日粮中添加甲基供体对妊娠母猪妊娠状况和产仔数的影响
2015-12-25vanWettereSmitsHughes
W.H.E.J.van Wettere, R.J.Smits, P.E.Hughes
(1.澳大利亚阿德莱德大学动物和兽医科学,罗斯沃斯校区,SA 5371;2.澳大利亚克洛瓦Rivalea有限公司,新南威尔士州2646;3.澳大利亚阿德莱德大学猪和家禽生产研究所,罗斯沃斯校区,SA 5371)
母猪的繁殖性能主要表现为窝产仔数和年产胎次数,其主要受母体系统对胚胎发育状况识别偏差而导致的流产、死胎(妊娠50 d前)等因素影响(Foxcroft等,2007)。虽然排卵数随着胎龄增加而增加,但是由于4胎龄以上的母猪胚胎的死亡率也有所增加,高胎龄母猪的胚胎损失是限制其窝产仔数的主要因素(Town等,2005)。
妊娠母猪在妊娠前期30~60 d时对叶酸、维生素B12等B族维生素的需要量显著增加(Matte和 Girard,1999)。然而,目前 B 族维生素的推荐用量是20~50年前给出的基于预防基因缺陷的最低剂量 (Stahly 等,2007;Matte 等,2006)。该推荐剂量建议,现今母猪达到最大繁殖性能所需要的B族维生素理想摄入量可能过高(Matte等,2006),同样观点还有,日粮叶酸的添加仅仅是为了在排卵率较高时可以减少胚胎的死亡率(Tremblay 等,1989),同时,在经产母猪和初产母猪的妊娠期叶酸添加量相同时,其窝产仔数也无差异(Matte 等,2006)。
叶酸同维生素B12、甘氨酸共同参与同型半胱氨酸 (HCY)转化蛋氨酸过程中的再次甲基化反应,从而促进胚胎的正常发育和细胞增殖(Kwong等,2010)。HCY被证明是一类胚胎致畸因子(van Mil等,2010;Matte 等,2006), 血浆中 HCY 含量上升,能导致多个物种胚胎发育畸形和胚胎死亡率增加(Di Simone 等,2004;Guay 等,2002a)。 与正常妊娠母猪比较,经历早期妊娠失败的母猪血浆中HCY含量较高。维生素B12作为HCY再次甲基化反应的酶辅助因子,若维生素B12缺乏时,可以通过补充叶酸来改善母猪的繁殖性能(Matte等,2006)。前期试验表明,在夏季配种的高胎龄妊娠母猪日粮中添加甜菜碱,能够增加母猪产仔数(van Wettere和 Herde,2009)。 在妊娠母猪日粮中提高叶酸原体、维生素B12和甜菜碱的含量对产仔数的影响需进一步探索。
因此,本试验主要有两个目的:(1)考虑在经产妊娠母猪日粮中单独添加甜菜碱或同时添加维生素B12和叶酸是否可以降低妊娠失败率和提高母猪产仔数。(2)研究B族维生素和甜菜碱对血浆中HCY含量稳定性的影响。
1 材料与方法
1.1 试验动物和饲养管理 试验在澳大利亚南威尔士州一商业猪场 (Rivalea Australia Pty Ltd)进行。试验共选取平均胎龄4.4±0.1(范围2~9胎龄)的长×大二元经产母猪1079头,试验开始时间共分5周,每周约有220头。试验母猪的平均发情间隔为(5.2±0.1)d,在母猪断奶后第一次发情时和随后的24 h分别进行人工受精 (2010-3-27至2010-5-5),在试验配种前,试验母猪的平均窝产仔数为(9.4±2.2)头。
