论鸟类飞行起源的“树栖”与“地栖”假说之争
2016-01-26季燕南王旭日黑须球子
季燕南,王旭日,黑须球子,季 强
(1.中国地质环境监测院,北京100081; 2.中国地质科学院地质研究所,北京100037; 3.中国地质大学(北京),北京100083)
论鸟类飞行起源的“树栖”与“地栖”假说之争
季燕南1,王旭日2,黑须球子3,季强2
(1.中国地质环境监测院,北京100081; 2.中国地质科学院地质研究所,北京100037; 3.中国地质大学(北京),北京100083)
摘要:简要介绍了鸟类飞行起源的2种假说:“树栖起源”假说与“地栖起源”假说。详细讨论了2种假说的理论支撑点及存在的问题,认为鸟类的祖先和原始鸟类可以借助自然界中任何有利的地形和地物来帮助滑翔和起飞,而不是仅仅依赖直立高大的树木。“地栖起源”假说更合乎情理,也更接近或更符合客观事实,同时提出了“地栖起源”假说的新模式图。
关键词:鸟类飞行起源;“树栖”假说;“地栖”假说;“翼助斜坡奔跑”假说
中图分类号:Q915.5;Q915.865
文献标识码:A
文章编号:1674-3636(2015)02-0201-06
收稿日期:2015-04-26;编辑:陆李萍
基金项目:科学技术部“973”项目(2012CB822004);中国地质调查局地质调查项目(121202114026801)
作者简介:季燕南(1983—),男,工程师,硕士,主要从事地层古生物和地质遗迹保护研究工作,E-mail:jirod@cags.ac.cn
doi:10.3969/j.issn.1674-3636.2015.02.207
0引言
中国辽西是晚中生代的化石宝库,以产出丰富的长羽毛恐龙闻名于世。自1996年发现中华龙鸟以来,鸟类起源研究取得了革命性进展,“鸟类是由小型兽脚类恐龙演化而来”的观点已成为共识,也成为当今国际科学界的主流科学思想。
鸟类的飞行起源与鸟类起源是2种性质完全不同但又有一定关系的科学问题。长期以来,关于鸟类的飞行起源存在2种观点对立、解释相背的理论,即“树栖起源”假说与“地栖起源”假说。
脊椎动物中主要有3种具有翅膀、会飞行的动物:爬行动物中的翼龙、鸟类和哺乳动物中的蝙蝠,它们的翅膀结构各不相同。翼龙的前肢十分发育,一般具有4指,第5指缺失,其第4指(飞行指)极度拉长,与皮膜一起形成翅膀,从而具有飞行能力。蝙蝠的前肢发育5指,第1指短小,其余4指明显拉长,似“伞骨”状,与皮膜一起形成翅膀,从而具有了飞行能力(图1)。鸟类的前肢比较特殊,通过手部骨骼的愈合和消失等变化与羽毛一起形成翅膀。粗壮的肱骨构成上臂;细长的尺骨和桡骨构成前臂;腕骨退化,近端腕骨仅有2块,远端腕骨与掌骨愈合成1块腕掌骨;指骨仅发育第2、3、4指,第2指为小翼羽的着生处。鸟类翅膀的这种结构究竟与“爬树”有关,还是与其他因素有关?本研究将简要介绍3种主要假说,并讨论鸟类飞行起源各种可能的原因。

图1 翼龙、鸟类和蝙蝠翅膀的形态结构对比图Fig.1 Comparison of morphological structures of pterosaurs, birds and bat wings
1“树栖起源”假说
“树栖起源”假说最早由Marsh(1880)提出,当时主要受到松鼠、飞蜥、树蛙等动物可以进行短距离滑翔现象的影响和启迪。他推测鸟类的祖先或早期鸟类的前肢可能发育了雏形羽毛,经过长期的自然选择,增强了它们在树林间跳跃时的滑翔能力,雏形羽毛不断进化,演变成飞行羽毛,最终获得了飞行能力。这一假说后来得到了许多学者的认可和支持,其中Heilmann(1926)还给出了鸟类的祖先利用四肢攀爬树枝的生态复原图(图2),表明鸟类的祖先具有利用四肢攀援树枝和滑翔的能力。Bock(1969,1986)进一步发展了这种假说。他认为鸟类的祖先应是地栖爬行动物,经过长期的自然选择,进化为两足行走、可以攀援树干的地栖原始鸟类。原始鸟类一开始在树枝间进行短距离跳跃,然后进行稍远距离的跳跃,从而逐渐获得了滑翔能力,进而可以从高树滑翔到矮树,甚至滑翔到地面,最终具有了远距离主动飞行的能力。Xu等(2000)报道了产于我国辽西早白垩世义县组的一种小型奔龙化石——赵氏小盗龙(图3)。通过研究,认为赵氏小盗龙是一种可以振翅飞行并且栖居树上的小型恐龙,有力支持了“树栖起源”假说。

