*
StateKeyLaboratoryofGrasslandAgro-ecosystem,CollegeofPastoralAgricultureScienceandTechnology,LanzhouUniversity,Lanzhou730020,China
Abstract:The patchiness and persistence of plant communities represent grassland responses to outside disturbance, and are the basis of sustainable development of plant communities and biodiversity. The aim of this study was to understand the factors contributing to the maintenance and development of plant communities and the effects of patchiness on soil properties after a disturbance. The changes in the types, number, and areas of mini-patches in meadows were evaluated in meadows with a light, moderate, and heavy density of plateau pika (Ochotona curzoniae) burrows. The plant community structure and soil properties were evaluated for five types of patches in meadows with a moderate number of burrows. Some mini-patches were characterized by a high abundance of a single species, with mini-patch areas ranging from 0.5 m2to 100 m2in an alpine meadow (not including background patches). These types of patches were named according to the main plant species. We chose three approximately 1-hectare sample sites with different burrow densities; light, moderate, and heavy. Three quadrats (20 m×20 m) were placed randomly to determine the types, number, and area of patches within each site. We chose five types of patches from the three sample sites, and selected three typical patches for each type to determine the plant community structure and soil properties. Quadrat sampling (1 m×1 m) was used to determine basic plant traits (composition, height, cover degree, above-ground biomass of vegetation) and soil properties (organic carbon, pH, total nitrogen, available nitrogen, total phosphorus, available phosphorus) in the patches. The meadows with moderate burrow density had the most types of patches and the greatest abundance of patches. The total area of patches