试验采用2×2×2析因法试验设计,试验因素水平分别是两个添加量水平的甜菜碱(0、3g/kg)、B 族维生素(0、20 mg/kg 叶酸+150 μg/kg 维生素B12)以及两个胎龄母猪(2~3胎龄、4胎龄及以上),甜菜碱和叶酸、维生素B12的添加含量分别参考 van Wettere和 Herde(2009)和 Guay等(2002a)推荐剂量。基础日粮中叶酸和维生素B12的含量分别为5 mg/kg和20 μg/kg,分别高于NRC(1998)的推荐剂量 0.3 mg/kg 和 10 μg/kg。试验日粮从配种后第一天开始饲喂到妊娠112天止,表1为日粮组成和营养水平。所有的试验母猪在妊娠第1~42、43~84、85~112天的日饲喂量分别为2.5、2.7、3.0 kg。试验母猪在妊娠第一天起至妊娠第112天进入产房前均单独饲喂在妊娠限位栏中,每排限位栏有55个栏位,各栏位采用一排相同的食槽和饮水槽。每天7:00时用标准量斗(量程为200 g)喂料一次,同一处理组为一排,避免不同处理组日粮间相互干扰。在配种后第18~24天,用查情公猪在母猪栏外与母猪接触来鉴别母猪发情,若母猪这时表现发情被认为是正常返情。在配种第30天后,采用5-MHz腹式超声波仪对母猪进行妊娠检查 (A-gri-Scan A-7,ECM)。为确保诊断准确,对诊断为妊娠失败的母猪,在一周后进行复查。返情和经超声波检查为妊娠失败的母猪需转出限位栏,并进行登记。若试验母猪突然死亡或者母猪体况较差,也同样进行登记。试验母猪经超声波检查为妊娠成功,但在产房并未生产的,被视为后期流产。
在试验母猪妊娠第112天时转入产房,所有试验母猪饲喂相同的哺乳期日粮 (消化能14.9 MJ/kg、总赖氨酸 9 g/kg、粗蛋白质 185 g/kg),每天7:00喂食一次,共3 kg,直至2 d后分娩。
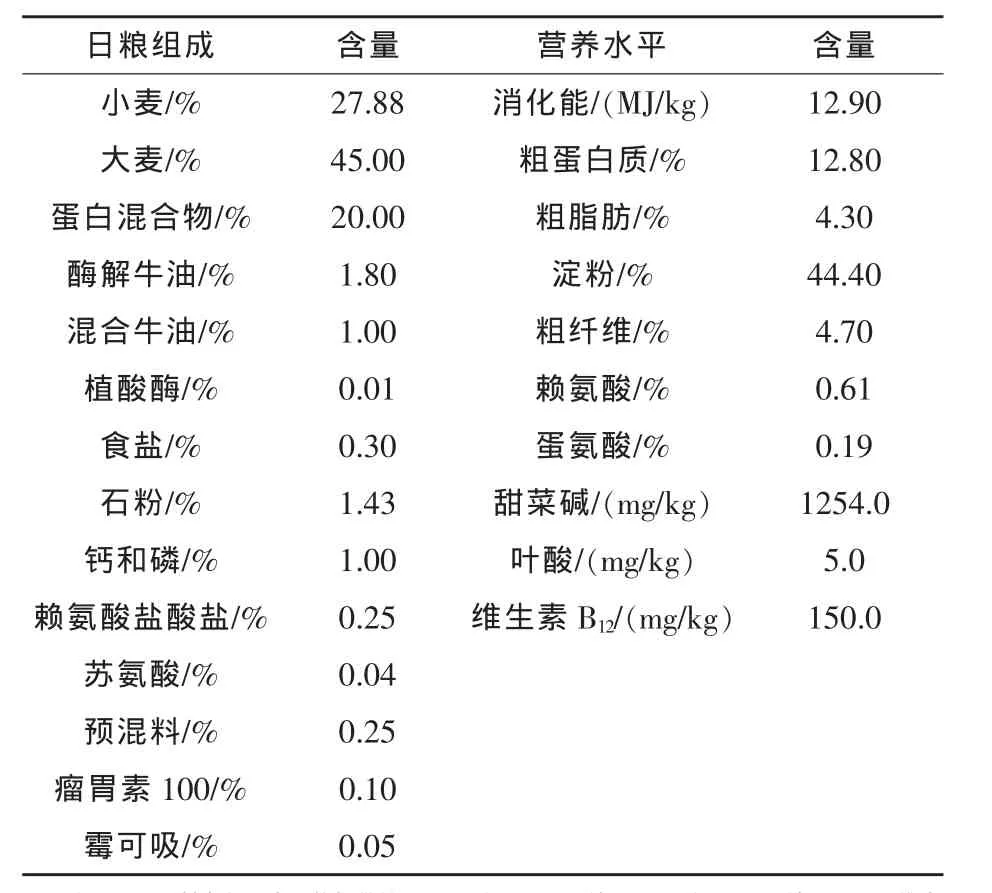
表1 妊娠期基础日粮组成及营养水平
1.2 生产性能测定 母猪配种后前3周返情数(返情)、早期(小于配种后30天)或后期(大于配种后30天)妊娠失败数、后期流产数等均视为非繁殖因素影响母猪而排除。母猪分娩时,记录总产仔数、产活仔数、产死仔数、产木乃伊数。每个处理组随机选取50窝仔猪,在仔猪出生后24 h内,对活猪仔进行逐只称重并作为窝重统计。
1.3 血样采集、激素含量和血液代谢成分分析每个处理组选取20头试验母猪,分别在妊娠期第3、30、107天早上空腹采集血样分析 HCY含量(人工受精后24 h为妊娠0天计)。采用前腔颈静脉采血法采集血样,血样放在9 mL氯化锂覆膜离心管中,将血样放冰块上静置约1 h,用常温离心机2000 g离心15 min,收集上层血浆于2 mL离心管中,-20℃环境下保存,供后续分析。

表2 不同胎龄母猪妊娠日粮中添加叶酸+维生素B12、甜菜碱对总产仔数、产活仔数、产死胎数和木乃伊数的影响
