冰片提取物对沙眼衣原体感染Hela细胞CT703及CT259表达的影响
2015-12-25郭翠玲,赵仲平,汤爱灿等
冰片提取物对沙眼衣原体感染Hela细胞CT703及CT259表达的影响
郭翠玲赵仲平汤爱灿欧波
(海南省中医院眼科中心,海南海口570203)
摘要〔〕目的探讨冰片提取物对沙眼衣原体感染Hela细胞CT703及CT259表达的影响。方法建立沙眼衣原体感染Hela细胞模型,将被感染后的Hela细胞放入A、B两培养瓶中培养,其中A培养瓶培养基中加入冰片提取物0.3 g,而B培养瓶只是加入等量的培养基,采用RT-PCR及 Western印迹法检测选取培养6、12、24 h后Hela细胞中CT703及CT259基因及蛋白表达水平。结果A培养瓶加入冰片提取物后,被感染的Hela细胞CT703、CT259 mRNA表达水平随着培养时间的延长而逐渐降低(P<0.05);B培养瓶中被沙眼衣原体感染的Hela细胞CT703、CT259 mRNA表达水平随着培养时间的延长未出现明显变化(P>0.05)。A培养瓶加入冰片提取物后,被感染的Hela细胞CT703、CT259蛋白表达水平随着培养时间的延长而逐渐降低(P<0.05);B培养瓶中被沙眼衣原体感染的Hela细胞CT703、CT259蛋白表达水平随着培养时间的延长未出现明显变化,差异无统计学意义(P>0.05)。结论被沙眼衣原体感染的Hela细胞中的CT703及CT259基因及蛋白表达均处于高水平,冰片提取物能够明显抑制Hela细胞中CT703及CT259基因及蛋白表达,临床能够用于沙眼衣原体感染的治疗。
关键词〔〕冰片提取物;沙眼衣原体;Hela细胞;CT703;CT259
中图分类号〔〕R285〔文献标识码〕A〔
第一作者:郭翠玲(1972-),女,副主任医师,主要从事眼底疾病临床与基础研究。
沙眼衣原体(Ct)是一种具有细胞内二相性生活周期的严格细胞内寄生的微生物,内含代谢活跃、无感染性的网状体和感染性强、代谢不活跃的原体。沙眼衣原体感染人后最常见是引起沙眼,还会引起包涵体结膜炎、输卵管炎、宫颈炎以及一些性传染性疾病,有关研究报道,人类生殖道感染沙眼衣原体后能够增加一些传染性疾病和生殖系统疾病的概率,如艾滋病、宫颈癌等〔1〕。沙眼衣原体能够干扰宿主细胞信号传导途径,从而影响生物学行为,躲避宿主对其免疫,导致机体持续处于感染状态〔2〕。在持续感染状态下,衣原体在宿主细胞内不再继续复制增殖,处于代谢率低下状态,但网状体将异常增大,很难被机体剔除,当宿主免疫系统出现紊乱或者低下或者其他微生物侵袭宿主时,持续感染状态下的衣原体能够恢复期正常的感染性,再次分化感染性强的原体。沙眼衣原体持续感染是致宿主发生疾病的一个重要方面。本文探究沙眼衣原体基因CT703、CT259在持续感染时的表达水平变化以及冰片提取物对沙眼衣原体的作用。
1资料与方法
1.1材料冰片提取物购于陕西钰堂生物科技发展有限公司;人宫颈癌Hela细胞株购于中国医学科学院基础研究所;沙眼衣原体购于美国德克萨斯州健康科学中心;RPMI-1640培养基及Trizol试剂均购于购于美国Gibco公司;RT-PCR试剂盒、超纯RNA提取试剂盒和逆转录试剂盒均购于美国Promega公司;兔抗人CT703抗体、兔抗人CT259抗体、引物、Tap酶和DAB显色剂购于Cell Signalling Technology 公司。
1.2Hela细胞培养无菌操作下,取人宫颈癌Hela细胞置于RPMI-1640培养基中,在37℃体积分数为5%的CO2饱和湿度环境中培养,培养24 h至生长对数期,用0.25%胰酶消化细胞,10%体积分数的小牛血清培养基终止消化,按照1∶3传代培养,每3天传代培养一次。
1.3沙眼衣原体感染模型建立预先置有玻片的24孔板内加入1 ml浓度为2×105/ml的人宫颈癌Hela细胞悬液,置于37℃体积分数为5%的CO2饱和湿度环境中培养,24 h长成单层后,抽取并除去上清液,向每孔加入2 μl感染复数为1的沙眼衣原体悬液,在适宜环境中培养2 h后,除去上清液,并加入适量的RPMI-1640培养基继续培养。在沙眼衣原体感染后24、48 h后取出玻片,采用姬姆萨染色成功,于光学显微镜及电子显微镜下观察。