tended to increase with greater deterioration caused by plateau pika. The number and areas of Potentilla anserine, Ligularia virgaurea, and Anemone rivularis patches tended to increase, and their areas increased dramatically from the moderate to heavy burrow density. The number and areas of Anaphalis lacteal and Leontopodium japonicum patches tended to decrease, and these patches tend to disappear from as the burrow density increased from moderate to heavy. There were few main types of pasture mini-patches, and the pattern of patches was relatively simple under heavy burrow density. An abnormal increase in the biomass of a single species was the main characteristic of the patches. The expanding propagation of single species affected the structures of plant functional groups and species abundance, and thus, cushioned the effects of other types of disturbance. The results of the principal component analysis showed that the amounts of total nitrogen, available nitrogen, and available phosphorus in soil were lower in four types of patches than in A. lacteal patches and background patches. The coefficients of variation were higher for soil total nitrogen, available nitrogen, and available phosphorus than for other soil properties in patches in the meadows with moderate burrow density. The contents and spatial heterogeneity of soil total nitrogen, available nitrogen, and available phosphorus were more sensitive than the responses of vegetation succession to plateau pika activity. Therefore, it is very important that the supply of total nitrogen and available nutrients are maintained to retain the stability of the alpine meadow ecosystem.
Key words:mini-patch; functional group; principal component analysis; soil heterogeneity; number of burrowing plateau pika