吸取25 μL血浆样品,选用BioRad试剂盒,按照操作说明,采用酶联免疫法测定血浆中HCY含量。试验原理如下:蛋白质结合的HCY被分解为游离HCY,后者在酶联免疫检测前,被分解转化为s-腺苷-L-高半胱氨酸。组间检测的变异系数远低于5%,组内的变异系数在10.8 μmol/L和20.6 μmol/L时分别为8.2%和6.4%。
1.4 数据统计分析 试验数据以 “平均值±标准差”表示,采用多因素方差分析对试验区组数据进行显著性检验,研究日粮组成和母猪胎龄对其生产性能的影响,前胎仔猪断奶头数和母猪发情间隔作为协变量。处理组间差异用LSD法检验,对各因素的主效应进行分析,当因素间存在交互效应时,也进行了相应标注。血浆中HCY含量采用线性混合模型进行分析,每头母猪作为一个独立试验单位,固定模型包括区组、配种胎龄、血浆样本采集时间等。应用卡方检验法分析处理因素对母猪妊娠率的影响。除卡方检验外,所有的数据统计均采用GENSTAT软件进行分析(第8版,英国洛桑试验站统计委员会)。
2 结果
2.1 生产性能指标 表2表明,未添加叶酸与维生素B12处理的母猪配种胎龄和日粮添加甜菜碱这两因素在窝产仔数指标上有极显著的交互作用,4胎龄以上的日粮中添加甜菜碱较4胎龄以上对照组母猪(未加甜菜碱)在总产仔数和产活仔数上都显著提高,分别为 11.7±0.18、11.1±0.19 和10.8±0.17、10±0.18。相比较,同样在妊娠日粮中添加叶酸和维生素B12,2~3胎龄母猪较4胎龄以上母猪产更多仔猪,分别为12.3±0.30和11.0±0.34。未加B族维生素及甜菜碱的不同胎龄的母猪(2~3胎龄和4胎龄以上)在窝初生仔猪总数和活仔数等指标上差异显著。
表3结果显示,妊娠日粮中添加叶酸和维生素B12等B族维生素的母猪较未添加组的母猪能够降低早期流产率,但妊娠日粮中添加甜菜碱与否对母猪早期流产率没有影响,B族维生素和甜菜碱对母猪返情、后期流产、非繁殖因素未产等发生率无影响。

表3 妊娠日粮添加甜菜碱和叶酸+维生素B12对母猪返情、早期流产和后期流产、非繁殖因素未产数的影响
妊娠日粮中添加甜菜碱与否对仔猪初生重[(1.48±0.02)kg 和(1.51±0.02)kg]及初生重变异系数[(0.20 ±0.01)和(0.20 ±0.01)]均无影响,同样,B族维生素添加与否对仔猪初生重[(1.49±0.02 kg)和(1.51±0.02)kg]及初生重变异系数(0.20 ±0.01)和(0.20±0.01)也无影响。然而,高胎龄母猪(4胎龄以上)的仔猪初生重较低胎龄母猪(2~3胎龄)的轻 80 g,分别为(1.48±0.02)kg 和(1.56±0.03)kg,差异显著(P<0.05),与初生重相似,高胎龄母猪的仔猪窝初生重变异系数显著高于低胎龄母猪,分别为 0.21±0.01 和 0.18±0.01。
