慢性铝暴露对大鼠海马钙离子稳态及CaMKⅡ和CREB活性的影响
2016-01-28张海燕孙雅萍郭庆国
宋 杨 张海燕 孙雅萍 郭庆国 孟 欣 王 彪
(沈阳市第四人民医院,辽宁 沈阳 110120)
慢性铝暴露对大鼠海马钙离子稳态及CaMKⅡ和CREB活性的影响
宋杨张海燕1孙雅萍郭庆国2孟欣2王彪2
(沈阳市第四人民医院,辽宁沈阳110120)
摘要〔〕目的通过研究慢性铝暴露对大鼠海马钙离子稳态及CaMKⅡ和CREB活性的影响,探讨慢性铝暴露诱导神经损伤可能存在的机制。方法选用60只断乳后雄性Wistar大鼠,分为3组,每组20只;设定其中1组为对照组,用蒸馏水喂养;其他2组为铝暴露组,分别用含有0.2%和0.4% AlCl3的蒸馏水喂养3个月。Morris 水迷宫测定各组大鼠学习记忆能力;Fura-2/AM测定各组大鼠海马神经细胞自由Ca2+浓度变化;Western印迹方法检测各组大鼠海马中CaMKⅡ、Ng和CREB的蛋白表达情况。结果大鼠的学习记忆能力明显降低;与对照组相比,铝暴露各组海马神经细胞中〔Ca2+〕i呈剂量依赖性升高;CaMKⅡ和CREB总蛋白表达无显著变化,但p-CaMKⅡ、Ng和p-CREB蛋白表达显著降低(P<0.05)。结论慢性铝暴露损伤大鼠的学习记忆可能与上调Ca2+及抑制CaMKⅡ和CREB活性相关。
关键词〔〕铝;钙;钙调蛋白依赖的蛋白激酶Ⅱ;cAMP反应元件结合蛋白;海马
1中国医科大学附属第一医院老年病教研室
2中国医科大学基础医学院生物化学与分子生物学教研室
第一作者:宋杨(1981-),女,硕士,主治医师,主要从事神经系统疾病研究。
铝(Al)是一种慢性神经毒性物质,随着铝的生物利用度增加,其与许多神经变性疾病密切相关,包括阿尔茨海默病(AD)、肌萎缩性侧索硬化症(ALS)、帕金森痴呆和海湾战争综合征〔1,2〕。动物实验证明,Al盐可引起许多动物脑内形成神经纤维缠结(NFT)病理变化,如兔〔3〕、猫〔4〕、小鼠〔5〕、大鼠〔6〕和猴〔7〕。最新的研究结果也证明,Al暴露可导致脑内产生AD样病理变化以及认知障碍〔8,9〕,并且饮水中Al含量与AD患者死亡率呈正相关〔10〕。慢性Al暴露可以增强AD患者脑内NFT和Aβ的形成,从而增加淀粉样斑块的体积和数量〔11〕。在神经元中,钙离子(Ca2+)持续性升高可造成神经毒性,与急慢性神经退行性病变中的神经损伤密切相关〔12〕。神经元内的Al可以影响静息Ca2+水平,减慢Ca2+流出细胞质〔13〕。同时,Al可以抑制调节Ca2+的相关蛋白表达。因此,了解铝暴露对Ca2+稳态的影响对于明确铝神经毒性的机制,具有十分重要的意义。Ca2+/钙调蛋白(CaM)依赖的蛋白激酶Ⅱ(CaMKⅡ)是神经细胞兴奋性转录耦联的关键媒介,也是突触可塑性改变的关键蛋白〔14〕。此外,cAMP 反应元件结合蛋白(CREB)是CaMKⅡ的下游分子之一,在突触可塑性中具有重要作用,激活CREB可促进多种与突触可塑性相关的转录因子的转录〔15〕,从而诱导产生一些与学习记忆十分相关的蛋白分子。研究表明,Al在脑内特定的敏感区域内选择性地沉积〔16〕,如海马区等。但是,慢性Al暴露对大鼠海马区神经元内的Ca2+稳态以及其下游CaMKⅡ、CREB信号分子活性的影响尚未见报道。本文通过对慢性Al暴露影响大鼠海马神经元Ca2+及其下游CaMKⅡ、CREB信号分子的观察,探讨Al诱导的神经损伤的可能机制。
1材料与方法