随着生境丧失和破碎程度的日益加重,草地空间异质性增强,在小尺度上由于不同路径或起始点演替的发生,连续的植物群落演变为不同特征斑块组成的镶嵌体[1]。植物群落在时间尺度上通常表现总体稳定而局部变化的特点,因此,研究斑块的群落特征以及生态作用过程,是了解植物群落空间组织过程和格局形成机制的切入点。植物群落斑块的生态过程和特点往往反映了生态系统的总体结构与功能的动态变化特征,从小尺度斑块入手对草地的总体特征进行整合研究显得非常重要[2]。近年来,小尺度上空间异质性的研究受到广泛关注,张卫国等[3]研究了不同放牧强度下斑块植物群落性状和格局的变化,指出草地微斑块的形成和草地退化有着相同的推动力,这种作用力主要来自于放牧压力和鼠类活动等外界干扰。受环境压力的不均匀性影响,草地空间异质性明显,景观水平上表现为不同的斑块类型、数量及大小的组合镶嵌,而植物群落的斑块化与维持也构成植物群落及其生物多样性持续发展的基础,推动着草地向着不同的方向演替[4]。鼠害是影响植物群落空间结构的环境因子之一,鼠类的扩张将推动草地植被的逆向演替,鼠害发生程度反映了草地的退化程度[5]。空间异质性通常表现为植被的空间格局以及土壤资源的异质性,两者相互影响和相互制约[6-7]。目前,关于草地微斑块的报导大多集中于斑块植物格局变化的研究,对微斑块植物群落及其土壤养分特征的研究较少[3,8]。为此,本文分析了鼠害发生程度与草地群落斑块空间格局变化的关系以及中度鼠害下草地几种微斑块的植物群落和土壤异质性特征,以了解存在干扰时植物群落维持与发展的特征以及斑块化对土壤性质的影响,为阐述在受到干扰时植物群落维持和演替机制提供理论基础。
1材料与方法
1.1 研究区自然概况
研究区位于玛曲县县城以西10 km处的亚高山草甸类草地,地理坐标为33°50′ N,102°05′ E,平均海拔3500 m左右,属于典型的寒温湿润气候,具有明显的高原气候特征;年均温度1.5℃,日平均通过0℃的年活动积温为1520℃;年降水量600~800 mm,主要集中在6-9月份;全年日照时数为2290 h左右。土壤为亚高山草甸土。群落中植物以莎草科和禾本科为主,混有其他杂草类,主要植物有禾叶嵩草(Kobresiagraminifolia)、垂穗披碱草(Elymusnutans)、长毛风毛菊(Saussureahieracioides)、丝叶毛茛(Ranunculustanguticus)、鹅绒委陵菜(Potentillaanserina)、莓叶委陵菜(Potentillafragarioides)、兰石草(Lanceatibetica)等。
1.2 研究方法
1.2.1样地设置亚高山高寒草甸草原中鼠害发生程度是草地退化的主要判断指标。于2012年5月初,在研究区以有效鼠洞[鼠种为高原鼠兔(Ochotonacurzoniae)]数量差异为梯度选择了3块样地,分别为轻、中、重度鼠害发生样地,每个样地面积大约1 hm2。样地间距最远为2 km,地势平坦。样地于2012年5月上旬围封。草地围封前为自由放牧地,放牧家畜为牦牛,以上3块样地由于离家畜宿营地距离不一致,家畜采食程度不一致,导致鼠害密度存在差别。
1.2.2微斑块的界定及种类划分根据亚高山高寒草甸植物群落的特点及研究的目的,参照张卫国等[3]的方法,将草地群落斑块界定为面积在0.5 m2到100 m2范围内的微斑块(不包括基质斑块),斑块与周围植物群落物种组成不同,表现为某一物种高度聚集,从而与周边植被具有明显的轮廓线,并以斑块中这种高度聚集的植物种对斑块进行命名,这些斑块周围的背景植被即为基质斑块。
1.2.3微斑块基本性状的测定2012年7月下旬,对3块样地中斑块的种类、数量和面积进行统计;再在中度鼠害样地中选择具有代表性的斑块进行斑块植物性状测定及其土壤取样,每种斑块3个重复。
斑块的种类、数量、面积的统计:各样地随机设置3个20 m×20 m的样方,将每个样方均分为面积为2 m×2 m 的100 个网格,并用线绳标记,逐一识别并命名斑块; 统计斑块的种类、数量、面积。
斑块植物性状测定:在中度鼠害发生地,选择共有的斑块类型,每种类型选3个典型斑块,在其内设置2个0.5 m×0.5 m 的小样方,1个为固定样方,调查植物的基本性状包括植被组成、高度、盖度,另1个为测产样方,分种调查地上生物量,各项指标的具体测定用常规方法进行[9]。
土壤取样及其各指标的测定[10]:在测产样方内,用土钻采集斑块内0~10 cm、10~20 cm土层的土壤样品,并使用环刀测定土壤容重。土壤样品带回实验室,测定土壤有机碳、pH、全氮、碱解氮、全磷、速效磷。
1.3 数据处理
用Origin 9.0作图,Excel 2007、SPSS 16.0软件对试验数据进行统计分析。
2结果与分析
2.1 有效鼠洞数量与斑块种数、数量、面积之间的关系