2.2 血浆中同型半胱氨酸 图1是妊娠日粮中添加甜菜碱和B族维生素对母猪血浆中HCY含量的影响。总体看来,添加B族维生素能够显著降低血浆HCY含量(P<0.001),而添加甜菜碱无显著影响。妊娠天数对血浆HCY含量影响明显,妊娠第 3 天[(14.6±0.27)μmol/L]、第107 天[(14.0±0.27)μmol/L]显著高于妊娠第 30 天[(12.8±0.28)μmol/L]。妊娠日粮组成与妊娠天数在HCY含量上有明显的交互作用。添加B族维生素组母猪在妊娠第3天和第107天的血浆HCY含量较未添加组分别降低2.2 μmol/L和2.8 μmol/L(P <0.001),但对妊娠第30天的HCY影响不大 (图2)。同样,添加甜菜碱降低妊娠第3天血浆HCY含量1.2 μmol/L,但对第30天和第107天的血浆HCY含量无影响(图3)。
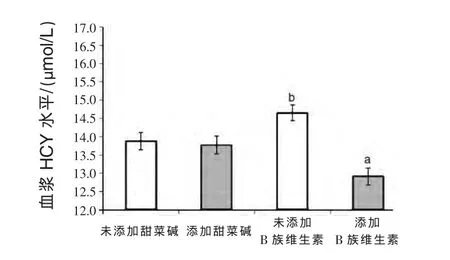
图1 妊娠日粮组成对血浆中HCY含量的影响

图2 B族维生素添加量对不同妊娠阶段母猪血浆中HCY含量的影响

图3 甜菜碱添加量对不同妊娠阶段母猪血浆中HCY含量的影响
3 讨论
母猪的繁殖能力由初生仔猪数量来衡量。在母猪妊娠期日粮中添加B族维生素(叶酸和维生素B12)和甜菜碱(三甲基甘氨酸)能够改善母猪繁殖性能。本试验研究证实在妊娠母猪日粮中同时添加叶酸和维生素B12,可以增加窝产仔数和降低早期妊娠失败率。与前期研究结果一致(van Wettere和 Herde,2009;Matte 等,2006)。母猪配种时的胎龄可以影响添加B族维生素和甜菜碱作用,特别是叶酸和维生素B12能够增加低胎龄母猪的窝产仔数,而甜菜碱能够增加高胎龄母猪的窝产仔数。添加B族维生素和甜菜碱能够改变妊娠期母猪血浆HCY原体的代谢过程。妊娠日粮添加B族维生素,母猪在妊娠第30天,其血浆HCY含量降低了8%,在妊娠第3天和第107天,则分别降低了18%和17%。
妊娠期的特征之一是机体代谢对B族维生素的需求急剧增加,同样需要B族维生素的还有血液系统 HCY 含量的改变(Holmes,2003;Matte等,2006)。在妊娠前期55~60 d,血浆中叶酸含量快速下降33% ~42%(Harpe等,1994)。子宫对叶酸和维生素B12的大量吸收以及转运至子宫腔,以在妊娠前15~25 d促进胚胎发育(Guay等,2002a;Vallet等,1999)。HCY 含量的动态平衡与B族维生素原体的含量水平密切相关(Matte等,2006),在孕妇体内HCY含量与甜菜碱含量呈反比(Velzing-Aarts等,2005)。 重要的是,血液中HCY的含量过高则表明机体甲基化途径失衡(Ikeda等,2010),将引起多种生物的妊娠异常和胚胎定植失败(Ikeda 等,2010;Matte等,2006)。因此,血液系统的HCY水平可以作为试验处理效应指标,来确定胚胎定植前(妊娠第3天)和定植后(妊娠第30天)、妊娠后期(妊娠第107天)母体和胎儿蛋氨酸循环情况。与相关报道一致(Simard等,2007;Guay 等,2002a),本试验添加叶酸和维生素B12,妊娠期第3天血浆中的HCY含量降低了14%。叶酸和甜菜碱为HCY再甲基化提供碳单位(Solanky 等,2010;Matte 等,2006),同时本试验研究表明甜菜碱能降低妊娠早期机体的HCY含量,甲基的有效利用率可以制约妊娠早期蛋氨酸再甲基化。
