补体-中性粒细胞反馈的炎症应答调控在过敏性紫癜中的作用
2015-12-31周贵滨,温炬,李婷等
补体-中性粒细胞反馈的炎症应答调控在过敏性紫癜中的作用
周贵滨1温炬1李婷1秦思1
(南方医科大学第三临床医学院,广东广州510630)
摘要〔〕目的探讨补体-中性粒细胞反馈的炎症应答调控在过敏性紫癜(HSP)中的作用机制。方法选取24只清洁级日本大耳白兔为受试对象,分为正常组和模型组,每组12只。以干姜、荜茇、胡椒水煎剂混合液的药物灌胃,腹腔注射卵白蛋白-弗氏佐剂、耳缘静脉和背部皮内注射卵白蛋白生理盐水的方式构建过HSP模型。比较两组兔的饮食、饮水量、体温、血液常规和尿液常规变化及活性氧自由基(ROS)、炎性因子和补体成分的改变,同时以N-乙酰半胱氨酸(NAC)和过氧化氢(H2O2)干预ROS水平,并观察上述指标的变化。结果模型组兔在激发过敏后,体温明显升高,精神状态萎靡、运动量减少,饮水量和饮食量均较致敏前显著降低(P<0.05);而且,血液常规指标WBC、MCHC、NEU、NEU%水平明显升高,RBC、HGB、EOS和EOS%含量显著降低(P<0.05),而MCH和PLT则无明显变化(P>0.05);尿液出现拉丝、气味大的情况,其中66.67%出现尿蛋白阳性、58.33%出现尿红细胞、25%尿白细胞阳性;另外,模型组兔的ROS、炎性因子、TGF-β、补体水平也均明显升高;在ROS抑制剂NAC处理之后,上述指标水平明显降低(P<0.05),而在应用ROS激动剂H2O2处理之后,这些指标却明显升高(P<0.05)。结论补体-中性粒细胞反馈的炎症应答调控在HSP的形成中具有重要作用,抑制ROS水平能抑制HSP的发生和进展。
关键词〔〕过敏性紫癜;补体;中性粒细胞;炎症应答
中图分类号〔〕R55〔文献标识码〕A〔
通讯作者:温炬(1966-),男 ,博士,主任医师,教授,主要从事皮肤性病方面的研究。
1南方医科大学附属南粤医院广东省第二人民医院皮肤性病科
第一作者:周贵滨(1986-),男,在读硕士,主要从事皮肤性病方面的研究。
过敏性紫癜(HSP)又称许兰-亨诺紫癜,是一种常见的异质性、炎症性和自身免疫性皮肤血管炎症,其主要病理改变为真皮浅层的白细胞破碎、血管内皮细胞肿胀、中性粒细胞浸润等〔1〕。HSP多由外界变应原如食物、药物、细菌感染等多种致病因素的刺激引发〔2〕,但目前HSP的致病机制仍未阐明。免疫系统异常及免疫-炎症反应失调被认为是诱发HSP的主要潜在致病机制〔3,4〕。补体系统是机体抵御病原体入侵的主要效应系统,并且补体-中性粒细胞之间还存在一定的反馈炎症应答调控机制,即补体活化激活中性粒细胞呼吸爆发释放的活性氧自由基能再次活化补体,在补体之间形成相互反馈活化、增强炎症应答的反应〔5〕。当前,该机制是否参与调控HSP的发生发展过程尚不清楚。本研究拟探讨补体活化和抑制情况下中性粒细胞介导的炎性反应的应答调控变化,以期为揭示HSP的致病机制提供科学依据。
1材料与方法