鼠害发生强度的增加提高了草地的空间异质性,在轻、中、重度3种鼠害发生强度下,微斑块的种类、数量、面积明显不同(表1),在一定程度上反映了草地空间异质性的程度。斑块的种类表现为中度鼠害处理下最为丰富,为8种,轻度和重度鼠害处理下分别为5和7种。在3种梯度下,斑块的总数量和斑块的种类表现出相同的变化趋势,即先增加后降低,中度和重度都显著高于轻度(P<0.05),但是鹅绒委陵菜、黄帚橐吾和草玉梅斑块的数量呈增加趋势,其中黄帚橐吾斑块在3个处理间都差异显著(P<0.05)。斑块的总面积随有效鼠洞数的增加而显著增加(P<0.05),鹅绒委陵菜、黄帚橐吾、草玉梅和裸地斑块的面积与总面积变化趋势相同,且从中度到重度梯度上差异显著(P<0.05),而火绒草和乳白香青等斑块表现出先增加后降低,且中度到重度梯度上差异显著(P<0.05)。其中,在重度鼠害发生地,鹅绒委陵菜、黄帚橐吾和草玉梅斑块的面积分别为17.3,30.2和19.5 m2,从中度鼠害到重度鼠害梯度上变化剧烈。3种鼠害梯度下,斑块总面积占样地的面积分别为4.3%,12.4%和20.4%。除鹅绒委陵菜、黄帚橐吾、草玉梅和裸地斑块,其余斑块的面积和与斑块总面积的比例从中度到重度梯度上由60.9%猛减为10.9%,可见,这些斑块具有消失的趋势,重度处理下斑块格局变的较为简单。这些结果说明,从中度干扰到重度干扰草地已经到了由量变(斑块种类和总数量多,各斑块面积和总面积都中等)到质变(斑块总面积大,一些斑块扩张剧烈)。
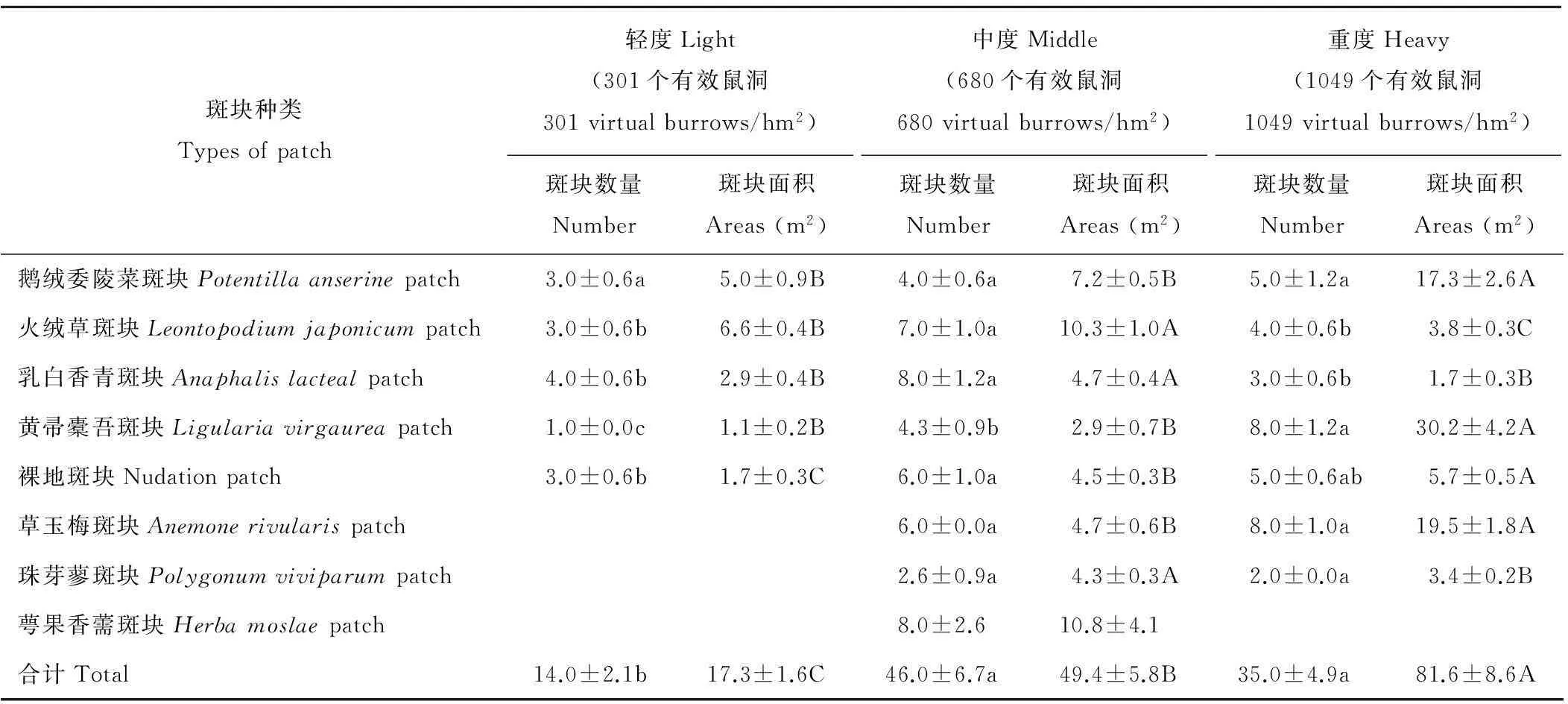
表1 不同鼠洞数量梯度下斑块的种类、数量、面积
注:表中数据为平均值±标准误。同行中不同小写字母表示斑块数量差异显著(P<0.05),不同大写字母表示斑块面积差异显著(P<0.05)。
Note: Data in the Table are means±standard error. Different lowercase letters and capital letters represent significant differences of number and areas in the same row (P<0.05).
2.2 不同微斑块群落结构的特征
中度鼠害发生地不同微斑块,在种群水平上(表 2),鹅绒委陵菜、火绒草、乳白香青和黄帚橐吾斑块中建群种生物量明显增加,其他种的生物量显著减少;乳白香青和黄帚橐吾斑块中,尽管表征性植物生物量很大,但物种数和基质斑块相差很小。在功能群水平上(图1),各斑块中生活型功能群组成基本一致,均由莎草类、禾草类、杂草类组成。植物生活型功能群生物量比例在不同斑块间差异显著(P<0.05),在基质斑块中3类生活型所占的比例相差不大,在其他4种斑块中杂草类的比例显著高于莎草类和禾草类,其中鹅绒委陵菜和黄帚橐吾斑块中杂草类的比例分别达到86%和83%,而禾草类相对于基质斑块显著降低(P<0.05)。在群落水平上,各斑块在植物种数、物种组成、地上生物量、地下生物量等性状指标上表现出明显的差异(表 2),鹅绒委陵菜斑块的物种数显著低于基质斑块,由于斑块中建群种本身的特征,乳白香青和黄帚橐吾斑块的地上生物量显著高于基质斑块(P<0.05),地下生物量乳白香青斑块和基质相差不大,其他各类斑块显著低于基质(P<0.05)。除基质斑块外,其他4种斑块在种群、功能群和群落水平上都表现出不均衡性。

图1 不同斑块上植物生活型功能群组成及生物量比例Fig.1 Structure and biomass proportion of plant functional group on different mini-patch
2.3 不同斑块土壤性质的主成分分析
对不同斑块0~10 cm土层土壤性质进行主成分分析(图2,表3),提取出3个特征值大于1的公共因子。第一主成分的方差贡献率为49.86%,在这一主成分中全N、碱解N、速效P和pH具有较高的载荷,第二主成分的方差贡献率为26.70%,这一主成分中有机碳和容重具有较高的载荷,第三主成分的方差贡献率为17.08%,这一主成分中全P具有较高的载荷。不同斑块的主成分得分反映出不同斑块土壤的养分和物理特性存在明显差异。在第一主成分轴上,乳白香青斑块有较高得分,基质次之,其他斑块得分都较低,表明乳白香青斑块的形成与土壤全N、碱解N、速效P含量的关密切系;在第二主成分轴上,黄帚橐吾斑块和基质得分较高, 各点在该轴上分布较零散;在第三主成分轴上, 火绒草有较高得分, 可能其斑块发育趋向于土壤具有较高的全磷特征。