本研究表明,未添加B组维生素或甜菜碱的母猪在妊娠期前30 d,其血浆HCY含量逐渐下降,而在妊娠107 d时恢复至围产期水平。近期研究也表明,相同的变化趋势也发生在日粮添加有叶酸和维生素 B12的母猪上(Simard 等,2007),同样日粮添加有甜菜碱的母猪也有同样变化趋势。因此,这似乎说明,HCY的变化是一正常的生理现象,与B族维生素的缺乏和甲基供体的利用率无关。与妊娠相关的血液稀释性和血浆白蛋白的变化都不能够解释母猪体内HCY含量下降现象(Simard 等,2007)。 因此,Holmes (2003) 提出了HCY的胎盘转移学说,统一于胎儿的代谢循环系统而被逐渐利用,这也许是在循环过程中HCY含量降低的原因。
添加B族维生素,母猪在配种后前30 d的流产比例降低了57%,同时对于2~3胎龄的母猪,其窝产子数提高了5%。妊娠日粮中添加叶酸对窝产仔数的影响还有争议,在叶酸的添加量相近时,窝产仔数增加了 10%(Lindemann和Ko rnegay,1989)和窝产仔数无变化等均有相关报道(Harper等,1994;Thaler等,1989)。 添加叶酸能够增加母猪窝产仔数可能是叶酸能够减少妊娠前30天的胚胎死亡率(Tremblay等,1989)。胚胎死亡数量的减少与胚胎定植时和定植后的前列腺素E2含量变化相关 (Matte等,2006;Guay等,2004),胚胎定植前的发育率增加也是如此(Guay等,2002b ;Duquette等,1997)。 目前研究揭示,母体对妊娠的识别过程引起的前列腺素E2含量变化,可以对减少母猪早期流产进行解释。Lindemann 和 Kornegay(1989)也指出,母猪日粮中添加叶酸降低了产仔数。同样,妊娠失败能够引起妊娠前期母猪体内HCY含量上升,推测由B族维生素引起的HCY含量降低或许是因为母猪流产数的减少。
添加叶酸能够提高窝产子数表明基础日粮叶酸不能够满足母猪的最佳繁殖性能,对于2胎龄和2胎龄以上母猪及待配的后备母猪,其窝产子数的增加仅源于较高的排卵率 (Town等,2005)。多数流产发生在胚胎定植前和定植后,主要是因为个体间的形态和发育的多样性而导致的次成熟胚胎未能存活(van Wettere等,2005)。胚胎发育不同步性的自然现象揭示卵泡发育和排卵时卵母细胞成熟度的差异,这种情况在排卵率较高时更为明显(vanWettere 等,2005;Xie 等,1990)。 一般认为添加叶酸能够促进次成熟胚胎生长和发育成熟,使得其成功定植。日粮中添加相近水平的叶酸(15 mg/kg)和维生素 B12(160 μg/kg),子宫液中HCY的含量降低30%(Guay等,2002a)。叶酸是细胞分裂和增殖过程中合成核酸的重要辅助因子,胚胎的快速发育对叶酸的需求量巨大(Solanky等,2010)。在子宫内,叶酸和 HCY 含量的反比关系是显而易见的(Solanky等,2010)。体外试验表明,叶酸能够促进猪胚囊的生长发育(Kim等,2009)。子宫腔内较高的B族维生素含量,可以为胚胎和胎盘的代谢和发育所利用,同时HCY含量也下降,这大概是母猪妊娠日粮补加叶酸和维生素B12改善其繁殖性能的原因。