1.1试剂与仪器卵白蛋白-弗氏佐剂(均可购自于美国Sigma公司,制作方法:将卵白蛋白溶于0.9%的生理盐水,20 mg卵白蛋白/ml配制,并以1∶1的比例和弗氏佐剂混合);白细胞介素(IL)-2,-4,-6,-8、活性氧自由基(ROS)转化生长因子(TGF)-β和补体成分C3、C4、C5酶联免疫吸附法(ELISA)试剂盒购于上海恒远生物科技有限公司;N-乙酰半胱氨酸(NAC)、过氧化氢(H2O2),北京索莱宝生物科技有限公司;美国BIORAD酶标仪,型号Model 680;恒温水浴箱由河北黄骅仪器厂提供;721分光光度计,上海精密科学仪器有限公司,型号16C14型;其他试剂均为国产分析纯。
1.2实验动物的选择取24只健康清洁级日本大耳白兔〔购自于中山大学动物实验中心,合格证号:SCXK(粤)2004.001〕,兔龄4月,雌雄各半,体重2.2~3.0 kg。将白兔按照体重随机编号分为正常组和模型组各12只,雌雄各半。动物饲养于中山大学动物中心,温度18℃~25℃,相对湿度40%~60%,光暗照明时间各12 h,每笼1只,自由进食和饮水。实验动物操作经动物使用管理委员会批准。
1.3动物模型构建
1.3.1连续抗原刺激致敏将干姜、荜茇、胡椒水煎剂(购于广州中医药大学第一附属医院)用1∶1∶1的比例配置成终浓度为15 g/100 ml的液体溶液,在实验开始前2 w,正常组兔子每日给予200 g/只普通饲料;模型组兔子在比基础之上灌胃1 ml上述药液,每日1次,正常组用等量生理盐水灌胃,其他条件尽可能保持一致。另外,模型组动物还采取腹腔注射的方式加用卵白蛋白-弗氏佐剂每日1 ml/只,连续3 w,以达到连续抗原刺激致敏的目的,正常组动物注射等量生理盐水。
1.3.2抗原冲击激发过敏反应腹腔注射3 w后,模型组动物再通过耳缘静脉注射0.5 ml浓度为10 mg/ml的卵白蛋白生理盐水,背部皮肤剪毛,皮下分5个位置点注射0.3%卵白蛋白生理盐水0.2 ml。在抗原冲击激发过敏反应之后,观察每点皮肤局部红肿的情况以验证模型是否构建成功。
1.4一般指标观察实验过程中分别对两组兔子进行一般症状观察:平均每天饮食、饮水量、体温、血液常规和尿液常规;采取考马斯亮蓝法收集动物24 h尿量,并测定24 h尿蛋白。
1.5ROS、炎性因子和补体水平检测采用ELISA法检测血清ROS、IL-2、IL-4、IL-6、IL-8、TGF-β和补体C3、C4、C5水平;于96孔酶标板每孔加入稀释好的标准品,按ELISA试剂盒说明书操作。实验过程中取对照组和模型组兔各6只(雌雄各3只),分别采取腹腔注射加用ROS拮抗剂NAC和促进剂H2O2的方式,观察对两组兔子血清炎性因子及补体成分的影响。
2结果
2.1兔皮肤外部特征的变化皮下注射之后,兔皮肤注射位置均出现直径1 cm左右的皮丘。持续观察4~8 h之后,正常组兔背部的皮丘逐渐消失,而模型组兔背部的皮丘呈现增大的趋势,且出现皮肤周围组织水肿、皮下出血等症状。注射结束后24 h,模型组兔背部皮肤均呈现紫色斑块,形状不规则,大小不一,部分紫斑随时间延长出现结痂或坏死,而正常组兔的皮肤则无明显改变。
2.2兔体温、饮食、饮水量的变化正常组兔在致敏前、激发过敏后的体温、饮食量和饮水量均无明显改变(P>0.05);而模型组兔在激发过敏后,精神状态萎靡、运动量减少,饮水量和饮食量均较致敏前显著降低(P<0.05);与正常组相比,模型组兔在激发过敏后体温明显升高(P<0.05),而进食和饮水量则显著减少(P<0.05)。见表1。
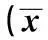