对10~20 cm土层(图3,表4),提取出2个特征值大于1的主成分,其中,第一主成分的方差贡献率为50.50%,这一主成分中全N、碱解N、速效P、pH和容重具有较高的载荷,第二主成分的方差贡献率为33.42%,这一主成分中有机碳、全P具有较高的载荷;不同斑块的主成分得分明显不同,在第一主成分轴上,乳白香青斑块得分较高,其他斑块得分都较低,在第二主成分轴上,裸地、鹅绒委陵菜、基质斑块得分较高。变异系数可以反应总体的变异程度,0~10 cm土层,不同斑块间土壤有机碳、全氮、碱解氮、全磷和速效磷的变异系数分别为0.14,0.45,0.42,0.22和0.69。
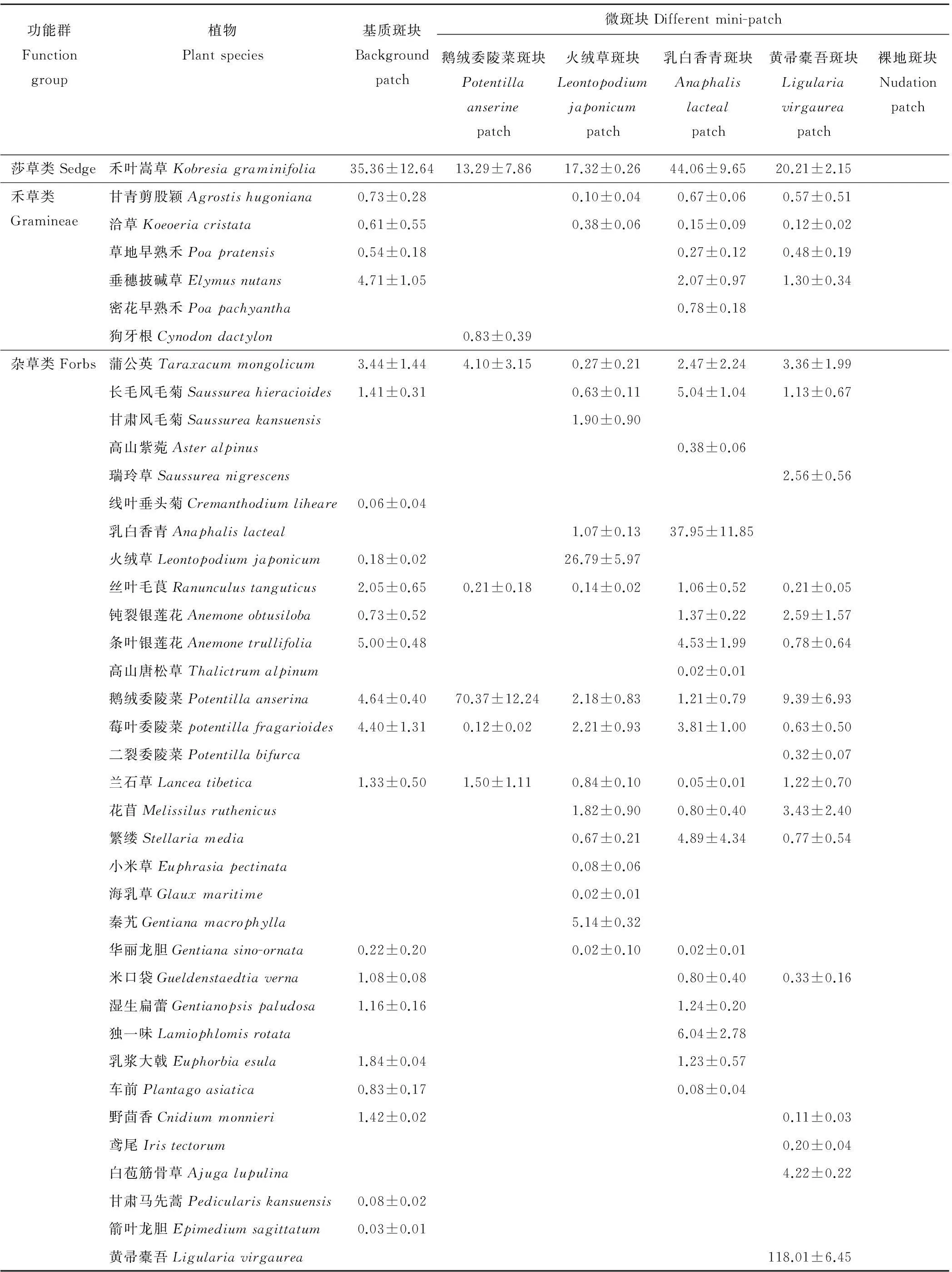
表2 不同微斑块的植物种类组成及生物量
续表2Continued
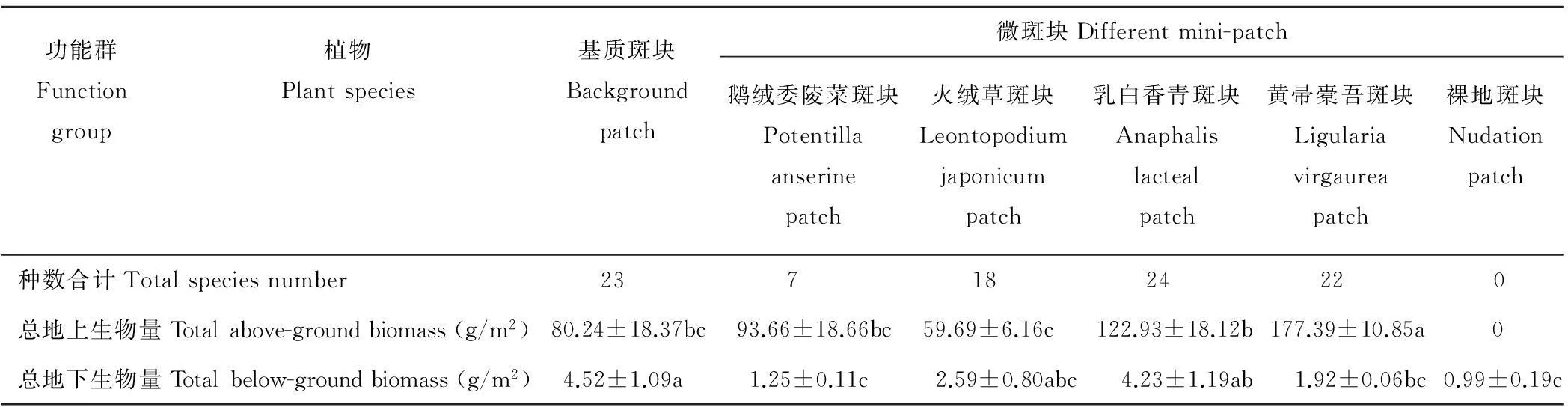
功能群Functiongroup植物Plantspecies基质斑块Backgroundpatch微斑块Differentmini-patch鹅绒委陵菜斑块Potentillaanserinepatch火绒草斑块Leontopodiumjaponicumpatch乳白香青斑块Anaphalislactealpatch黄帚橐吾斑块Ligulariavirgaureapatch裸地斑块Nudationpatch种数合计Totalspeciesnumber 23 7 18 24 22 0总地上生物量Totalabove-groundbiomass(g/m2)80.24±18.37bc93.66±18.66bc59.69±6.16c122.93±18.12b177.39±10.85a 0总地下生物量Totalbelow-groundbiomass(g/m2)4.52±1.09a1.25±0.11c2.59±0.80abc4.23±1.19ab1.92±0.06bc0.99±0.19c
注:表中数据(除后3行)为各植物种在斑块中的地上生物量±标准误(g/m2),同行中不同小写字母表示差异显著(P<0.05)。空白部分表示该斑块没有此物种,裸地斑块中的地下生物量为死根。
Note: Data in the Table are means±standard error(g/m2),the means is above-ground biomass of each plant species in the patch (not including after the three lines). Different lowercase letters represent significant differences in the same row (P<0.05). The blank section shows without this species on the patches. Below-ground biomass of nudation patch is dead root.
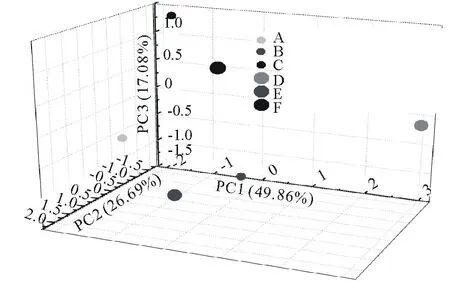
图2 不同斑块在0~10 cm土层土壤性质主成分分析的第一、二、三主成分上的分布Fig.2 The plot of the first to three component of principal component analysis results for soil properties at 0-10 cm soil horizon
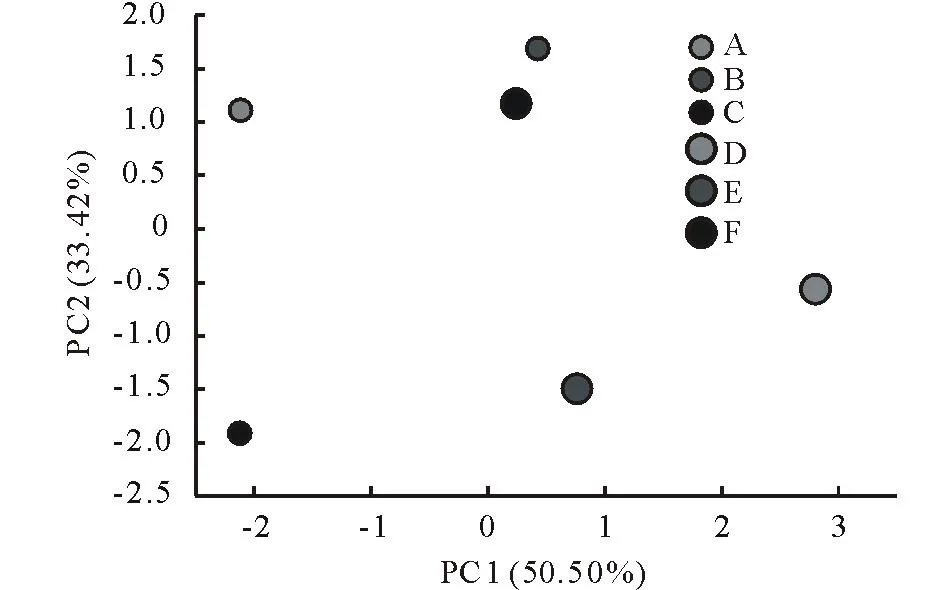
图3 不同斑块在10~20 cm土层土壤性质主成分分析的第一、二主成分上的分布Fig.3 The plot of the first and second component of principal component analysis results for soil properties at 10-20 cm soil horizon
A:裸地斑块Nudation patch;B:鹅绒委陵菜斑块Potentillaanserinapatch;C:火绒草斑块Leontopodiumjaponicumpatch;D:乳白香青斑块Anaphalislactealpatch;E:黄帚橐吾斑块Ligulariavirgaureapatch;F:基质斑块Background patch.