1.4分组及方法向预先准备好的A、B两个50 cm3培养瓶中分别加入浓度为2×105/ml的人宫颈癌Hela细胞悬液10 ml,置于37℃体积分数为5%的CO2饱和湿度环境中培养,24 h长成单层后,抽取并除去上清液,除去上清液,向每个培养瓶中加入50 μl感染复数为1的沙眼衣原体悬液,在适宜环境中培养2 h后,除去上清液,并加入适量的RPMI-1640培养基继续培养。培养24 h后,向A培养瓶加入0.3 g冰片提取物,向B培养瓶加入等量RPMI-1640培养基,置于适宜环境中继续培养,分别取培养6、12、24 h后细胞备用。
1.5RT-PCR法检测不同时间段Hela细胞CT703、CT259基因表达水平
1.5.1提取总RNA分别取A、B两培养瓶不同时间段Hela细胞置于0.2%胶原酶中消化,采用D-Hanks液清洗,4℃水中30 min,1 200 r/min离心8 min,制备成单细胞沉淀,向单细胞沉淀中加入1 ml Trizol试剂,采用超纯RNA提取试剂盒提取Hela细胞中总RNA。实验操作按产品说明书进行,用紫外分光光度法测定RNA的OD260/OD280确定总RNA纯度。
1.5.2RT-PCR根据逆转录试剂盒要求进行反转录反应操作,按照相关文献资料设计PCR中内参照CT110、CT703和CT259引物,将RNA模板、引物、5倍缓冲液和RNase-free水溶解并置于冰上备用,将2 μl引物混合物试剂和10 μl消化后RNA分别加入20 μl反应体系的第一部分,混匀。PCR反应体系经过PCR反应条件的优化,PCR循环条件为94℃30 s,55℃30 s,72℃1 min,共35个循环,最后72℃再延伸5 min。引物序列:CT259-正义链5′-TGGAATTCGGATTGTTGCGGATTTCG-3′,反义链5′-CTCGAGCGTTAACGCATCCGAACCAG-3′,产物747 bp;CT703-正义链5′-ACTCGTAAGGTTTCGTCA-3′,反义链5′-CTCGTAAATAAATGGGACTT-3′,产物352 bp;CT110-正义链5′-CTAATGAGTTGACCGCCAGT-3′,反义链5′-GACGCTTTGGTTCTAATCTACGA-3′,产物323 bp。
1.5.3结果分析PCR产物经1.2%琼脂糖凝胶电泳后,溴乙啶染色,通过成像分析scion软件计算和比较CT703和CT259产物条带的吸光度值,并与CT110条带光密度值比较,计算出CT703和CT259在Hela细胞中mRNA表达含量相对值。
1.6Western印迹法检测不同时间段Hela细胞CT703、CT259蛋白表达无菌操作下,分别取A、B两培养瓶不同时间段Hela细胞,将选取好的Hela细胞经PBS缓冲液洗涤,洗涤后离心收集,用冰冷PBS洗涤细胞3次后,加入裂解缓冲液,摇匀,置于4℃水中冰浴30 min,1 200 r/min离心10 min,吸取上清液,即胞质蛋白。采用BCA法对样品蛋白质进行蛋白定量。调整样品蛋白量为30~40 μg后进行SDS-PAGE凝胶电泳分离,并将蛋白质转移到PVDF膜上。PBST洗膜5 min后,PBST溶解封闭PVDF膜2 h。继而加入兔抗人CT703抗体、兔抗人 CT259抗体(按1∶1 000稀释)或抗β-actin(1∶2 000稀释),置于4℃环境中过夜。PBST洗涤3次,每次10 min,加入二抗,温室孵育2 h。再次用PBST洗涤3次,每次10 min。将PVDF膜ECL显色,在暗室中将PBVF曝光于X光片中,用凝胶成像系统扫描分析结果。
1.7统计学方法采用SPSS17.0进行t检验。
2结果
2.1总RNA提取结果紫外分光光度法测定中可知Hela细胞CT703、CT259 RNA OD260/OD280值在1.8~2.0之间,在1.2%的琼脂糖凝胶电泳可观察到28S、18S、5S三条产物条带,说明提取Hela细胞中的总RNA完整性较好。如图1。
2.2不同时间段Hela细胞CT703、CT259 mRNA表达比较A培养瓶加入冰片提取物后,被感染的Hela细胞CT703、CT259 mRNA表达水平随着培养时间的延长而逐渐降低,且培养24 h后Hela细胞CT703、CT259 mRNA表达水平明显低于培养6 h(P<0.05);而B培养瓶中被感染的Hela细胞CT703、CT259 mRNA表达水平随着培养时间的延长未出现明显变化(P>0.05);A培养瓶培养24 h后Hela细胞CT703、CT259 mRNA表