图2 “树栖起源”假说生态复原图(据Heilmann,1926)Fig.2 Ecological restoration plan of the “arborealorigin” hypothesis(after Heilmann,1926)

图3 赵氏小盗龙生态复原图(据Xu等, 2000)Fig.3 Ecological restoration plan of the Microraptorzhaoianus(after Xu et al, 2000)
2“地栖起源”假说
“地栖起源”假说亦称“陆地奔跑起源”假说,最早由Williston(1879)首次提出。他认为两足行走的恐龙在快速奔跑过程中,由辅助性的捕食、抓握和平衡功能的前肢逐渐拉长,演变为具有羽毛的翅膀,进而获得了滑翔乃至飞行的能力。Nopsca(1907,1923)撰文支持这种假说,认为鸟类起源于两足疾走的长尾爬行动物,鸟类的祖先或原始鸟类在快速奔跑时,其前肢是辅助后肢加快奔跑速度的“推动器”。此后,Ostrom(1974,1976,1979)、Padian(1982,1985)等通过对始祖鸟骨骼解剖的研究,进一步完善了“地栖起源”假说,提出“昆虫网捕”理论,认为脚趾的对握与否是判别鸟类的祖先或鸟类是否具有树栖能力的重要特征(图4)。他们认为始祖鸟的后肢非常发育,胫骨长,跗跖骨与股骨的长度比为2∶1,这与奔跑的机理结构相吻合。始祖鸟的第1趾爪(母趾爪)没有特别拉长,明显短于其他趾爪,而且其所有趾爪都不强烈弯曲,也不锐利,屈肌突小,这些特征更接近于现生地栖鸟类的趾爪,因而始祖鸟的脚不具有抓握或树栖功能。Maye等(2005)对新标本的研究也证明始祖鸟的第1趾没有反转,不具有抓握或树栖功能。季强等(2002)报道了1件产自中国辽西早白垩世义县组的初鸟类化石——中华神州鸟。认为中华神州鸟具有较强的飞行能力,但其第1趾(母趾)没有反转,与其他趾的方向相同,表明其具有陆地奔跑能力,而不具有抓握或树栖功能,有力地支持了“地栖起源”假说。

图4 “地栖起源”假说中的昆虫网捕理论示意图(据Currie,1991)Fig.4 Sketch showing the insect net theory of the“terrestrial origin” hypothesis(after Currie, 1991)
3斜坡起源假说
“斜坡起源”假说系Dial(2003)首次提出,主要依据实验室中观察到的一些有趣的现象。将一些尚不具备飞行能力的幼鸟放在不同角度的斜板上反复进行试验,观察它们在不同条件下的反应。结果表明,这些幼鸟在30°的斜板上可以轻松地利用后肢从底部跑到顶部;在45°的斜板上仍可以利用后肢从底部跑到顶部;在60°的斜板上利用后肢跑显得有些吃力,开始扇动翅膀,困难地从底部跑到顶部;在大于70°的斜板上根本就跑不上去。不过,当将一些成鸟放在大于70°的斜板上时,它们就会利用翅膀的扇动辅助后肢从底部“跑”到顶部(图5)。

图5 “斜坡起源”假说示意图(据李志恒等,2008)Fig.5 Sketch showing the “slope origin” hypothesis(after Li et al, 2008)
4问题讨论
通过上述介绍可以看出,“树栖起源”假说特别强调鸟类的祖先或原始鸟类最初必须具有爬树能力,因为树木的高度为“滑翔”或“飞行”提供了看似合乎情理的客观条件,进而认为始祖鸟后肢的母趾与其他3趾相对是适于攀援、抓握的有力证据(实际上最新的研究证明始祖鸟的母趾并没有反转,也没有树栖能力)。表面看来,“树栖起源”假说似乎比较合乎情理,但细究之下有许多方面令人难以理解。鸟类的祖先为什么非要爬树不可?它们爬树的动机是什么?是为了捕食还是为了躲避敌害?
