低温和低氧储藏对糙米发芽前后γ-氨基丁酸含量的影响
2015-12-18丁俊胄杨特武董梦钇熊善柏
丁俊胄 周 强 杨特武 董梦钇 张 璐 熊善柏
(华中农业大学食品科技学院1,武汉 430070)
(黄冈市农业科学院2,黄冈 438000)
(华中农业大学植物科技学院3,武汉 430070)
低温和低氧储藏对糙米发芽前后γ-氨基丁酸含量的影响
丁俊胄1周 强2杨特武3董梦钇1张 璐1熊善柏1
(华中农业大学食品科技学院1,武汉 430070)
(黄冈市农业科学院2,黄冈 438000)
(华中农业大学植物科技学院3,武汉 430070)
通过测定糙米在不同温度(-18、4、10℃、室温)和不同气体储藏环境(CO2、N2、空气、真空)下的发芽前后GABA含量、GAD和蛋白酶活力变化,以及储藏后糙米的发芽率、细胞膜透性、脂肪酸值、MDA值和淀粉粒微观形态,以确定生产高GABA发芽糙米的适宜储藏条件。结果表明,储藏早期(0~90 d),10℃储藏适于维持糙米中GABA含量,储藏60 d时糙米中GABA含量高出室温下64.46%;4℃和-18℃适于维持发芽糙米中GABA含量,储藏120 d时糙米发芽72 h后GABA含量分别高出室温下124.24%和43.59%。在N2和CO2环境中储藏可有效减缓糙米中GABA含量的下降,其中CO2较适于高GABA糙米的短期储藏,在30 d时高出空气环境下储藏的98.80%;低氧储藏有利于糙米发芽后GABA的积累,在N2、CO2、真空条件下储藏60 d时发芽糙米中GABA含量分别高出空气储藏下70.58%、55.39%、71.53%。
糙米 储藏 发芽 低氧 γ-氨基丁酸 脂质氧化
γ-氨基丁酸(γ-aminobutyric acid,GABA)主要是由谷氨酸脱羧酶(glutamic acid decarboxylase,GAD)催化L-谷氨酸(L-Glu)脱羧生成的,糙米发芽后蛋白酶、GAD等内源酶活性及游离氨基酸、GABA等小分子成分含量显著升高[1]。GABA在高等动物神经活动中起重要作用,具有活化脑神经,缓解焦虑,预防精神分裂和高血压等生理功效[2-3]。发芽糙米中的GABA含量与糙米品种、储藏条件及发芽工艺紧密相关,不同基因型和生态型糙米品种之间存在较大差异,早稻高于中晚稻[4]。此外,早籼米烹煮后口感较差,市场售价低廉,多用作米制品加工原料,因此采用早籼稻生产发芽糙米制品既可获得高GABA含量,又可大幅提升稻米加工附加值。由于在室温及有氧环境下,糙米胚及糊粉层中的脂类物质易被氧化,导致发芽率下降,保持糙米的发芽率及发芽后GABA含量对于高GABA米制品的生产至关重要。