与本研究结果一致,前期研究发现叶酸对繁殖力的作用受胎龄影响(Matte等,2006)。近期的报道表明,B族维生素并不能让2~3胎龄母猪获得理想的胚胎发育和胚胎存活率。Town等(2005)研究认为,排卵率在2~3胎龄和4胎龄及以上的母猪之间并无明显差异。因此,排卵率与胚胎发育的差异并不能归咎于叶酸对提高高胎龄母猪窝产仔数无效。暂不论试验处理和相关文献的差异,Mahan(1998)报道窝产仔数在第3胎龄达到最高,之后一直稳定至第6胎龄。曾有报道认为,第4胎龄以后窝产仔数是下降的,导致窝产仔数下降的原因目前还不清楚,可能是因为子宫功能受损或者与年龄相关的卵母细胞发育能力的衰弱(Armstrong,2001)。
然而,B族维生素对增加高胎龄母猪的窝产仔数作用不明显,表明B族维生素被有效利用并不是限制高胎龄母猪潜在的繁殖能力的因素。相反,对于低胎龄母猪,B族维生素能够提高窝产仔数,表明其体内B族维生素不足,尤其是维生素B12。叶酸对快速生长的初产母猪的窝产仔数增加作用不明显,因为叶酸主要是作为HCY再次甲基化反应酶促因子和参与母体生长对维生素B12的竞争性吸收(Matte等,2006)。前期试验表明,母猪的体重和背膘厚是持续增加的,直至第4胎龄达到成熟的体重和体组成,因此对于低胎龄母猪,其仍然处于生长阶段,相较于年龄较大、生理更成熟的母猪,其对维生素B12的需求将持续增加。
相比B族维生素,妊娠日粮中添加甜菜碱能够增加高胎龄母猪的窝产仔数,这与前人试验结果一致(van Wettere和 Herde,2009)。 甜菜碱的主要功能是作为甲基供体参与HCY再次甲基反应,至少在妊娠前期,其作用体现在能够降低妊娠第3天血浆中HCY含量,然而,甜菜碱的正效应为什么仅限于高胎龄母猪,目前的研究尚不完全明确。在生长猪日粮中添加甜菜碱,其生长激素(GH)和生长激素脉冲振幅分别提高了42%和35%(Huang等,2007)。 在妊娠前期,母体生长激素能够增加胎盘养分转运至胚胎,促进妊娠28 d时较小胚胎的生长和提高妊娠50 d胎儿总重,同时也可以提高胚胎的成活率 (Gatford等,2009;Rehfeldt等,2001)。 另外,Gatford 等(2010)指出母体生长激素对经产母猪的影响较初产母猪更明显,由于初产母体需要营养来维持生长限制了胎盘对GH的应答,可以认为甜菜碱能够促进母体GH的分泌或增加GH的分泌方式,从而增加高胎龄母猪的窝产仔数。
4 结论
本试验研究表明,添加B族维生素可以增加低胎龄母猪的窝产仔数和减少各胎龄母猪早期胚胎流产发生率。低胎龄母猪因缺乏B族维生素,其繁殖性能受到影响。另外,在高胎龄母猪日粮中添加甜菜碱能够提高其窝产仔数。
(致谢:本项目由CRC的Internationally Competitive Pork Industry提供资金支持,项目编号:2D-129,在此表示衷心感谢。同时对Paul Herde和David Cadogan博士为本试验提供的帮助表示谢意。)
[1]Armstrong D T.Effects of maternal age on oocyte developmental competence[J].Theriogenology,2001,55:1303 ~ 1322.
