Lactobacillus casei Zhang传1 000-2 000代过程中的实验室进化特性*
2015-12-25马瑞芬惠文彦白梅张文羿孙天松
马瑞芬,惠文彦,白梅,张文羿,孙天松
(内蒙古农业大学乳品生物技术与工程教育部重点实验室,内蒙古呼和浩特,010018)
益生菌是指通过摄入适当量从而对宿主产生有益作用的活性微生物,其通常可来源于发酵食品、有益共生物和环境[1]。这些微生物和宿主间的相互作用机理多种多样,例如:影响共生菌群的结构组成和功能、改变宿主上皮细胞结构或者免疫应答[2]。近些年的研究成果显示,益生菌的摄入在特定条件下可改变特定人群的肠道菌群结构,包括过敏性皮炎、结肠炎和肠应激综合征,但一直缺乏有力的支撑证据[3-5]。
微生物的实验室进化研究(adaptive laboratory evolution,ALE)是在人为因素的作用下通过控制环境来研究微生物的进化。早在1983年,细菌学家Russell就列举了在进化生物学领域研究中细菌的一些“明显生长优势”,其中就提出严格控制细菌生长的变化的环境,并在适当时间和空间内为其更换新鲜培养基来研究细菌[6]。直到20世纪末实验室进化理论才以一个独立的学科问世[7]。最初的实验室进化是对恒化器中适应性进行准确测量以揭示自然突变与工程突变之间的微妙变化与联系[8]以及代谢与适应性间的关系[9]。之后的研究是长期连续传代实验。伴随着分子技术的进步,实验室进化已经延伸到基因水平的研究。通过全基因组序列的测定,可进一步揭示进化的动力学问题[10]。
Lactobacillus casei Zhang(L.casei Zhang)是从内蒙古地区蒙古族家庭制作的16份酸马奶样品中分离得到的1株益生乳杆菌[11]。具有以下特点:优良的益生特性,包括酸和胆盐耐受性[12]、降胆固醇、抗氧化、抗菌、免疫调节以及调节肠道菌群结构等[13-16]。本文在已有工作基础之上,检测了L.casei Zhang在普通MRS培养基中连续传代1 000~2 000代期间3个谱系的进化特性变化情况。
1 材料与方法
1.1 材料
1.1.1 菌株来源
益生菌L.casei Zhang培养1 000代后的衍生菌株是由内蒙古农业大学“乳品生物技术与工程”教育部重点实验室乳酸菌菌种库(LABCC)提供,LABCC编号IMAU10048。
1.1.2 试剂与仪器
试剂:法国梅里埃合成无糖MRS培养基,纽西兰超特级脱脂乳粉,葡萄糖、HCl、NaOH、NaCl、琼脂粉等常用试剂均为国产分析纯试剂。
仪器:洁净工作台(SW-CJ-1F型),苏净集团苏州安泰空气技术有限公司;恒温培养箱(LRH-500F型),上海一恒科学仪器有限公司;台式高速冷冻离心机(Eppendorf,5810R 型),德国;pH 计(SJ-3F 型),上海精密科学仪器有限公司雷磁仪器厂;超纯水系统(MUL-9000,昆山总馨企业有限公司;紫外可见分光光度计(UV-1700 PharmaSpec型,SHIMADZU);全自动高压灭菌锅(MLS-3751L型,PANASONIC)等。
1.2 实验方法
1.2.1 L.casei Zhang连续传代培养及培养代数计算
将1 000代益生菌L.casei Zhang三株平行菌株L.casei Zhang-1、L.casei Zhang-2、L.casei Zhang-3 按体积分数1%接种量进行长期连续传代培养至2 000代。在每个传代周期(24 h)内,菌体菌数约增长100倍,生长代数约为6.64 代[17]。
1.2.2 L.casei Zhang细胞形态观察