首先,如果说这种假说的支撑点是产生滑翔所需的“高差”的话,高大直立的树木是一种可能,但产生滑翔所需的“高差”在自然界中随处可见,如土堆、陡坎、山崖、斜倒的树干等均可形成高差。也就是说,高大直立的树木并不是产生“高差”的唯一可能,特别强调“具有爬树能力”就显得太牵强附会了。“爬树”是众多可能条件中的一种条件,但绝不是必要的充分条件。
其次,鸟类的祖先是小型食肉性恐龙,食物来源一般是地面生活的小型动物,如哺乳动物、爬行动物以及昆虫等,它们不可能放弃地面上轻易可得的食物,非要上树寻找其他食物不可。显然,为了捕食而上树看来不能作为支撑这种假说的主要原因。
再者,鸟类的祖先是小型食肉性的恐龙,它们处于食物链的顶层,是捕食者,一般利用它们的锐利的牙齿和强壮的后肢去攻击敌害或捕食小动物。偶尔遇到比它们强大的敌害的情况可能会发生,慌不择路偶尔爬树躲避敌害的可能是有的,但绝不是普遍常见的现象。这也表明,鸟类的祖先为了躲避敌害而上树的理由显得很不充分。
还有,从骨骼解剖学的角度来看,无论是奔龙还是伤齿龙,它们的后肢长而强壮,明显长于前肢,除第2趾爪外,其余的趾爪均不强烈弯曲。这些特征表明,它们是善于快速奔跑的捕食者。季强等(2003)专文讨论了鸟类飞行的起源,认为鸟类的祖先和原始鸟类在一定阶段可能具有一定的(有限的)“爬树”能力,但并不意味着“爬树”是获得飞行能力的“必由之路”,它们可以利用自然界中的各种有利条件(地形、地物)来帮助它们滑翔和起飞。李志恒等(2008)也专文讨论了鸟类飞行起源问题,认为鸟类飞行起源的进化途径只有3种:地面进化、树上进化和斜坡进化。笔者认为,简单地把飞行起源进化归为3种是不科学的,也不符合客观实际情况,进化途径应该是多种多样的。
“地面进化”思想应当来自Williston(1879)提出的“陆地奔跑起源”假说(“地栖起源”假说)。当时仅仅是提出一个简单的构想和描述,此后该假说在相当长的一段时间内并没有引起人们太多的关注。美国学者Ostrom(1974,1976,1979)的研究对于该假说的复苏发挥了非常重要的作用,他详细研究了鸟类飞行中的关键动作“扇翅”的起源,提出了“昆虫网捕”理论并加以解释(图6)。笔者认为“昆虫网捕”理论成立与否并不重要,始祖鸟是一种陆地快速奔跑的两足猎食动物,而且不能在树上栖居,为什么还要将这种理论与“上树”联系在一起?