1.1主要材料选择60只断乳后Wistar大鼠(体重约30 g),由中国医科大学实验动物中心提供。CREB抗体、磷酸化(p)-CREB抗体(Cell Signal公司);CaMKⅡ抗体、p-CaMKⅡ抗体、神经颗粒素(Ng)抗体(美国SANTA Cruz公司)、β肌动蛋白(β-actin)抗体、辣根过氧化物酶(HRP)标记的二抗(北京中杉生物公司);ECL发光试剂(美国Sigma公司)、Chemilmager 5500 V2103(美国)。其他试剂均为分析纯。
1.2动物模型建立将60只(AlCl3)大鼠随机分3组,每组20只。对照组用蒸馏水喂养,Al暴露组分别用含有0.2%和0.4%氯化铝的蒸馏水喂养。动物室温度保持在18℃~23℃、相对湿度为45%~55%,光照时间12 h∶12 h。Al暴露大鼠保持健康,体重为对照大鼠的(90±6)%。3个月后,进行大鼠学习记忆的行为学及其他生化指标测定。
参考文献1.3Morris 水迷宫测定大鼠学习记忆行为〔17〕的方法,Morris水迷宫测试在内径180 cm、高60 cm的圆形水池中进行。水深48 cm,水温为(22±0.5)℃,水池内放入奶粉使之成为乳白色。将水池分为西南、西北、东南和东北(SW、NW、SE和NE)四个象限,NW象限正中距池壁20 cm处放置逃避平台(直径约8 cm),平台位于水下2 cm。水池正上方安装带有显示系统的摄像机,计算机自动跟踪计时并记录游泳轨迹,实验期内迷宫外参照物保持不变。Morris水迷宫实验包括定位航行实验和空间探索实验两个部分,根据相关文献〔17〕,简单地说,实验大鼠每天游泳4次,分别从不同象限将动物头朝池壁入水,并在水池上标定相同一点作为每次实验大鼠的入水点,寻找平台时间上限设置为120 s。如果在规定时间内未找到平台,由实验者将其引导至平台并停留30s以加强记忆,缓解紧张情绪。逃避潜伏期记录为120 s,每只大鼠的游泳时间间隔为15 min。观察和记录120 s内大鼠穿越平台的次数、在原平台象限搜索时间占总时间的百分比和在原平台象限搜索距离占总距离的百分比,并记录大鼠在120 s内搜索平台的路线图。空间探索实验只进行1次。 4
1.4脑组织和血液中Al含量测定根据文献〔18,19〕,准确分离并称取各组大鼠海马组织(约0.1~0.5 g)或准确吸取0.2~0.5 ml的全血于石英烧杯中,放入聚四氟乙烯(PTFE)中,加入5~8 ml混酸(硝酸∶高氯酸体积比为4∶1),同时做空白对照。低温加热至试样全部溶解,继续加热蒸发至近干后加0.2%硝酸溶解残留物,并以0.2%硝酸定容至50 ml,采用石墨炉原子吸收分光光度法测定脑铝或血铝含量。
1.5大鼠海马神经细胞内Ca2+含量测定〔20〕冰上分离大鼠海马组织后,放入D-Hank液中。37℃下用0.125%胰酶消化20 min,反复吹打后离心,制备密度为106~107个/ml的单细胞悬液。台盼蓝染色,确定分离出的单细胞存活率在95%以上。将终浓度为4 μmol/L的Fura-2/AM加入上述分离出来的细胞悬液中,37℃下孵育35 min后,用含0.2% BSA的Hank液清洗残留的Fura-2/AM。然后,将细胞重悬于无Mg2+的Hank液中(mmol/L):HEPES 10,KCl 5.0,NaCl 145,CaCl21.3,L-glucose 10,Na2HPO4。采用日立F-3000型荧光双波长分光光度计测定细胞Ca2+浓度。激发光波长设定为340~380 nm,发射光波长为510 nm。根据下面公式计算Ca2+浓度〔Ca2+〕i=Kd〔(R-Rmin)/(Rmax-R)〕(Fmin/Fmax)。Kd值为224 nmol/L,R为荧光值,〔Ca2+〕i为nmol/L,而Fmin为加入EGTA后测得的荧光值,Fmax为加入TritonX-100后测得的荧光值。