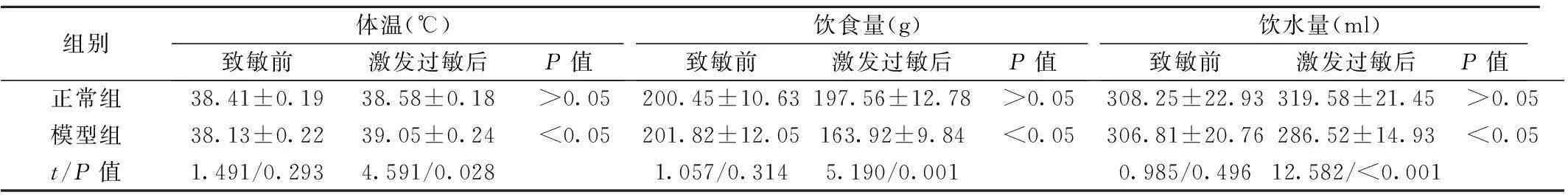
组别体温(℃)致敏前激发过敏后P值饮食量(g)致敏前激发过敏后P值饮水量(ml)致敏前激发过敏后P值正常组38.41±0.1938.58±0.18>0.05200.45±10.63197.56±12.78>0.05308.25±22.93319.58±21.45>0.05模型组38.13±0.2239.05±0.24<0.05201.82±12.05163.92±9.84<0.05306.81±20.76286.52±14.93<0.05t/P值1.491/0.2934.591/0.0281.057/0.3145.190/0.0010.985/0.49612.582/<0.001
2.3兔血液、尿液常规的变化致敏前模型组和正常组血液常规各项指标比较均无明显差异(P>0.05),且激发过敏后正常组兔的血液常规指标也无显著改变(P>0.05);但模型组兔激发过敏后,WBC、MCHC、NEU、NEU%水平明显升高,RBC、HGB、EOS和EOS%含量显著降低(P<0.05),而MCH和PLT则无明显变化(P>0.05)。另外,致敏前正常组和模型组兔的尿液常规检测指标均基本正常,激发过敏后,正常组兔尿液无明显变化,而模型组兔尿液出现拉丝、气味大的情况,其中66.67%出现尿蛋白阳性(8只)、58.33%出现尿红细胞(7只)、25%尿白细胞阳性(3只),而24 h排尿量、尿糖、尿胆红素、尿酮体、尿胆原、尿比重和酸碱度则均无明显变化,24 h尿蛋白从103.25 mg/24 h升高至402.45 mg/24 h(P<0.05)。见表2。
2.4兔ROS、炎性因子和补体水平变化致敏前正常组与模型组的ROS、IL-2、IL-4、IL-6、IL-8、TGF-β、补体C3、C4和C5的水平比较无明显差别(P>0.05);激发过敏后,正常组上述指标未发生显著改变,而模型组ROS、炎性因子、TGF-β、补体水平均明显升高;而且,模型组兔的这些指标水平也明显高于正常组(P<0.05)。ROS抑制剂NAC处理之后,ROS、炎性因子、TGF-β和补体C3、C4和C5的水平明显降低(P<0.05),而应用ROS激动剂H2O2处理之后,上述指标均明显升高(P<0.05)。见表3。
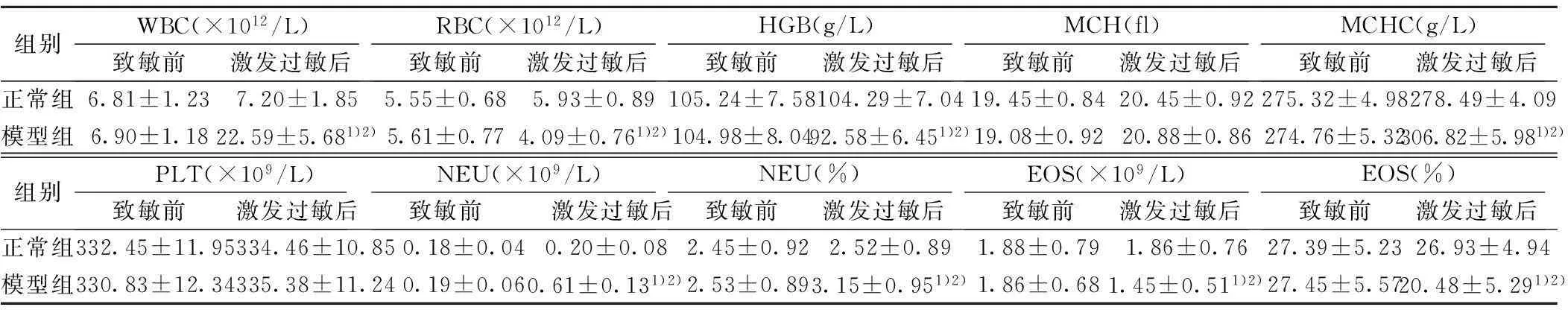
组别WBC(×1012/L)致敏前激发过敏后RBC(×1012/L)致敏前激发过敏后HGB(g/L)致敏前激发过敏后MCH(fl)致敏前激发过敏后MCHC(g/L)致敏前激发过敏后正常组6.81±1.237.20±1.855.55±0.685.93±0.89105.24±7.58104.29±7.0419.45±0.8420.45±0.92275.32±4.98278.49±4.09模型组6.90±1.1822.59±5.681)2)5.61±0.774.09±0.761)2)104.98±8.0492.58±6.451)2)19.08±0.9220.88±0.86274.76±5.32306.82±5.981)2)组别PLT(×109/L)致敏前 激发过敏后NEU(×109/L)致敏前 