达水平明显低于B培养瓶(P<0.01)。见表1,图2。
2.3不同时间段Hela细胞CT703、CT259蛋白表达比较A培养瓶加入冰片提取物后,被感染的Hela细胞CT703、CT259蛋白表达水平随培养时间延长而逐渐降低,且培养24 h后Hela细胞CT703、CT259蛋白表达水平明显低于培养6 h(P<0.05);而B培养瓶中被感染的Hela细胞CT703、CT259蛋白表达水平随着培养时间的延长未出现明显变化,差异无统计学意义(P>0.05);A培养瓶培养24 h后Hela细胞CT703、CT259蛋白表达水平明显低于B培养瓶(P<0.01)。见表2,图3。
图1 RNA样品凝胶电泳结果

图2 沙眼衣原体感染Hela细胞不同时间点CT259、CT703、 CT110 PCR产物琼脂糖凝胶电泳图

组别CT7036h12h24hCT2596h12h24hA培养瓶0.64±0.110.51±0.130.34±0.061)2)0.71±0.120.43±0.120.31±0.041)2)B培养瓶0.66±0.050.71±0.120.69±0.110.72±0.130.77±0.110.78±0.12
与培养6 h比较:1)P<0.05;与B培养瓶比较:2)P<0.01,下表同

表2 不同时间段Hela细胞CT703、CT259蛋白表达比较
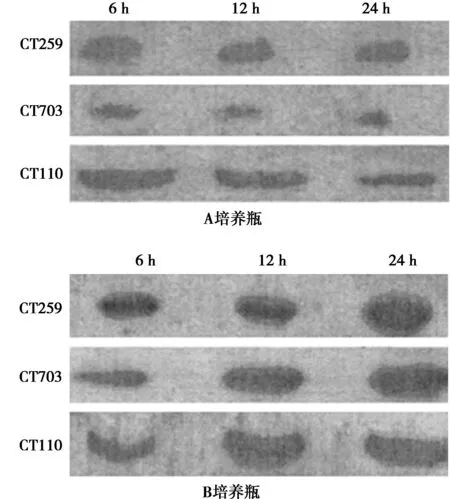
图3 沙眼衣原体感染Hela细胞不同时间点CT259、CT703、 CT110蛋白表达
3讨论
沙眼衣原体寄生宿主细胞后期代谢活跃,无感染性的网状体在细胞内逐渐增大,当宿主免疫功能出现低下或其他微生物侵袭宿主时,衣原体会趁机恢复其二相性生活周期,分化成具有强传染性的原体,从而导致宿主发生病变〔3〕。研究表明,沙眼衣原体持续感染状态下基因转录发生很大的变化,与信号传导有关的基因会随着沙眼衣原体寄生宿主的状态而发生改变,转变或上调或下调〔4〕。
本研究结果显示,当感染一定时间后,随着时间后的延长Hela细胞中CT259、CT703基因及蛋白表达量为出现明显变化,说明持续感染状态下,沙眼衣原体在宿主细胞内处于休眠状态,维持无感染性的网状体生活相,以保护沙眼衣原体的活性。当宿主机体处于低水平的免疫状态或者其他微生物侵袭时,沙眼衣原体将恢复其感染状态,分化具有强感染性的原体,破坏宿主细胞、组织从而导致疾病发生〔5~7〕。本研究说明冰片提取物能够协助宿主免疫系统清除因持续感染而逐渐增多的沙眼衣原体网状体,从而有效阻止沙眼衣原体所导致宿主细胞、组织损伤发生疾病。冰片提取物主要成分为龙脑、异龙脑、右旋龙脑等化合物〔8〕。龙脑、异龙脑均具有耐缺氧作用,具有较好的镇静、止痛功效,较高浓度的龙脑及异龙脑对葡萄球菌、链球菌、肺炎双球菌、大肠杆菌以及皮肤真菌等均有较强的抑制作用〔9~11〕。冰片提取物其化学成分能够激活宿主的巨噬细胞、自然杀伤细胞活性,提高单核细胞的循环和免疫细胞表面抗原和抗体的表达,增强宿主免疫系统对沙眼衣原体网状体的识别和清除,有效阻止了网状体转化为具有感染性、致病性的原体〔12〕。
综上所述,沙眼衣原体寄生宿主细胞后干扰细胞信号传导,躲避宿主免疫系统的清除,进而转化为无感染性的网状体,
使沙眼衣原体感染后Hela细胞中CT259、CT703基因及蛋白表达处于高水平状态。冰片提取物通过增强对网状体的识别和宿主免疫系统,有效的清除长期寄生于细胞内的沙眼衣原体。冰片提取物能够清除机体内寄生的沙眼衣原体,有利于提高沙眼衣原体感染所致疾病的治疗,对临床具有重要的指导意义。
4参考文献
1Rita Ferreira,Vitor Borges,Alexandra Nunes,etal.Assessment of the load and transcriptional dynamics of Chlamydia trachomatis plasmid according to strains' tissue tropism〔J〕.Microbiol Res,2013;168(6):333-9.