1.6逆转录实时PCR法检测mRNA表达冰上分离各组大鼠海马组织,按照TRIzol说明书(Invitrogen,USA)抽提各组海马组织的总RNA。应用紫外分光光度仪测定RNA样品260/280 nm波长处吸光度比值为1.9~2.0。根据试剂盒说明书要求,采用oligo (dT)和super script Ⅱ进行逆转录后,用双标记荧光探针混合物(SYBR Green PCR Master mix)在ABI prism7500自动序列分析系统(Biosystems)中进行实时定量PCR分析,每组重复3次。CaMKⅡ基因引物特异序列分别为5′-AGCCATCCTCACCACTAT-3′(上游) 和5′-CTTCACGCCATCATTCTTC-3′(下游);CREB基因引物特异序列分别为:5′-CCAGCCATCAGTTATTCAGT-3′(上游)和5′-TTACACTATCCACAGACTCCT′(下游);β-actin 作为内参照,其引物特异序列为5′-GAG ACC TTC AAC ACC CCA GCC-3′(上游) 和5′-GGA TCT TCA TGA GGTA GTCA G-3′。两种基因扩增结果用β-actin进行标化。
1.7免疫印迹检测蛋白表达采用颈椎脱臼法处死大鼠,冰上分离大鼠海马,剔除血管。在每个装有海马的离心管中,按照1∶9比例加入裂解液(10 mmol/L Tris-HCl pH 7.5,1 mmol/L EDTA,1 mmol/L EGTA,100 mmol/L NaCl,50 mmol/L NaF,1% TritonX-100,1 mmol/L Na3VO4,1 mmol/L PMSF),1 μg/ml亮肽素,1 μg/ml抑蛋白酶 (Santa Cruz,USA),50 mmol/L β-glycerol phosphate,1 mmol/L二硫苏糖醇和0.09% Brij-35。4℃下超声粉碎后27 000 g离心1.5 h,取上清,即总蛋白提取液。BCA蛋白定量试剂盒测定样品蛋白浓度,取50 μg总蛋白,采用10% SDS-聚丙烯酰胺凝胶进行电泳(110 V,2 h)。待电泳完成后,把蛋白转印至硝酸纤维素(NC)上。分别用抗tCREB、p-CREB (Ser133) (1∶100)、Ng(1∶500)、p-CaMKⅡ(1∶500)和β-actin (1∶1 000)抗体孵育,最后加入HRP标记的相应二抗,ECL发光并成像分析。
1.8免疫组化检测蛋白表达各组大鼠经4%多聚甲醛灌流固定后取海马组织,30%蔗糖(用4%多聚甲醛配置)后固定48 h,冷冻切片 (40 μm/片 )。分别经0.3%过氧化氢甲醇溶液、0.3% Triton-100处理后,0.01 mol/L PBS清洗5 min×3次。加入一抗CaMKⅡ(1∶200)4℃孵育过夜、二抗(1∶1 000)室温下孵育2 h、DAB法显色。
1.9统计学分析应用SPSS11.0软件,组间资料统计用单因素方差分析 (ANOVA)。采用单因素重复测量方差分析评估行为学数据。
2结果