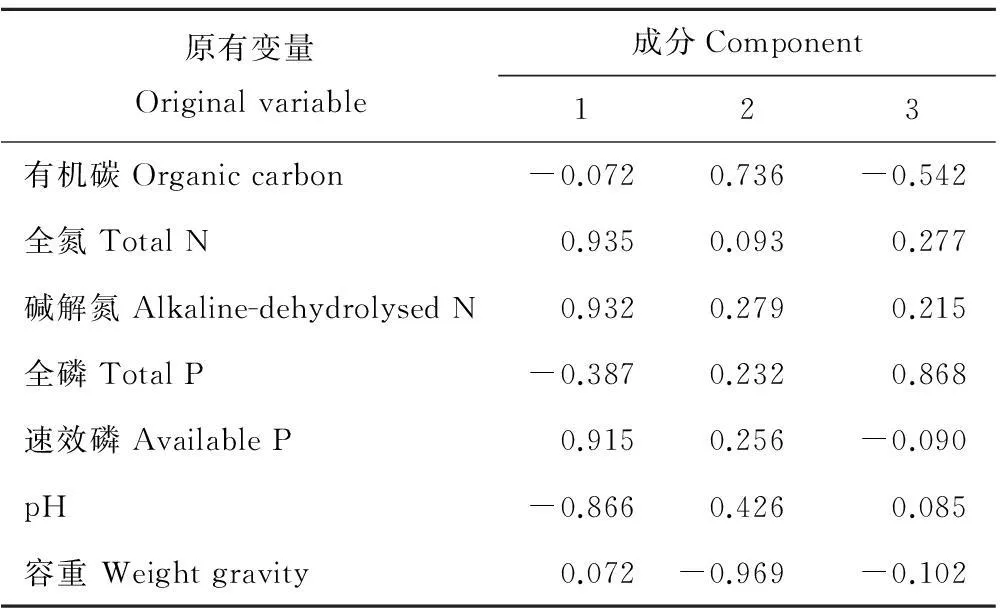
表3 0~10 cm土层因子载荷矩阵
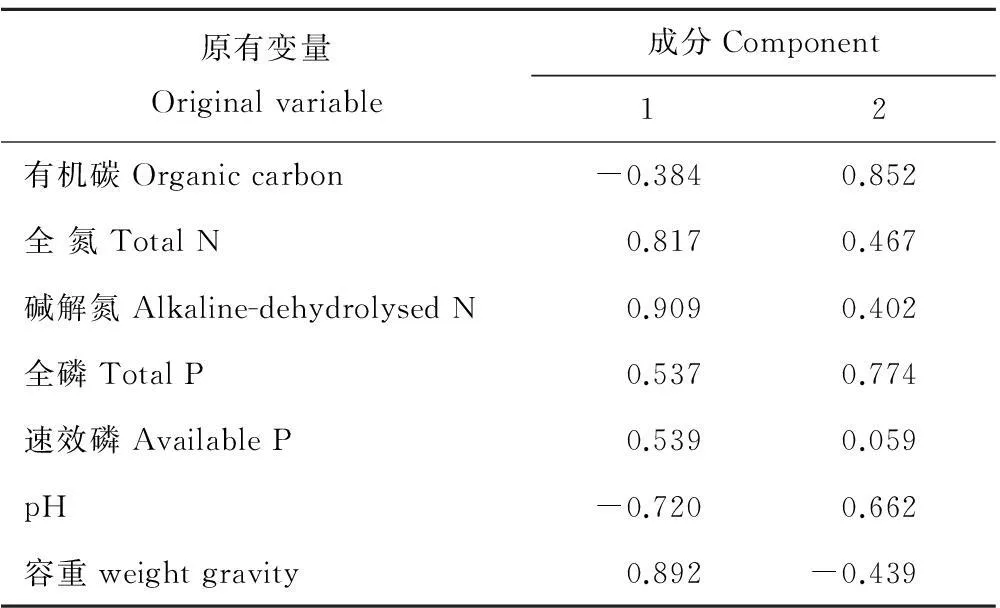
表4 10~20 cm土层因子载荷矩阵
3讨论
有效鼠洞密度的差异是反映草地退化程度具有表征意义的重要指标[5]。在轻、中、重3个鼠害发生梯度下,草地微斑块的种类和总数量表现为先随鼠害强度增加而上升,中度梯度下达最高值后转为随鼠害强度增加而下降,而鹅绒委陵菜、黄帚橐吾和草玉梅斑块的数量呈增加趋势,可见干扰活动(放牧,鼠害等)影响了植被的空间格局,促使了背景植被(基质)的分割,草地表现为斑块化的镶嵌结构。斑块化-持久性假说(patchiness persistence hypothesis)[11]认为,群落处于逆行演替的中期阶段时,斑块化的程度最大,我们研究的结果也支持这一假说:在中度鼠害发生强度下,斑块的种类和数量最高。鼠害强度的增加为某些植物的定植创造了条件,表现为非均匀性同种个体的聚集,斑块化程度增强,随着鼠类活动的进一步增强,植物种数目减少,一些物种的竞争及扩散被限制,斑块格局变的较为简单[12]。与此同时,斑块的总面积表现为随鼠害强度增加而增加,从中度鼠害到重度鼠害梯度上,鹅绒委陵菜、黄帚橐吾和草玉梅斑块的面积剧烈增大,而其余斑块的面积减小,具有消失的趋势。干扰活动改变了草地的生境条件,干扰敏感种减少甚至消失,而适性植物大量繁殖,斑块总面积呈增加趋势。在重度干扰下,草地微斑块表现为由少数起主导作用的斑块组成,草地群落具有均质化的趋势,而草地的生态作用和利用价值发生了根本性的转变(斑块总面积占样地的面积为20.4%)。
群落中的少数关键种决定着该群落的基本特征,斑块中表征性植物种大量繁殖对干扰具有一定的缓冲作用[7]。均衡效应(portfolio effect)可作为群落具有高稳定性的原因之一[13],受干扰的草地群落结构发生变化,群落通过不同方式(物种多样性增加,单一物种生物量异常增加)增加均衡效应使群落趋于稳定。在种群水平上,斑块(除基质外)中建群种多度显著高于其他种,可见不同植物对干扰的敏感性不同,在竞争过程中,干扰敏感种减少了其生物量,通过竞争释放竞争空间使其他物种的多度增加,从而对系统的功能起到补偿作用[12-15]。在功能群水平上,不仅群落中功能群植物的比例发生了变化,而且功能群内物种组成也发生了明显的变化,如在火绒草和乳白香青斑块中,它们的多年生杂类草功能群在群落中分别占65%和50%,而该功能群中的物种数分别为15和18种,可见在生态适应上功能群的生态补偿作用表现出多样化[15]。在群落水平上,乳白香青和黄帚橐吾斑块中,虽然表征性植物生物量很大,但物种数和基质斑块差异不大,这种多物种的共存可能是由于不同物种对生境资源利用的分化与互补[2],说明这些斑块通过高的物种丰富度和单一物种生物量变异性增加两种方式使群落趋于稳定。
草地微斑块的存在是退化草地最基本的特征之一,其形成受环境因子以及植物对环境的生物和生态适应性差异的影响,而这些植物斑块的维持与发展将影响土壤养分和结构的异质化过程[16-18]。对不同斑块土壤特性进行主成分分析发现,在微斑块内0~10 cm土层土壤特性主要受控于全N、碱解N、速效P、pH和有机碳以及全P三大因素影响,各因素的方差贡献率分别为49.86%,26.70%和17.08%;在10~20 cm土层,主要受控于全N、碱解N、速效P、pH、容重和有机碳、全P两大因素影响,各因素的方差贡献率分别为50.50%和33.42%,说明土壤的异质性随深度增加而减小。植物通过改变凋落物的质与量或改善小气候条件影响土壤养分资源的循环,进而引起土壤的空间异质性[19]。表层土壤性质的主成分分析显示,乳白香青斑块具有较高的土壤全N、碱解N、速效P,可能是乳白香青斑块中发达的植物根系影响着土壤-植物系统的养分循环,高的地下生物量又增加了土壤有效养分的输入[20]。黄帚橐吾斑块在第二主成分上具有较高得分,表明有机碳含量较高,可能是受凋落物的显著影响[19]。0~10 cm土层,在第一主成分轴上,除乳白香青和基质斑块,其余斑块得分都较低,表明这四种斑块土壤都具有较低全氮、碱解氮、速效磷,且不同斑块间土壤全氮、碱解氮和速效磷的变异系数(分别为0.45,0.42,0.69)都较高,可见,土壤全氮、碱解氮和速效磷的含量以及空间异质性在响应植被的演替上较敏感,因此,高寒草甸生态系统中维持全氮和速效养分资源的供应对维持生态系统的稳定具有重要的意义。