激发过敏后NEU(%)致敏前激发过敏后EOS(×109/L)致敏前激发过敏后EOS(%)致敏前激发过敏后正常组332.45±11.95334.46±10.850.18±0.040.20±0.082.45±0.922.52±0.891.88±0.791.86±0.7627.39±5.2326.93±4.94模型组330.83±12.34335.38±11.240.19±0.060.61±0.131)2)2.53±0.893.15±0.951)2)1.86±0.681.45±0.511)2)27.45±5.5720.48±5.291)2)
与致敏前比较:1)P<0.05;与正常组比较:2)P<0.05
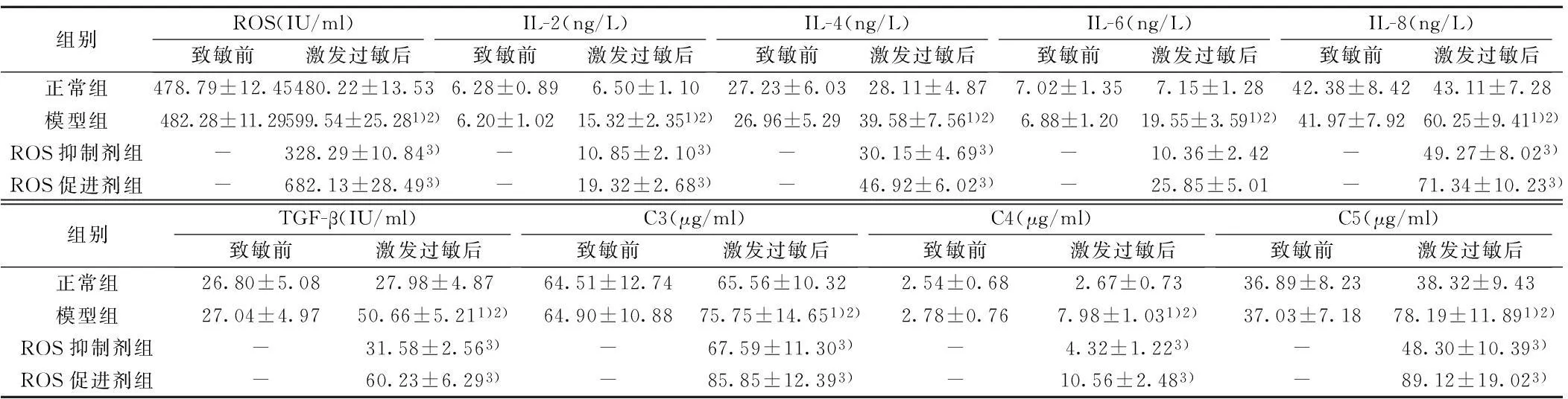
组别ROS(IU/ml)致敏前激发过敏后IL-2(ng/L)致敏前激发过敏后IL-4(ng/L)致敏前激发过敏后IL-6(ng/L)致敏前激发过敏后IL-8(ng/L)致敏前激发过敏后正常组478.79±12.45480.22±13.536.28±0.896.50±1.1027.23±6.0328.11±4.877.02±1.357.15±1.2842.38±8.4243.11±7.28模型组482.28±11.29599.54±25.281)2)6.20±1.0215.32±2.351)2)26.96±5.2939.58±7.561)2)6.88±1.2019.55±3.591)2)41.97±7.9260.25±9.411)2)ROS抑制剂组-328.29±10.843)-10.85±2.103)-30.15±4.693)-10.36±2.42-49.27±8.023)ROS促进剂组-682.13±28.493)-19.32±2.683)-46.92±6.023)-25.85±5.01-71.34±10.233)组别TGF-β(IU/ml)致敏前激发过敏后C3(μg/ml)致敏前激发过敏后C4(μg/ml)致敏前激发过敏后C5(μg/ml)致敏前激发过敏后正常组26.80±5.0827.98±4.8764.51±12.7465.56±10.322.54±0.682.67±0.7336.89±8.2338.32±9.43模型组27.04±4.9750.66±5.211)2)64.90±10.8875.75±14.651)2)2.78±0.767.98±1.031)2)37.03±7.1878.19±11.891)2)ROS抑制剂组-31.58±2.563)-67.59±11.303)-4.32±1.223)-48.30±10.393)ROS促进剂组-60.23±6.293)-85.85±12.393)-10.56±2.483)-89.12±19.023)
与致敏前比较:1)P<0.05;与正常组比较:2)P<0.05;与模型组比较:3)P<0.05
3讨论
本研究结果显示,在HSP模型兔中,炎性因子水平明显升高,而拮抗机体的ROS水平能够减弱中性粒细胞介导的炎症反应应答;并且,在炎症反应发生强度较大的情况下,模型兔的血液常规指标,如WBC、MCHC、NEU、NEU%等水平也出现明显异常改变以适应机体内的炎性应激反应。