2.1慢性Al暴露可诱导大鼠学习记忆损伤Morris水迷宫测试结果表明,与对照组相比较,铝暴露各组大鼠的穿越平台次数、在原平台象限搜索时间占总时间的百分比和在原平台象限搜索距离占总距离的百分比显著降低(P<0.05);在慢性Al暴露各组之间,0.4% AlCl3组大鼠穿越平台次数和搜索距离明显低于0.2% AlCl3组(P<0.05),呈现剂量依赖性;各组搜索时间无统计学差异。见表1。
2.2慢性Al暴露可增加大鼠血Al和海马组织内Al的含量与对照组相比较,Al暴露各组大鼠的血液和海马组织中Al含量明显增加(P<0.05);在Al暴露各组大鼠之间,0.4% AlCl3组大鼠的血Al和海马组织Al含量明显高于0.2%组(P<0.05)。见表2。

表1 Morris 水迷宫测试结果
与对照组相比:1)P<0.05;与0.2% AlCl3组相比:2)P<0.05;下表同
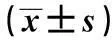
表2 各组大鼠血Al和脑Al的比较
血清Al含量测定:n= 10,海马组织Al含量测定:n=6
2.3慢性Al暴露可增加大鼠海马神经细胞内的自由钙离子(〔Ca2+〕i)浓度与对照组相比,Al暴露各组大鼠海马神经细胞内〔Ca2+〕i明显增高,从(155.75 ± 15.33)nmol/L(对照组)分别增至(515.92 ± 53.59)nmol/L(0.2% AlCl3组)和(646.87 ± 62.32)nmol/L(0.4% AlCl3组);与0.2% AlCl3组相比,0.4% AlCl3组大鼠海马神经细胞内〔Ca2+〕i显著升高(P<0.05)。
2.4慢性Al暴露对CaMKⅡ和CREB mRNA表达的影响各组CaMKⅡ及CREB mRNA表达无显著变化(P>0.05)。对照
组设为1,0.2% AlCl3组与0.4% AlCl3组CaMKⅡ mRNA表达量分别为0.98±0.21、1.11±0.18,CREB mRNA表达量分别为0.99±0.19、1.03±0.15。
2.5慢性Al暴露对CaMKⅡ和CREB总蛋白表达的影响免疫组化结果显示,各组CaMKⅡ染色密度元显著差异(P>0.05)。同时,Western印迹结果也显示(图1),各组CREB总蛋白表达水平无显著变化(P>0.05),CREB总蛋白表达分别为0.55 ± 0.065(对照组)、0.54 ± 0.062(0.2% AlCl3组)和0.57 ± 0.064(0.4% AlCl3组)(P>0.05)。
2.6慢性Al暴露影响学习记忆关键蛋白分子CaMKⅡ和CREB的活性与对照组相比,Al暴露组大鼠海马组织中p-CaMKⅡ、Ng和p-CREB表达水平均明显降低(P<0.05);同时,随着Al暴露浓度的增加,p-CaMKⅡ、Ng和p-CREB表达逐渐降低,呈剂量依赖性(P<0.05)。见图2。

图1 慢性Al暴露对CaMKⅡ和CREB总蛋白表达的影响

与对照组相比:1)P<0.05;与0.2% AlCl3组相比:2)P<0.05图2 慢性Al暴露对大鼠海马神经元p-CaMKⅡ、Ng和p-CREB表达的影响
3讨论
Al暴露因素是AD发病机制中研究最多的环境风险因素。Al在神经系统中的蓄积可产生毒性作用,导致神经元损伤,从而引起智力和认知能力的缺欠〔21〕。同时,AD患者脑Al含量较高,神经原纤维缠结(NTFs)中含有高浓度的Al。因此,研究Al暴露与AD的关系十分重要。
Morris水迷宫行为学评估结果证明,慢性Al暴露确实可以造成大鼠的学习记忆损伤,这些结果与以往国内外一些研究的结果相似〔22〕。