在菌株L.casei Zhang连续传代过程中,培养至2000代时对菌株进行革兰氏染色、油镜观察、照相,观察培养周期末菌株细胞形态。
1.2.3 L.casei Zhang培养周期末活菌数变化
在菌株传代过程中,每培养200代测定各个菌株在培养周期末(24 h)的活菌数。样品经倍比稀释到合适梯度后,采用MRS固体培养基平皿倾注法,37℃厌氧培养72 h后计菌落总数,选取菌落数在30~300之间的平板作为活菌总数测定的指标。
1.2.4 L.casei Zhang培养周期末浊度变化
在菌株传代过程中,每培养200代测定连续3天的培养周期末的培养液浊度。传代培养液用未接菌培养基适当稀释,采用比浊法于600 nm处测定其浊度。
1.2.5 L.casei Zhang产酸活力测定
在菌株传代过程中,每培养200代测定菌株的产酸活力。各菌株以1×107/mL接种于10 mL灭菌脱脂乳中,37℃恒温培养24 h,取出后立即放入冰浴,每样加30 mL蒸馏水稀释后,以酚酞作指示剂,以0.1 mol/L的NaOH滴定至微红色30 s不退色为止,记录消耗NaOH的体积以计算出样品中的乳酸浓度,再换算为活力单位。
发酵活力:以每107个菌体细胞在此条件下发酵1 mL脱脂乳产生1 μg乳酸计为1个活力单位(U)[18]。
U=0.1 V×M
式中:U为活力单位;V为消耗NaOH的体积;M为乳酸的摩尔质量。
1.2.6 数据处理
采用DPS软件对数据进行方差分析;采用Origin8.6绘图软件进行绘图。各项指标均做3个平行。
2 结果与分析
2.1 L.casei Zhang细胞形态观察
图1所示为L.casei Zhang分别在1 000代和2 000代时显微镜照相的照片。由图1可知,L.casei Zhang 1 000代的3个谱系的细胞均为短杆状、成对或呈链杆状排列。与1 000代3个谱系细胞形态相比,在MRS培养基中连续传代至2 000代,3个谱系的细胞形态与之前相同,细胞体积也没有发生显著变化。这说明,L.casei Zhang细胞未发生明显的变异情况。当然,这一结论只能作为评价该谱系群体进化规律的部分参考,关于其细胞内部成分组成及其比例是否发生变化仍然需要更进一步的研究。

图1 L.casei Zhang传1 000~2 000代期间细胞形态对比Fig.1 Cell appearance of Lactobacillus casei Zhang during subculturing for 1 000~2 000 generations
2.2 L.casei Zhang生长周期末活菌数
L.casei Zhang在连续培养1 000~2 000代期间,不同代数3个谱系所形成的群体在培养周期末的活菌数变化情况见图2。
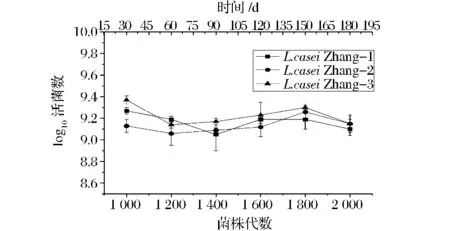
图2 L.casei Zhang在不同培养代数的活菌数变化Fig.2 Viable counts of L.casei Zhang of different generations
由图2可以看出,在MRS培养基中,3个谱系活菌数在传代过程中变化不明显;纵向比较3个平行培养的菌落发现,在最初开始培养时活菌数就有差异,但这样的差异并没有随培养代数的增加而持续。总体来看,L.casei Zhang在传代1 000~2 000代期间,3个谱系间活菌数的进化趋势基本平行,菌株的平均适应性也相对稳定。侧面反映了该菌株具有稳定的表型特性。
2.3 L.casei Zhang周期末浊度