References:
[1]Liu Z G, Li Z Q. Perspectives on small-scale spatial structure of plant species in plant communities. Acta Phytoecologica Sinica, 2006, 29(6): 1020-1028.
[2]Ye W H. The maintenance mechanism of plant community and its species diversity. Chinese Biodiversity, 2000, 8(1): 17-24.
[3]Zhang W G, Huang W B, Yang Z Y. The study on the relationship between mini-patch and degradation of pasture. Acta Prataculturae Sinica, 2003, 12(3): 44-50.
[4]Bai Y F, Xu Z X, Li D X. On the small scale spatial heterogeneity of soil moisture, carbon and nitrogen inStipacommunities of the Inner Mongolia Plateau. Acta Ecologica Sinica, 2002, 22(8): 1215-1223.
[5]Zhang W G, Jiang X L, Wang S M,etal. Effects of mound-building activity of zokor (Myospalaxbaileyi) and different regimes of grazing-prohibited on vegetation productivity. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(10): 1882-1887.
[6]Begona Peco, Sanchez A M, Azcarate F M. Abandonment in grazing systems: Consequences for vegetation and soil. Agriculture, Ecosystems and Environment, 2006, 113(1): 284-294.
[7] Alados C L, Navarro T, Komac B,etal. Do vegetation patch spatial patterns disrupt the spatial organization of plant species. Ecological Complexity, 2008, 6(2): 197-207.
[8]Wu Y N, Luo W T, Huo G W,etal. Spatial heterogeneity dynamics of soil carbon and nitrogen in the mini-patches during degeneration succession of grassland communities. Acta Ecologica Sinica, 2014, 19: 5549-5557.
[9]Ren J Z. Research on Pratacultural Science Methods[M]. Beijing: Chinese Agricultural Press, 1998: 11-15.
[10]Bao S D. Soil Agro-chemistrical Analysis[M]. Beijing: Chinese Agricultural Press, 2000.
[11]Hastings H M, Pekelney R, Monticciolo R,etal. Time scales, persistence and patchiness. BioSystems, 1982, 15(4): 281-289.