Ca2+对真核细胞的代谢及各种生理功能具有十分重要的作用,并且细胞外Ca2+浓度远远高于细胞内Ca2+浓度。此外,在神经系统中,Ca2+作为最基本的第二信使,可以调节神经递质的释放、调控突触可塑性,甚至是基因表达〔23〕。有研究表明,Al3+离子半径与Ca2+相似,可以与Ca2+结合位点作用,从而在一定程度上影响细胞内的钙稳态〔24〕。在体外培养的条件下,Al暴露后可显著提高星形胶质细胞内Ca2+浓度〔25〕。我们的实验结果证明,慢性Al暴露可显著提高大鼠海马神经细胞内的Ca2+浓度,并呈剂量依赖性。此结果与慢性Al暴露可显著增加神经突触内钙浓度的研究结果〔24〕相似。
CaMKⅡ是空间学习和记忆的分子基础,Ca2+内流可磷酸化激活CaMKⅡ,活化的CaMKⅡ从细胞质移动到突触并结合NMDA型谷氨酸受体,而CaMKⅡ-NMDA受体复合物是诱导长时程增强的关键分子〔26,27〕。研究表明,在LTP后期,CaMKⅡ具有放大并强化突触可塑性的结构性功能〔28〕。我们的结果证明,慢性Al暴露并未影响大鼠海马区CaMKⅡ总蛋白的表达,但是却降低了p-CaMKⅡ的表达,说明Al暴露可抑制CaMKⅡ的活性。
Ng是一种突触后蛋白,可以与CaM单体结合〔29〕,对于CaMKⅡ的活性具有重要的影响。研究表明,Ng增高可以增加CaMKⅡ激活,从而增强突触可塑性;而在Ng敲除的小鼠中,CaMKⅡ的活性也明显降低〔30〕。我们的结果表明,Al暴露可以降低大鼠海马神经细胞中Ng的蛋白表达。因此,总体来说,慢性Al暴露干扰了大鼠海马神经细胞的钙稳态,降低了p-CaMKⅡ和Ng的蛋白表达,这可能是Al暴露诱导学习记忆损伤、诱导AD发病的机制之一。
CREB是一种分子量为43 kD的核转录因子蛋白,对于神经突触可塑性长时间的改变具有十分重要的作用〔31〕。CREB上的丝氨酸(Ser)133是许多蛋白激酶的作用靶点,例如CaMKⅡ和Ⅳ、蛋白激酶A和蛋白激酶C等。一旦Ser-133位点被磷酸化,CRE介导的转录即刻启动,从而最终激发LTP相关蛋白的合成。本结果证明,慢性Al暴露显著抑制p-CREB(S133)的表达,但是CREB总蛋白表达未受到显著影响。这可能是由于p-CaMKⅡ蛋白的降低抑制了CREB(S133)磷酸化过程。综上所述,本文结果表明慢性Al暴露损伤了大鼠的学习记忆能力,这可能是由于Al暴露造成了海马神经细胞内钙离子紊乱,从而抑制了CaMKⅡ/CREB通路。除此之外,Al暴露还抑制了海马神经细胞的Ng蛋白表达,进一步抑制了CaMKⅡ的激活。
1Tomljenovic L.Aluminum and Alzheimer's disease:after a century of controversy,is there a plausible link〔J〕? J Alzheim Dis,2011;23(4):567-98.
2Petrik MS,Wong MC,Tabata RC,etal.Aluminum adjuvant linked to Gulf War illness induces motor neuron death in mice〔J〕.Neuromol Med,2007;9(1):83-100.
3Perl DP, Moalem S.Aluminum and Alzheimer's disease,a personal perspective after 25 years〔J〕.J Alzheimer Dis:JAD,2006;9(3 Suppl):291-300.