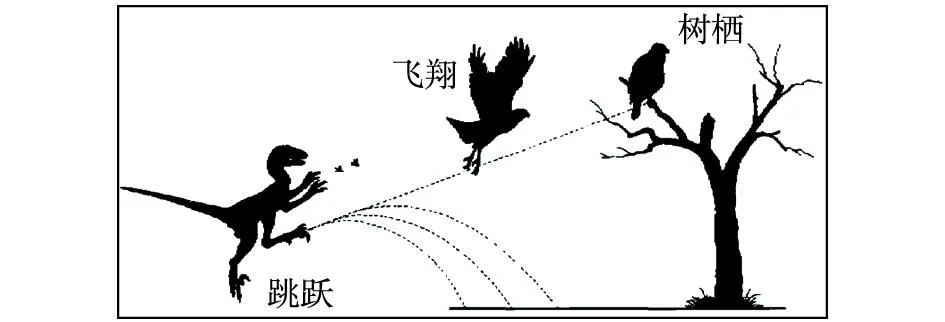
图6 “地栖起源”假说示意图(据李志恒等,2008)Fig.6 Sketch of the “terrestrial origin” hypothesis(after Li et al, 2008)
“树上进化”思想应当来自Marsh(1880)提出的“树栖起源”假说,此后得到了许多学者的接受和支持。李志恒等(2008)认为,赵氏小盗龙、宁城树栖龙、顾氏小盗龙等恐龙的发现是支持“树栖起源”假说的有力证据,它们具有明显的树栖特征,如相对较低的第1趾(母趾)位置,较大的趾爪弧度,倒数第2趾加长,相对长的前肢等(图7)。笔者认为,他们提出的这些特征在奔龙、疾走龙、伤齿龙等中是十分常见的,是快速奔跑的特征,而不是树栖的特征。以赵氏小盗龙为例:其后肢明显长于前肢,前肢的3指短小,指爪也不强烈弯曲;其第1趾(母趾)没有反转,位置较低,趾爪短且不强烈弯曲;其他各趾均显得细长,趾爪既不强烈弯曲也不锐利。这些特征均表明赵氏小盗龙更适于陆地奔跑,而不善于爬树(图8、图9、图10、图11、图12)。它的趾爪不具有抓握功能,又如何能在树上栖居?另一方面,如果赵氏小盗龙的前肢适于爬树,在攀爬的过程中势必会不断强化它的攀爬功能,那么又怎么可能导致它的前肢向鸟类的翅膀演变呢?
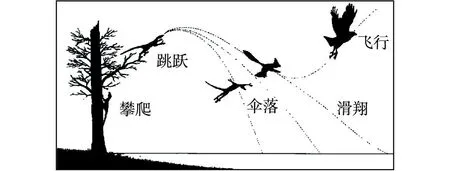
图7 “树栖起源”假说示意图(据李志恒等,2008)Fig.7 Sketch of the “arboreal origin” hypothesis(after Li et al, 2008)
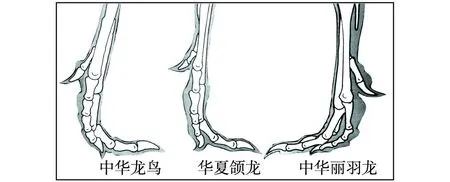
图8 美颌龙类恐龙脚趾形态结构图Fig.8 Morphological structures of toes of Compsognathus dinosaurs
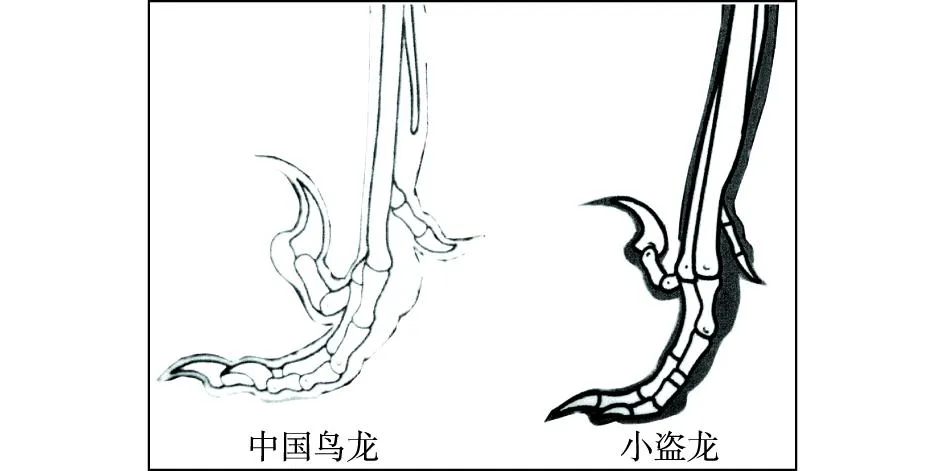
图9 奔龙类恐龙脚趾形态结构图Fig.9 Morphological structures of toes of Dromaeosaur dinosaurs
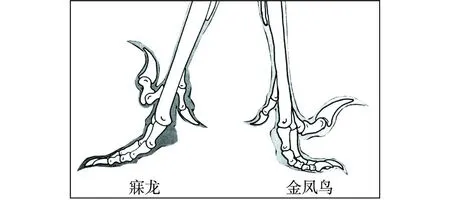
图10 伤齿龙类恐龙脚趾形态结构图Fig.10 Morphological structures of toes of Troodontids dinosaurs
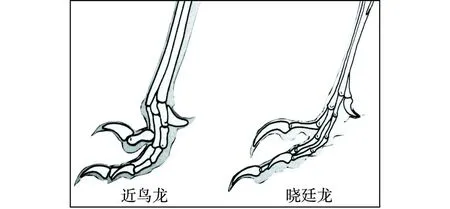