目前的普遍观点认为炎症性疾病,尤其是在可形成免疫复合物的疾病中(如HSP等),组织的炎症损伤反应主要由补体系统的活化所介导〔6,7〕。有研究表明,活化的补体可产生过敏毒,进而对中性粒细胞产生趋化作用,并在细胞松弛素B作用下引起白细胞溶酶体酶释放〔8〕。动物实验研究亦证实,抑制补体的活化或清除ROS均可有效缓解动物皮肤动脉血管炎的症状。更值得关注的是,补体活化片段还可以趋化中性粒细胞,并激活其发生级联反应,释放ROS〔9〕。本研究结果也提示补体系统在HSP发生过程中被活化,更为重要的是,笔者发现抑制了ROS的水平能够减弱补体的活化程度,减轻炎症反应,表明补体-中性粒细胞反馈调节系统参与了HSP的形成和进展。
国外学者也曾报道,炎症损伤过程中中性粒细胞的激活及其产生的氧化代谢产物是活化补体的一种重要方式,并且两者之间存在相互促进的反馈作用,即炎性损伤过程中,补体活化诱发中性粒细胞呼吸爆发,而中性粒细胞产生的自由基又可促进补体的活化〔10,11〕。该反馈作用极可能是调节炎症反应及损伤的一个重要方式。此外,在器官和血管等炎症发生时,肿瘤坏死因子、IL、TGF-β等炎性因子表达增加,而中性粒细胞也可大量产生上述因子,诱发组织损伤,并且这些因子还能反过来激活机体产生更多的中性粒细胞,形成恶性循环〔12〕。在补体活化的前提下,炎性因子还可刺激骨髓产生并释放中性粒细胞,增加表面黏附分子的表达,加速中性粒细胞向组织侵入,诱导其发生呼吸爆发和脱颗粒,促使血管内皮细胞产生纤溶酶原激活物抑制因子,导致血管炎症发生〔13,14〕。本研究通过构建HSP动物模型发现补体-中性粒细胞反馈的炎症应答调控在HSP的形成中具有重要作用,而且还采用了NAC和H2O2双向调控ROS水平,从正反两方面证实了干预ROS能够抑制HSP的发生和进展。
4参考文献
1傅卓,任少敏,高晓峰,等.儿童过敏性紫癜早期肾损伤中VCAM-1的临床意义〔J〕.现代生物医学进展,2014;14(6):1095-6,1100.
2Tayabali S,Andersen K,Yoong W.Diagnosis and management of henoch-schonlein purpura in pregnancy:a review of the literature〔J〕.Arch Gynecol Obstet,2012;286(4):825-9.
3Kiryluk K,Moldoveanu Z,Sanders JT,etal.Aberrant glycosylation of IgA1 is inherited in both pediatric IgA nephropathy and Henoch-Schonlein purpura nephritis〔J〕.Kidney Intern,2011;80(1):79-87.
4Jen HY,Chuang YH,Lin SC,etal.Increased serum interleukin-17 and peripheral Th17 cells in children with acute Henoch-Schonlein purpura〔J〕.Pediat Aller Immunol,2011;22(8):862-8.
5Park SJ,Suh JS,Lee JH,etal.Advances in our understanding of the pathogenesis of Henoch-Schonlein purpura and the implications for improving its diagnosis〔J〕.Expert Rev Clin Immunol,2013;9(12):1223-38.