在连续培养1 000~2 000代过程中,L.casei Zhang在培养周期末的OD600值变化情况见图3。由图3可以看出L.casei Zhang三个谱系的OD600值在1 400代之前均呈现增长的趋势,到1 600代时达到最低点又整体下降,之后又有所增加。通过对比活菌数发现菌株在1 600代时适应性与之前相比无明显差异,因此推断3个谱系在连续进化1 000~2 000代期间OD600值产生的不规律的变化可能是最初接菌时产生的误差所致。3个谱系之间周期末浊度没有发生明显变异.且处于平行进化的趋势。
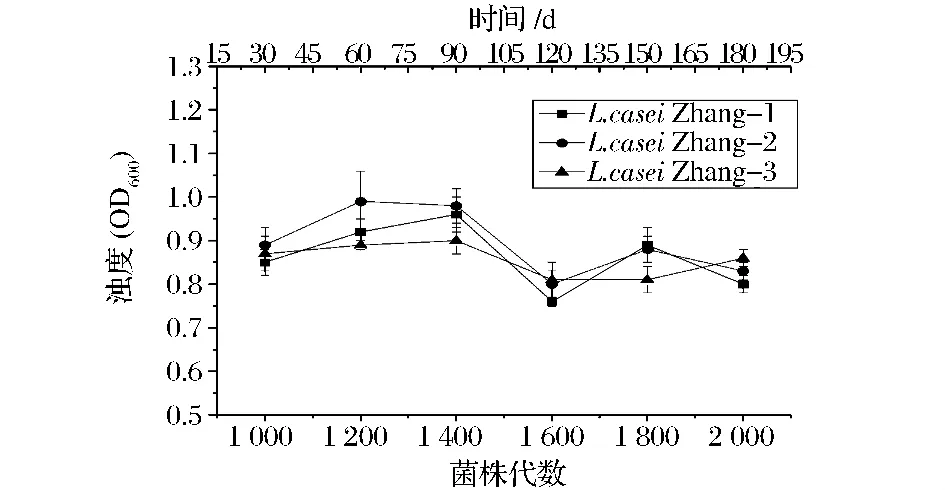
图3 L.casei Zhang在不同培养代数的浊度变化Fig.3 The OD600value of L.casei Zhang of different generations
2.4 L.casei Zhang产酸活力变化
实验研究了连续传代培养1 000~2 000代过程中,每200代测定菌株的产酸活力,测定结果如表1所示。由表1可以看出,菌株在1 400代时3个谱系的产酸活力均有微小下降,但变化不显著。

表1 L.casei Zhang在不同培养代数的菌体产酸活力变化(U)Table 1 Acid-producing ability of L.casei Zhang of different generations
3 讨论
近年来,关于微生物实验室进化特性的研究越来越多,早期有对单细胞藻类的研究[21],还有利用分子生物学技术对酵母进化特性进行研究[22]。一般来说,在微生物进化过程中,必然的会发生菌株变异,从而导致菌株部分特性的改变,这也是进化的必然趋势。例如,Merry等人对土壤细菌实验室进化1 000代进化特性的观察发现群体细胞长度有增加的现象[23]。此外,对大肠杆菌连续传代40 000~50 000代进化特性研究发现中,其平均适应性呈现递增趋势,其中增加最多达3%[24]。除了菌株适应性增长之外,还有一些显著的表性特征的改变,如细胞的体积随着传代的进行呈现增加趋势。或者是菌落形态比原始菌株形状更圆等[25]。这些表观特性的变化可能是细菌为了更好地适应环境而做出的“自我改变”。
与其他人研究结果不同的是,在对益生菌L.casei Zhang在普通MRS培养基中传代1 000~2 000代期间实验室进化特性的研究发现,该菌株的表观特性包括细胞形态、活菌数、浊度和菌株活力等指标均未发生显著变异,而且保持着几乎平行的进化趋势。由此可以侧面反映出L.casei Zhang是一株相对稳定的益生菌菌株。但是,由于实验所测指标仍然存在一定局限性,该结果并不能够全面评价该菌株的进化特性。
4 结论
研究结果表明,益生菌L.casei Zhang在正常培养条件下3个谱系间表性特征进化趋势相对平行,然而,这些进化特性的结果仍存在局限性。一方面,细菌由于世代时间较短,繁殖迅速,即使出现某些突变,也可能会在短期内消失,这就给研究带来一定困难,另一方面,本实验传代时间较短,而菌株的一些特性需要突变不断积累才会表现出来,这也需要更充足的数据来验证。总体来说,这些数据虽不能全面说明L.casei Zhang在生理学特性方面是否发生了微妙的变化,但该实验结果仍然可以作为评价该菌株进化特性的有力依据,也可为该菌株的产业化生产提供很好的保障。
[1] Sanders M E,Guarner F,Guerrant R,et al.An update on the use and investigation of probiotics in health and disease[J].Gut,2013,62(5):787-796.