[2]Di Simone N,Riccardi P,Maggiano N,et al.Effect of folic acid on homocysteine-induced trophoblast apoptosis[J].Molecular Human Reproduction,2004,10:665 ~ 669.
[3]Duquette J,Matte J J,Farmer C,et al.Pre-and postmating dietary supplements of folic acid and uterine secretory activity in gilts[J].Canadian Journal of Animal Science,1997,77:415 ~ 420.
[4]Foxcroft G R,Vinsky M D,Paradis F,et al.Macroenvironment effects on oocytes and embryos in swine[J].Theriogenology,2007,68(1),:S30 ~ S39.
[5]Gatford K L,De Blasio M J,Roberts C T,et al.Responses to maternal GH or ractopamine during early-mid pregnancy are similar in primiparous and multiparous pregnant pigs[J].The Journal of Endocrinology,2009,203:143 ~154.
[6]Gatford K L,Smits R J,Collins C L,et al.Maternal responses to daily maternal porcine somatotropin injections during early-mid pregnancy or earlylate pregnancy in sows and gilts[J].Journal of Animal Science,2010,88:1365 ~1378.
[7]Guay F,Matte J J,Girard C L,et al.Effects of folic acid and vitamin B12 supplements on folate and homocysteine metabolism in pigs during early pregnancy[J].The British Journal of Nutrition,2002a,88:253 ~ 263.
[8]Guay F,Matte J J,Girard C L,et al.Effect of folic acid and glycine supplementation on embryo development and folate metabolism during early pregnancy in pigs[J].Journal of Animal Science,2002b,80:2134 ~ 2143.
[9]Guay F,Matte J J,Girard C L,et al.Effect of folic acid plus glycine supplement on uterine prostaglandin and endometrial granulocyte-macrophage colony-stimulating factor expression during early pregnancy in pigs[J].Theriogenology,2004,61:485 ~ 498.
[10]Harper A F,Lindemann M D,Chiba L I,et al.An assessment of dietary folic acid levels during gestation and lactation on reproductive and lactational performance of sows:a cooperative study.S-145 Committee on Nutritional Systems for Swine to Increase Reproductive Efficiency[J].Journal of Animal Science,1994,72:2338 ~ 2344.
[11]Holmes V A.Changes in haemostasis during normal pregnancy:does homocysteine play a role in maintaining homeostasis?[J].Proceedings of the Nutrition Society,2003,62(2):479 ~ 493.
[12]Huang Q C,Xu Z R,Han X Y,et al.Effect of betaine on growth hormone pulsatile secretion and serum metabolites in finishing pigs[J].Journal of Animal Physiology and Animal Nutrition,2007,91:85 ~ 90.
[13]Ikeda S,Namekawa T,Sugimoto M,et al.Expression of methylation pathway enzymes in bovine oocytes and preimplantation embryos[J].Journal of Experimental Zoology.Part A,Ecological Genetics and Physiology,2010,313A:129 ~ 136.
[14]Kidd M T,Ferket P R,Garlich J D.Nutritional and osmoregulatory functions of betaine.World’s[J].Poultry Science Journal,1997,53:125 ~ 139.
[15]Kim E S,Seo J S,Eum J H,et al.The effect of folic acid on in vitro maturation and subsequent embryonic development of porcine immature oocytes[J].Molecular Reproduction and Development,2009,76:120 ~ 121.
[16]Kwong W Y,Adamiak S J,Gwynn A,et all.Endogenous folates and single-carbon metabolism in the ovarian follicle,oocyte and pre-implantation embryo[J].Reproduction(Cambridge,England),2010,139:705 ~ 715.