6Tahan F,Dursun I,Poyrazoglu H,etal.The role of chemokines in Henoch Schonlein purpura 〔J〕.Rheumatol Intern,2007;27(10):955-60.
7McDonald B,Pittman K,Menezes GB,etal.Intravascular danger signals guide neutrophils to sites of sterile inflammation〔J〕.Science,2010;330(6002):362-6.
8Zipfel PF,Skerka C.Complement regulators and inhibitory proteins〔J〕.Nature Rev Immunol,2009;9(10):729-40.
9Mantovani A,Cassatella MA,Costantini C,etal.Neutrophils in the activation and regulation of innate and adaptive immunity〔J〕.Nature Rev Immunol,2011;11(8):519-31.
10Chou RC,Kim ND,Sadik CD,etal.Lipid-cytokine-chemokine cascade drives neutrophil recruitment in a murine model of inflammatory arthritis〔J〕.Immunity,2010;33(2):266-78.
11Phillipson M,Kubes P.The neutrophil in vascular inflammation〔J〕.Nature Med,2011;17(11):1381-90.
12Fox S,Leitch AE,Duffin R,etal.Neutrophil apoptosis:relevance to the innate immune response and inflammatory disease〔J〕.J Inn Immun,2010;2(3):216-27.
13Wang R,Lu B,Gerard C,etal.Disruption of the complement anaphylatoxin receptor C5L2 exacerbates inflammation in allergic contact dermatitis〔J〕.J Immunol,2013;191(8):4001-9.
14Zhang J,Koh J,Lu J,etal.Local inflammation induces complement crosstalk which amplifies the antimicrobial response〔J〕.PLoS Path,2009;5(1):e1000282.
〔2014-02-17修回〕
(编辑徐杰)