图11 伤齿龙类恐龙脚趾形态结构图Fig.11 Morphological structures of toes of Troodontids dinosaurs
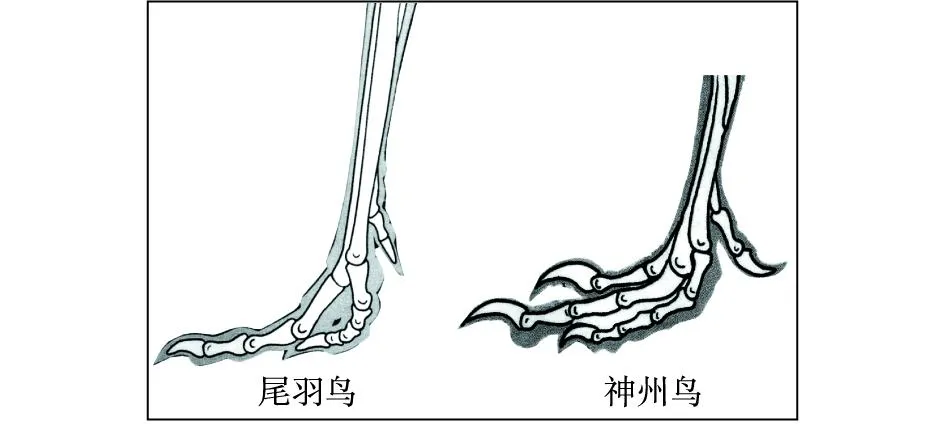
图12 窃蛋龙类恐龙与初鸟类脚趾形态结构图Fig.12 Morphological structures of toes of Oviraptorid dinosaurs and primitive birds
“斜坡进化”理论实际就是Dial(2003)提出的“斜坡起源”假说。有些人认为对于解释鸟类飞行起源具有潜在意义,因为具有早成性特征的鸟类祖先可能最初也经历过这种爬坡过程,而后获得了飞行能力。笔者不完全苟同于这些学者的看法,将个别特定的自然条件(如上树、爬坡等)来解释复杂的飞行起源问题显得局限、狭隘并带有浓厚的惯性思维色彩。实验结果恰恰表明,鸟类的祖先或原始鸟类在获得飞行能力之前主要是依赖后肢进行奔跑,为了加快奔跑速度或在一定的困难条件下,它们才利用前肢或翅膀扇动产生的推动力来帮助滑翔和起飞,同时也说明鸟类的祖先是两足奔跑的动物,具体来讲,就是小型食肉性恐龙。它们可以利用树木或斜坡,但不意味着必须依赖树木或斜坡来获得滑翔能力和飞行能力。
恐龙或爬行动物毕竟是动物,它们不可能具有人类的思维,更不可能在爬树之初就预想到几百万年后会长出羽毛,演变成鸟类而飞向天空。现在的研究表明,羽毛不是鸟类的专属特征,许多兽脚类恐龙早在鸟类之前就长出了羽毛。羽毛出现之初的主要功能是保持体温,是变温动物向恒温动物演变的必然结果。随着自身的发展演化,羽毛后来才逐渐具有了保护身体、运动平衡、性别展示、滑翔、飞行等功能。尽管现生鸟类的羽毛,特别是飞行羽毛,在飞行中发挥了重要作用,但现在还没有任何证据证明鸟类的祖先或原始鸟类羽毛的出现和早期演化与爬树有关。任何生物,包括恐龙和鸟类,保护自己的生命是首先必须想到的。恐龙的祖先在没有发育羽毛或羽毛不够发育的情况下是不会从高处向下跳的。相反,正是羽毛发育到一定的程度,足以保护自身的生命,它们才有可能从一定的高度向下滑落,而不至于摔伤或摔死。
目前,有关鸟类飞行起源的研究与讨论还没有结束,现有的研究材料还不足以对此问题过早得出结论。也许2种假说都有一定的道理,争论的焦点在于哪一种假说更合理,更具有代表性和普遍性。笔者认为,“地栖假说”更合乎逻辑,更有道理。鸟类的祖先在一定条件下,完全可以本能地借助自然界中的各种有利地形、地物来滑翔或起飞,如较大的岩石块体、土堆、斜坡、陡坎、山崖、倒卧的树干、低矮的灌木等,而不是仅仅依赖于直立高大的树木。后来有一天,它们突然发现自己展开双翅后,能够从高处向低处滑翔,继而进行短距离飞行,最终能够自由翱翔于蓝天与大地之间。为此,笔者给出了一个全新的“地栖起源”假说示意图(图13)。