低温储藏对延长稻米储藏期、维持鲜度效果显著,气调与真空储藏通过降低氧气含量对糙米陈化和霉变有显著抑制作用[5-6],同时,适宜的储藏温度和气调环境可保持烹制米饭的品质[7]。关于糙米储藏条件对发芽前后GABA含量影响的系统性研究尚未见报道。本试验以“两优287”为原料,测定在不同温度和气体环境下储藏270 d期间的发芽前后糙米GABA含量、GAD和蛋白酶活力,以及储藏后糙米的发芽率、浸出液电导率、游离脂肪酸值、丙二醛含量(MDA),探讨温度和气体环境对发芽糙米GABA积累的影响,确定适宜的糙米储藏条件。
1 材料与方法
1.1 试验材料
选用“两优287”为材料,属籼型两系杂交水稻,为湖北省典型早稻品种之一。
1.2 主要仪器和试剂
1.2.1 主要仪器设备
HP250GS型人工气候箱:武汉瑞华仪器设备有限责任公司;DZ400/ZD型充气/真空包装机(排氧量20 m3/h,极限真空度 1.333 kPa):上海余特包装机械制造有限公司;Hitachi S-4800型扫描电镜:日本日立公司。
1.2.2 主要试剂
γ-氨基丁酸:Sigma公司;磷酸吡多醛:北京拜尔迪生物公司;L-谷氨酸:上海伯奥生物科技有限公司。
1.3 主要试验方法
1.3.1 糙米的储藏包装
采用聚丙烯袋进行糙米样品的真空和充气包装。真空环境下在不同温度(-18、4、10℃、室温)和10℃下在不同气体环境(CO2、N2、空气、真空)下储藏,每次取样后立即重新进行真空和充气包装。
1.3.2 发芽糙米制备
采用本课题组自行设计改装的超声波水雾供湿通气培养装置(ZL201010221392.6),称取一定质量的糙米,按照文献[8-9]中的发芽方法进行,每间隔12 h使用纯水淋洗1次,待培养72 h时取样检测。
1.3.3 糙米发芽率的测定
参考 GB/T 5520—1985的方法测定糙米发芽率。
1.3.4 GABA含量的测定
[4]中的方法测定糙米及发芽糙米中的GABA含量。
1.3.5 GAD的提取及活力测定
综合参考文献[4,10]中的方法测定糙米及发芽糙米中的GAD活力。
1.3.6 蛋白酶活力的测定
参考SB/T 10317—1999的方法测定蛋白酶活力。
1.3.7 糙米浸出液电导率的测定
参考文献[11]中的方法,取5.0 g样品,去离子水冲洗一遍后用滤纸吸干,然后加入100 mL去离子水,25℃下浸泡12 h,测定浸泡开始时与结束时浸出液的电导率。
1.3.8 糙米游离脂肪酸值和MDA含量的测定
糙米充分粉碎后过40目筛得糙米粉,参考GB/T 15684—1995测定游离脂肪酸值,参考文献[12]中的方法测定MDA含量。
1.3.9 糙米内部形态的观察
使用ES-2030电镜专用真空冷冻干燥装置,真空度3E-3,样品喷金后于5 kV电压下在扫描电镜下选择观测点和合适的视野。
1.4 数据处理与分析