[12]Xin X P, Gao Q, Li Y Y,etal. Fractal analysis of grass patches under grazing and flood disturbance in an alkaline grassland. Acta Botanica Sinica, 1999, 41(3): 307-313.
[13]Doak D F, Bigger D, Harding E K,etal. The statistical inevitability of stability-diversity relationships in community ecology. The American naturalist, 1998, 151(3): 264-276.
[14]Bai Y F, Chen Z Z. Effects of long-term variability of plant species and functional groups on stability of aLeymuschinensiscommunity in the Xilin river basin, Inner Mongolia. Acta Phytoecologica Sinica, 2000, 24(6): 641-647.
[15]Jiang X L, Zhang W G, Yang Z Y,etal. The inf luence of disturbance on community structure and plant diversity of alpine meadow. Acta Botanica Boreali-Occidentalia Sinica, 2004, 23(9): 1479-1485.
[16]Liang S C, Zhang S M, Yu F H,etal. Small-scale spatial cross-correlation between ramet population variables ofPotentillareptansvar. sericophylla and soil available phosphorus. Journal of Plant Ecology, 2007, 31(4): 613-618.
[17]Cheng X L, An S Q, Li Y,etal. The individual distribution patterns and soil elements heterogeneity during the degradation of grassland in Ordos. Acta Phytoecologica Sinica, 2003, 27(4): 503-509.
[18]Li Q, Zhou D W, Song Y T. The distribution features of two widespread legumes and their relationships with soil factors in Songnen grassland. Acta Prataculturae Sinica, 2014, 23(1): 31-40.
[19]Hu Z L, Pan G X, Li L Q,etal. Changes inpools and heterogeneity of soil organic carbon, nitrogen and phosphorus under different vegetation types in Karst mountainous area of central Guizhou Province, China. Acta Ecologica Sinica, 2009, 29(8): 4187-4195.
[20] Fornara D A, Tilman D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. Journal of Ecology, 2008, 96(2): 314-322.
参考文献:
[1]刘振国, 李镇清. 植物群落中物种小尺度空间结构研究. 植物生态学报, 2006, 29(6): 1020-1028.
[2]叶万辉. 物种多样性与植物群落的维持机制. 生物多样性, 2000, 8(1): 17-24.
[3]张卫国, 黄文冰, 杨振宇. 草地微斑块与草地退化关系的研究. 草业学报, 2003, 12(3): 44-50.
[4]白永飞, 许志信, 李德新. 内蒙古高原针茅草原群落土壤水分和碳、氮分布的小尺度空间异质性. 生态学报, 2002, 22(8): 1215-1223.
[5]张卫国, 江小蕾, 王树茂, 等. 鼢鼠的造丘活动及不同休牧方式对草地植被生产力的影响. 西北植物学报, 2004, 24(10): 1882-1887.
[8]乌云娜, 雒文涛, 霍光伟, 等. 草原群落退化演替过程中微斑块土壤碳氮的空间异质动态. 生态学报, 2014, 19: 5549-5557.
[9]任继周. 草业科学研究方法[M]. 北京: 中国农业出版社, 1998: 11-15.
[10]鲍士旦.土壤农化分析[M]. 北京: 中国农业出版社, 2000.
[12]辛晓平, 高琼, 李宜垠, 等. 放牧和水淹干扰对松嫩平原碱化草地空间格局影响的分形分析. 植物学报, 1999, 41(3): 307-313.
[14]白永飞, 陈佐忠. 锡林河流域羊草草原植物种群和功能群的长期变异性及其对群落稳定性的影响. 植物生态学报, 2000, 24(6): 641-647.
[15]江小蕾, 张卫国, 杨振宇, 等. 不同干扰类型对高寒草甸群落结构和植物多样性的影响. 西北植物学报, 2004, 23(9): 1479-1485.
[16]梁士楚, 张淑敏, 于飞海, 等. 绢毛匍匐委陵菜与土壤有效磷的小尺度空间相关分析. 植物生态学报, 2007, 31(4): 613-618.
[17]程晓莉, 安树青, 李远, 等. 鄂尔多斯草地退化过程中个体分布格局与土壤元素异质性. 植物生态学报, 2003, 27(4): 503-509.
[18]李强, 周道玮, 宋彦涛. 松嫩草地两种广布豆科植物分布特征及其与土壤因子关系. 草业学报, 2014, 23(1): 31-40.
[19]胡忠良, 潘根兴, 李恋卿, 等. 贵州喀斯特山区不同植被下土壤C、N、P含量和空间异质性. 生态学报, 2009, 29(8): 4187-4195.
石明明,张永超,张典业,任运涛,宗文杰,傅华,牛得草. 高寒草甸草地微斑块植物特征及其土壤性质的研究.草业学报, 2015, 24(9): 197-205.
SHI Ming-Ming, ZHANG Yong-Chao, ZHANG Dian-Ye, REN Yun-Tao, ZONG Wen-Jie, FU Hua, NIU De-Cao. Plant traits and soil properties in pasture mini-patches in an alpine meadow. Acta Prataculturae Sinica, 2015, 24(9): 197-205.
通讯作者*Corresponding author. E-mail:xiaocao0373@163.com
作者简介:石明明(1990-),男,甘肃天水人,在读硕士。E-mail:734341754@qq.com
基金项目:公益性行业(农业)科研专项经费(201203041),“长江学者和创新团队发展计划(IRT13019)”,国家科技支撑项目(2012BAD13B05)和国家自然科学基金(31201837,31172258)资助。
收稿日期:2015-02-20;改回日期:2015-05-06
DOI:10.11686/cyxb2015083http://cyxb.lzu.edu.cn