[2] Bengmark S.Gut microbiota,immune development and function[J].Pharmacological Research,2013,69(1):87-113.
[3] Arora T,Singh S,Sharma R K.Probiotics:Interaction with gut microbiome and antiobesity potential[J].Nutrition,2013,29(4):591-596.
[4] Barraud D,Bollaert P E,Gibot S.Impact of the administration of probiotics on mortality in critically ill adult patients probiotics in the ICUA Meta-analysis of randomized controlled trials[J].CHEST Journal,2013,143(3):646-655.
[5] De Greef E,Vandenplas Y,Hauser B,et al.Probiotics and IBD[J].Acta Gastro-enterologica Belgica,2013,76(1):15-19.
[6] Russell H L.Bacteriology in its general relations[J].American Naturalist,1893,27(322):847-859.
[7] Elena S F,Lenski R E.Evolution experiments with microorganisms:the dynamics and genetic bases of adaptation[J].Nature Reviews Genetics,2003,4(6):457-469.
[8] Dykhuizen D E,Hartl D L.Selection in chemostats[J].Microbiological Reviews,1983,47(2):150.
[9] Dykhuizen D E,Dean A M,Hartl D L.Metabolic flux and fitness[J].Genetics,1987,115(1):25-31.
[10] Hegreness M,Kishony R.Analysis of genetic systems using experimental evolution and whole-genome sequencing[J].Genome Biol,2007,8(1):201.
[11] 孟和毕力格,乌日娜,王立平,等.不同地区酸马奶中乳杆菌的分离及其生物学特性的研究[J].中国乳品工业,2004,32(11):6.
[12] 张和平,孟和毕力格,王俊国,等.分离自内蒙古传统发酵酸马奶中L.casei Zhang潜在益生特性的研究[J].中国乳品工业,2006,34(4):4-10
[13] WU R,WANG W,YU D,et al.Proteomics analysis of Lactobacillus casei Zhang,a new probiotic bacterium isolated from traditional home-made koumiss in Inner Mongolia of China[J].Molecular& Cellular Proteomics,2009,8(10):2 321-2 338.
[14] WANG J C,ZHANG W Y,ZHONG Z,et al.Transcriptome analysis of probiotic Lactobacillus casei Zhang during fermentation in soymilk[J].Journal of Industrial Microbiology& Biotechnology,2012,39(1):191-206.
[15] WANG J,WU R,ZHANG W,et al.Proteomic comparison of the probiotic bacterium Lactobacillus casei Zhang cultivated in milk and soy milk[J].Journal of Dairy Science,2013,96(9):5 603-5 624.
[16] Lenski R E,Rose M R,Simpson S C,et al.Long-term experimental evolution in Escherichia coli.I.adaptation and divergence during 2,000 generations[J].American Naturalist,1991,138(6):1 315-1 341.
[17] WANG J,GUO Z,ZHANG Q,et al.Fermentation characteristics and transit tolerance of probiotic Lactobacillus casei Zhang in soymilk and bovine milk during storage[J].Journal of Dairy Science,2009,92(6):2 468-2 476.
[18] 李妍.益生菌Lactobacillus casei Zhang高密度培养技术及发酵过程中关键酶基因表达变化的研究[D].呼和浩特:内蒙古农业大学,2008.
[19] Colegrave N,Kaltz O,Bell G.The ecology and genetics of fitness in Chlamydomonas.VIII.The dynamics of adaptation to novel environments after a single episode of sex[J].Evolution,2002,56(1):14-21.
[20] Zeyl C.Experimental evolution with yeast[J].FEMS Yeast Research,2006,6(5):685-691.
[21] Riley M S,Cooper V S,Lenski R E,et al.Rapid phenotypic change and diversification of a soil bacterium during 1000 generations of experimental evolution[J].Microbiology,2001,147(4):995-1 006.
[22] Wiser M J,Ribeck N,Lenski R E.Long-term dynamics of adaptation in asexual populations[J].Science,2013,342(6164):1 364-1 367.
[23] Lenski R E,Mongold J A.Cell size,shape,and fitness in evolving populations of bacteria[C].Scaling in Biology.Oxford University Press,2000:221-235.