2Althaus CL,Turner KME,Schmid BV,etal.Transmission of Chlamydia trachomatis through sexual partnerships:a comparison between three individual-based models and empirical data〔J〕.J Royal Soc Interf,2012;9(66):136-46.
3Won BY,Lee DW,Shin SC,etal.A DNA intercalation-based electrochemical method for detection of Chlamydia trachomatis utilizing peroxidase-catalyzed signal amplification〔J〕.Biosensors Bioelectronics,2008;24(4):665-9.
4Kawana K,Quayle AJ,Ficarra M,etal.CD1d degradation in Chlamydia trachomatis-infected epithelial cells is the result of both cellular and chlamydial proteasomal activity〔J〕.J Biol Chem,2007;282(10):7368-75.
5Voevodskaya N,Galander M,Hogbom M,etal.Structure of the high-valent FeⅢFeⅣ state in ribonucleotide reductase (RNR) of Chlamydia trachomatis--combined EPR,57Fe-,1H-ENDOR and X-ray studies〔J〕.Biochim Biophy Acta Biomem,2007;1774(10):1254-63.
6Zheng Y,Zhao WM,Wang H,etal.Codon usage bias in Chlamydia trachomatis and the effect of codon modification in the MOMP gene on immune responses to vaccination〔J〕.Biochem Cell Biol,2007;85(2):218-26.
7陆洋,杜守颖,姚宗玲,等.天然冰片、合成冰片对栀子提取物黏膜促渗作用研究〔J〕.中国中药杂志,2009;34(10):1207-10.
8Vardhan H,Bhengraj AR,Jha R,etal.Higher expression of ferritin protects Chlamydia trachomatis infected HeLa 229 cells from reactive oxygen species mediated cell death〔J〕.Biochem Cell Biol,2010;88(5):835-42.
9Dessus-Babus S,Moore CG,Whittimore JD,etal.Comparison of Chlamydia trachomatis serovar L2 growth in polarized genital epithelial cells grown in three-dimensional culture with non-polarized cells〔J〕.Microb Infect,2008;10(5):563-70.
10Sun HS,Wilde A,Harrison RE,etal.Chlamydia trachomatis inclusions induce asymmetric cleavage furrow formation and ingression failure in host cells〔J〕.Mol Cell Biol,2011;31(24):5011-22.
11蔡冬梅,陈雪梅,殷胜利,等.通心络胶囊治疗原发性高血压病人眼底病变的疗效观察〔J〕.中国中医基础医学杂志,2002;8(9):51-2.
12伍海涛,唐由之,王宁生,等.含冰片眼用制剂的文献调查〔J〕.中国中医眼科杂志,2012;22(2):141-4.
〔2013-12-13修回〕
(编辑安冉冉/曹梦园)