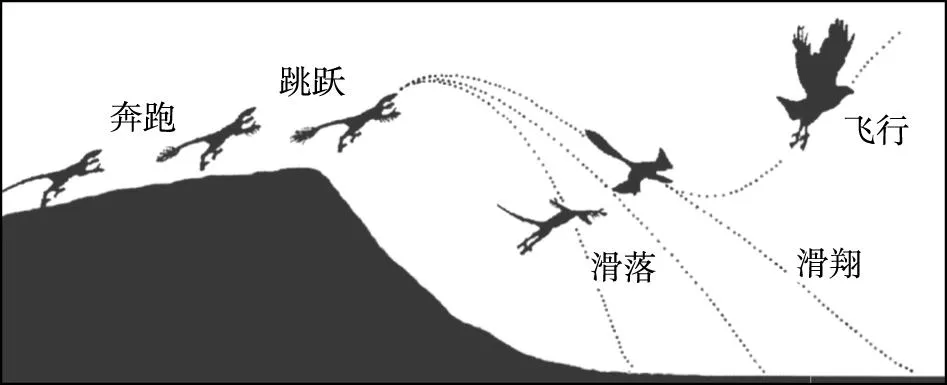
图13 新的“地栖起源”假说示意图Fig.13 Sketch showing the new “terrestrial origin”hypothesis
国际上早就在讨论鸟类起源、鸟类的飞行起源、四翼恐龙、羽毛的起源与演化等诸多问题,有些是肯定的意见,也有许多不同的声音,但国内知之甚少,究其原因主要是一些专家垄断了资源、信息和新闻,各种不同或反对的意见难以在国内见诸于世。若这种情况再继续下去,则会影响今后研究工作的开展,不利于我国科学技术的发展与进步。
2014年年末,《Science》评选出了2014年度“国际十大科技进展”,鸟类起源与鸟类飞行起源研究排名第二,文中的最后一句话十分耐人寻味:“尽管鸟类飞向了蓝天,但它们的飞行源自它们恐龙祖先的陆地奔跑(Birds took off, but their dinosaur ancestors had given them a running start.)”。
参考文献:
季强,姬书安. 1996.中国最早鸟类化石的发现及鸟类的起源. 中国地质,(10):30-33.
季强,姬书安.1997.原始祖鸟(Protarchaeopteryxgen. nov.):中国的始祖鸟类化石. 中国地质,(3):38-41.
季强,姬书安,尤海鲁,等.2002.中国首次发现真正会飞的“恐龙”:中华神州鸟(新属新种).地质通报,21(7):363-369.
季强,袁崇喜,季鑫鑫,等.2003.论鸟类飞行的起源. 地质论评,49(1):1-3.
李志恒,张玉光,周忠和.2008.鸟类飞行起源的研究.自然杂志,30(1):32-38.
郑光美.1995.鸟类学. 北京:北京师范大学出版社.
BOCK W J. 1969. The origin and radiation of birds. ANN N Y acad Sci, 167: 147-155.
BOCK W J. 1986. The arboreal origin of avian flight // PADIAN K. The origin of birds and the evolution of flight. San Francisco: Calif. Acad Sci,57-72.