4Perl DP, Good PF.The association of aluminum Alzheimer's disease,and neurofibrillary tangles〔J〕.J Neural Trans Suppl,1987;24:205-11.
5Games D,Adams D,Alessandrini R,etal.Alzheimer-type neuropathology in transgenic mice overexpressing V717F beta-amyloid precursor protein〔J〕.Nature,1995;373(6514):523-7.
6Walton JR.Functional impairment in aged rats chronically exposed to human range dietary aluminum equivalents〔J〕.Neurotoxicology,2009;30(2):182-93.
7Uno H,Alsum PB,Dong S,etal.Cerebral amyloid angiopathy and plaques,and visceral amyloidosis in aged macaques〔J〕.Neurobiol Aging,1996;17(2):275-81.
8Exley C, Esiri MM.Severe cerebral congophilic angiopathy coincident with increased brain aluminium in a resident of Camelford,Cornwall,UK〔J〕.J Neurol Neurosurg Psychiatry,2006;77(7):877-9.
9Colomina MT,Roig JL,Sanchez DJ,etal.Influence of age on aluminum-induced neurobehavioral effects and morphological changes in rat brain〔J〕.Neurotoxicology,2002;23(6):775-781.
10Yumoto S,Kakimi S,Ohsaki A,etal.Demonstration of aluminum in amyloid fibers in the cores of senile plaques in the brains of patients with Alzheimer's disease〔J〕.J Inorganic Biochem,2009;103(11):1579-84.
11Walton JR.Cognitive deterioration and associated pathology induced by chronic low-level aluminum ingestion in a translational rat model provides an explanation of Alzheimer's disease,tests for susceptibility and avenues for treatment〔J〕.Intern J Alzheimer Dis,2012;2012:914947.
12Akasofu S,Kimura M,Kosasa T,etal.Protective effect of donepezil in primary-cultured rat cortical neurons exposed to N-methyl-d-aspartate (NMDA) toxicity〔J〕.Eur J Pharmacol,2006;530(3):215-22.
13Walton JR.Aluminum disruption of calcium homeostasis and signal transduction resembles change that occurs in aging and Alzheimer's disease〔J〕.J Alzheimer's Dis:JAD,2012;29(2):255-73.
14Giese KP,Fedorov NB,Filipkowski RK,etal.Autophosphorylation at Thr286 of the alpha calcium-calmodulin kinase Ⅱ in LTP and learning〔J〕.Science,1998;279(5352):870-3.
15Porte Y,Buhot MC, Mons NE.Spatial memory in the Morris water maze and activation of cyclic AMP response element-binding (CREB) protein within the mouse hippocampus〔J〕.Learning Mem,2008;15(12):885-94.
16Flaten TP.Aluminium as a risk factor in Alzheimer's disease,with emphasis on drinking water〔J〕.Brain Res Bulletin,2001;55(2):187-96.
17宗志红,王彪.慢性铝暴露对雄性大鼠海马组织中TLR2蛋白及其mRNA表达的影响〔J〕.环境与健康杂志,2011;28(3):208-11.
18王军明,宋立伟,史立新,等.塞曼石墨炉原子吸收光谱法直接测定职业接触者血液中铝〔J〕.中国卫生检验杂志,2006;16(2):210-1.
19Wang B,Xing W,Zhao Y,etal.Effects of chronic aluminum exposure on memory through multiple signal transduction pathways〔J〕.Environ Toxicol Pharmacol,2010;29(3):308-13.
20周平,唐秋实,靳翠红,等.慢性染铝对大鼠海马长时程增强及Ca2+浓度影响〔J〕.中国公共卫生,2009;25(12):1497-8.
21Yokel RA.The toxicology of aluminum in the brain:a review〔J〕.Neurotoxicology,2000;21(5):813-28.