[17]Lindemann M D,Kornegay E T.Folic acid supplementation to diets of gestating-lactating swine over multiple parities[J].Journal of Animal Science,1989,67:459 ~ 464.
[18]Mahan D C.Relationship of gestation protein and feed intake level over a five-parity period using a high-producing sow genotype[J].Journal of Animal Science,1998,76:533 ~ 541.
[19]Matte J J,Girard C L.An estimation of the requirement for folic acid in gestating sows:the metabolic utilization of folates as a criterion of measurement[J].Journal of Animal Science,1999,77:159 ~ 165.
[20]Matte J J,Guay F,Girard C L.Folic acid and vitamin B12in reproducing sows:new concepts[J].Canadian Journal of Animal Science,2006,86:197 ~205.
[21]Murphy M M,Scott J M,McPartlin J M,et al.The pregnancy-related decrease in fasting plasma homocysteine is not explained by folic acid supplementation,hemodilution,or a decrease in albumin in a longitudinal study[J].The American Journal of Clinical Nutrition,2002,76:614 ~ 619.
[22]National Research Council.‘Nutrient requirements of swine.’ 10th edn[S].National Academy Press:Washington DC,1998.
[23]Rehfeldt C,Kuhn G,Nurnberg G,et al.Effects of exogenous somatotropin during early gestation on maternal performance,fetal growth,and compositional traits in pigs[J].Journal of Animal Science,2001,79:1789 ~ 1799.
[24]Simard F,Guay F,Girard C L,et al.Effects of concen-trations of cyanocobalamin in the gestation diet on some criteria of vitamin B12 metabolism in first-parity sows[J].Journal of Animal Science,2007,85:3294 ~3302.
[25]Solanky N,Requena Jimenez A,D’Souza S W,et al.Expression of folate transporters in human placenta and implications for homocysteine metabolism[J].Placenta,2010,31:134 ~ 143.
[26]Stahly T S,Williams N H,Lutz T R,et al.Dietary B vitamin needs of strains of pigs with high and moderate lean growth[J].Journal of Animal Science,2007,85:188 ~ 195.
[27]Thaler R C,Nelssen J L,Goodband R D,et al.Effect of dietary folic acid supplementation on sow performance through two parities[J].Journal of Animal Science,1989,67:3360 ~ 3369.
[28]Town S C,Patterson J L,Pereira C Z,et al.Embryonic and fetal development in a commercial dam-line genotype[J].Animal Reproduction Science,2005,85:301 ~ 316.
[29]Tremblay G F,Matte J J,Dufour J J,et al.Survival rate and development of fetuses during the first 30 days of gestation after folic acid addition to a swine diet[J].Journal of Animal Science,1989,67:724 ~ 732.
[30]Vallet J L,Christenson R K,Klemcke H G.A radioimmunoassay for porcine intrauterine folate binding protein [J].Journal of Animal Science,1999,77:1236 ~ 1240.
[31]van Mil N H,Oosterbaan A M,Steegers-Theunissen R P.Teratogenicity and underlying mechanisms of homocysteine in animal models:a review[J].Reproductive Toxicology,2010,30:520 ~ 531.
[32]van Wettere W H E J,Herde P.Supplementing gestation diets with betaine increases litter size of summer mated sows[N].In‘Manipulating pig production 12’,2009,140.
[33]van Wettere W H E J,Mitchell M,Revell D K,et al.Management and nutritional factors affecting puberty attainment and first litter size in replacement gilts[N].In ‘Manipulating pig production X’,2005,180 ~ 192.
[34]Velzing-Aarts F V,Holm P I,Fokkema M R,et al.Plasma choline and betaine and their relation to plasma homocysteine in normal pregnancy[J].The American Journal of Clinical Nutrition,2005,81:1383 ~ 1389.
[35]Xie S,Broermann D M,Nephew K P,et al.Ovulation and early embryogenesis in swine[J].Biology of Reproduction,1990,43:236 ~ 240.