BURGERS P, CHIAPPE L M.1999. The wing ofArchaeopteryxas a primary thrust generator . Nature, 399: 60-62.
CHATTERJEE S. 1997. The rise of birds . Baltimore, MD,USA:Johns Hopkins Univ Press.
CHIAPPE L M. 1997.ClimbingArchaeopteryx? A response to Yalden . Archaeopteryx, 15: 109-112.
DIAL K D. 2003.Wing-assisted incline running and the evolution of flight . Science, 299: 402-404.
FEDUCCIA A. 1993. Evidence from claw geometry indicatingarboreal habits ofArchaeopteryx. Science,259: 790-793.
FARLOW J O.2000.Theropod locomotion . American Zoologist,40: 640-663.
HEILMANN G. 1926. Origin of Birds .London,UK: Witherby.
JI QIANG.1998. Two feathered dinosaurs from northeasternChina . Nature, 393: 753-761.
JI QIANG. 2001. The distribution of integumentary structures in a feathered dinosaur . Nature, 410: 1084-1088.
LONGRICH N. 2006. Structure and function of hindlimb feathers inArchaeopteryxlithographica. Paleobiology,32 (3): 417-431.
MARSH O C. 1880. Odontornithes:A Monograph on the Extinct Toothed Birds of North America; With Thirty-four Plates and Forty Woodcuts. Washington, USA:Govt print off.
MAYE G, POHL B, PETERS D S A. 2005. A well-preservedArchaeopteryxspecimen with theropod features . Science, 310: 1483-1486.
NOPSCA F. 1907. Ideas on the origin of flight . Proc Zool Soc Lond,15: 223-236.
NOPSCA F. 1923. On the origin of flight in birds . Proc Zool Soc Lond, 31:463-477.
NORELL M A.2002.“Modern” feathers on a non-avian dinosaur. Nature, 416: 36-37.
OSTROM J.1974.Archaeopteryxand the origin of flight . Quart Rev Biol,49: 27-47.
OSTROM J.1976.Archaeopteryxand the origin of birds . Bio J Linnean Soc,8: 91-182.
OSTROM J. 1979. Bird flight: how did it begin? . American Scientist, 67: 46-56.
PADIAN K.1982.Running, Leaping, lifting off . The Sciences,10-15.
PADIAN K. 1985. The origin and aerodynamics of flight in extinct vertebrates . Palaeontology,28 (3): 413-433.
PADIAN K. 2003. Four winged dinosaurs, bird precursors, or neither? . Bio Science,53 (5): 450-452.
PADIAN K, DIAL K P. 2005. Origin of flight: could four-winged dinosaurs fly? . Nature,438: E3-E4.
WILLISTON S. 1879. Are birds derived from dinosaurs? .Kansas City Rev Sci, 3: 457-460.
XU XING, ZHOU ZHONGHE, WANG XIAOLIN. 2000. The smallest known non-avian theropod dinosaur . Nature, 408: 705-708.
YALDEN D W. 1997. ClimbingArchaeopteryx. Archaeopteryx, 15: 107-108.
A debate between the “arboreal” and “cursorial” hypotheses of the origin of bird flight
JI Yan-nan1, WANG Xu-ri2, Kurosu Mariko3, JI Qiang2
(1. China Geological Environmental Monitoring Institute, Beijing 100081, China; 2. Institute of Geology, Chinese Academy of Geological Sciences, Beijing 100037, China; 3. China University of Geosciences (Beijing), Beijing 100083, China)
Abstract:This paper briefly introduces two hypotheses of the origin of bird flight: the “arboreal” hypothesis and the “cursorial” (running)hypothesis, and discusses their theoretical bases and some problems in detail. It is considered that the ancestors and primitive birds could get their gliding ability and flight power with the help of favorable natural terrains, such as hill, hillock, slope, brae, cliff, steep ridge, huge boulder and inclined tree trunk, rather than only by the aid of erect tree trunk. It is suggested that the “running” hypothesis seems more reasonable, more close to, or, more coincident with the objective facts. Herein is given a new theoretical model of bird flight.
Keywords:bird flight; arboreal hypothesis; cursorial hypothesis; wing-assisted incline running hypothesis