22王彪,赵伟,周丽坤,等.慢性铝暴露对大鼠海马神经元中toll样受体4蛋白及其mRNA表达的影响〔J〕.中华神经科杂志,2010;43(3):186-90.
23Raheja G,Gill KD.Calcium homeostasis and dichlorvos induced neurotoxicity in rat brain〔J〕.Mol Cell Biochem,2002;232(1-2):13-8.
24Kaur A, Gill KD.Disruption of neuronal calcium homeostasis after chronic aluminium toxicity in rats〔J〕.Basic Clin Pharmacol Toxicol,2005;96(2):118-22.
25Guo GW, Liang YX.Aluminum-induced apoptosis in cultured astrocytes and its effect on calcium homeostasis〔J〕.Brain Res,2001;888(2):221-6.
26Barria A, Malinow R.NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKⅡ〔J〕.Neuron,2005;48(2):289-301.
27Gardoni F,Polli F,Cattabeni F,etal.Calcium-calmodulin-dependent protein kinase Ⅱ phosphorylation modulates PSD-95 binding to NMDA receptors〔J〕.Eur J Neurosci,2006;24(10):2694-704.
28Lisman J,Yasuda R,Raghavachari S.Mechanisms of CaMKⅡ action in long-term potentiation〔J〕.Nature Rev Neurosci,2012;13(3):169-82.
29Reddy AS,Ben-Hur A,Day IS.Experimental and computational approaches for the study of calmodulin interactions〔J〕.Phytochemistry,2011;72(10):1007-19.
30Huang KP,Huang FL,Jager T,etal.Neurogranin/RC3 enhances long-term potentiation and learning by promoting calcium-mediated signaling〔J〕.J Neurosci,2004;24(47):10660-9.
31Paramanik V, Thakur MK.Role of CREB signaling in aging brain〔J〕.Arch Ital de Biol,2013;151(1):33-42.
〔2014-09-15修回〕
(编辑徐杰)
·基础研究·
Effect of chronic aluminum exposure on calcium homeostasis and activities of CaMKⅡ and CREB in hippocampus of rats
SONG Yang, ZHANG Hai-Yan, SUN Ya-Ping,etal.
Shenyang The Fourth Hospital of People, Shenyang 110120, Liaoning, China
【Abstract】ObjectiveTo study the potential mechanism of aluminum-induced memory injury through the research on effect of chronic aluminum exposure on calcium homeostasis and activities of CaMKⅡ and CREB in hippocampus of rats.Methods60 weaning rats were divided into 3 groups, which were given distilled water containing 0.2% AlCl3and 0.4% AlCl3respectively for 3 months to establish the chronic aluminum exposure model. Morris water maze was used to evaluate the learning and memory ability; the freedom Ca2+in hippocampal neurons of rats was measured with Fura-2/AM; Western blot was used to determine the expressions of CaMKⅡ, Ng and CREB.ResultsLearning and memory abilities of rats were significantly reduced; the expressions of p-CaMKⅡ, Ng and p-CREB in the hippocampus of aluminum-treated groups were significantly lower than those in control group(P<0.05), but there were no changes in the expressions of total CaMKⅡ and CREB; and the aluminum-treated groups had the higher Ca2+level than that of control group(P<0.05).ConclusionsChronic aluminum exposure could impair the ability of learning and memory of rats, which is closely related to the up-regulation of Ca2+level and the down-regulation of CaMKⅡ and CREB activities.
【Key words】Aluminum; Calcium; Calmodulin-dependent protein kinase Ⅱ; cAMP response element binding protein; Hippocampus
基金项目:国家自然科学基金项目(81271292);辽宁省教育厅基金(2008848);沈阳市科技局资助项目(F13-220-9-44)
中图分类号〔〕R749〔
文献标识码〕A〔
文章编号〕1005-9202(2015)21-6001-05;doi:10.3969/j.issn.1005-9202.2015.21.001