各组试验和各个指标测定3次平行,3次重复,应用Excel 2007绘制折线图,用SAS 8.1统计软件(SASSoftware Institute,Cary,NC)进行统计分析。
2 结果与分析
2.1 低温与低氧储藏对糙米GABA含量及GAD、蛋白酶活力的影响
2.1.1 GABA含量
如图1a所示,在30~150 d的储藏期内,10℃储藏的GABA含量显著高于其他温度条件(P<0.05),而-18℃和4℃储藏处理差异不显著;其中,在储藏60 d时10℃下储藏的糙米GABA高出室温储藏处理64.46%。如图1b所示,在储藏的前60 d内,在N2和CO2环境下储藏的糙米GABA含量降幅显著小于真空和空气环境储藏的糙米(P<0.05),其中以在CO2中储藏的糙米最优,在30 d时高出空气环境下98.80%,适于高GABA糙米的短期储藏。

图1 储藏温度和气体环境对糙米中GABA含量的影响
2.1.2 GAD活力
糙米在各低温、低氧条件储藏下的第2个月是GAD活力下降幅度最大的时间段。如图2a所示,在储藏60 d期间,在-18℃条件下的降幅最大,而在10℃下显著高于其他温度条件(P<0.05)。如图2b所示,在储藏的30~120 d期间,N2环境可显著减缓GAD活力的下降。

图2 储藏温度和气体环境对糙米中GAD活力的影响
2.1.3 蛋白酶活力
在各温度和气体环境下储藏,糙米蛋白酶活力在120 d后有显著下降(P<0.05),如图3 a所示,在室温条件下储藏30 d和60 d后蛋白酶活力显著低于其他温度,在4℃和10℃低温环境下储藏可以较好的维持糙米中蛋白酶活性,且二者差异不显著(P>0.05)。如图3 b所示,在空气环境下储藏糙米蛋白酶活力水平较低,90 d后活力显著低于无氧环境(P<0.05),蛋白酶活力受气调种类影响并不大。
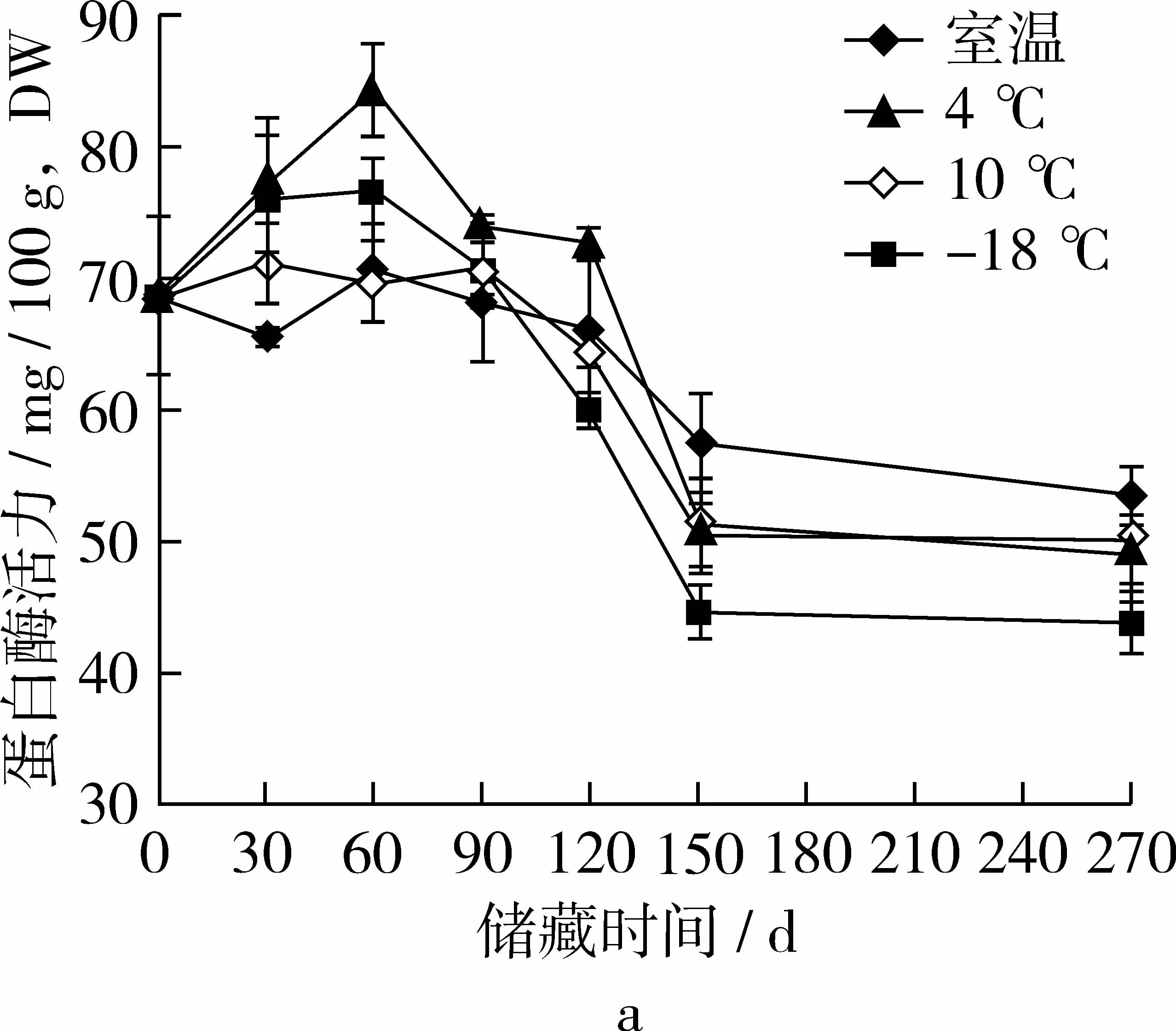

图3 储藏温度和气体环境对糙米中蛋白酶活力的影响
2.2 低温与低氧储藏对发芽糙米中GABA含量及GAD和蛋白酶活力的影响
2.2.1 GABA含量
如图4a所示,低温储藏条件的GABA含量显著高于室温(P<0.05),其中在-18℃和4℃条件下储藏120 d后,GABA的积累量较高,在120 d时分别高出室温下124.24%和43.59%。如图4 b所示,在储藏90 d内,对于维持发芽糙米GABA积累水平,CO2、N2或真空条件储藏的效果均优于空气储藏,在60 d时,N2、CO2、真空下储藏的发芽糙米GABA含量分别高出空气储藏下70.58%、55.39%、71.53%;在90~270 d间,CO2储藏环境显著优于N2(P<0.05)。整体而言,低温及低氧环境可以较好的维持发芽糙米中GABA积累水平,而在室温和空气条件下活力水平较低。
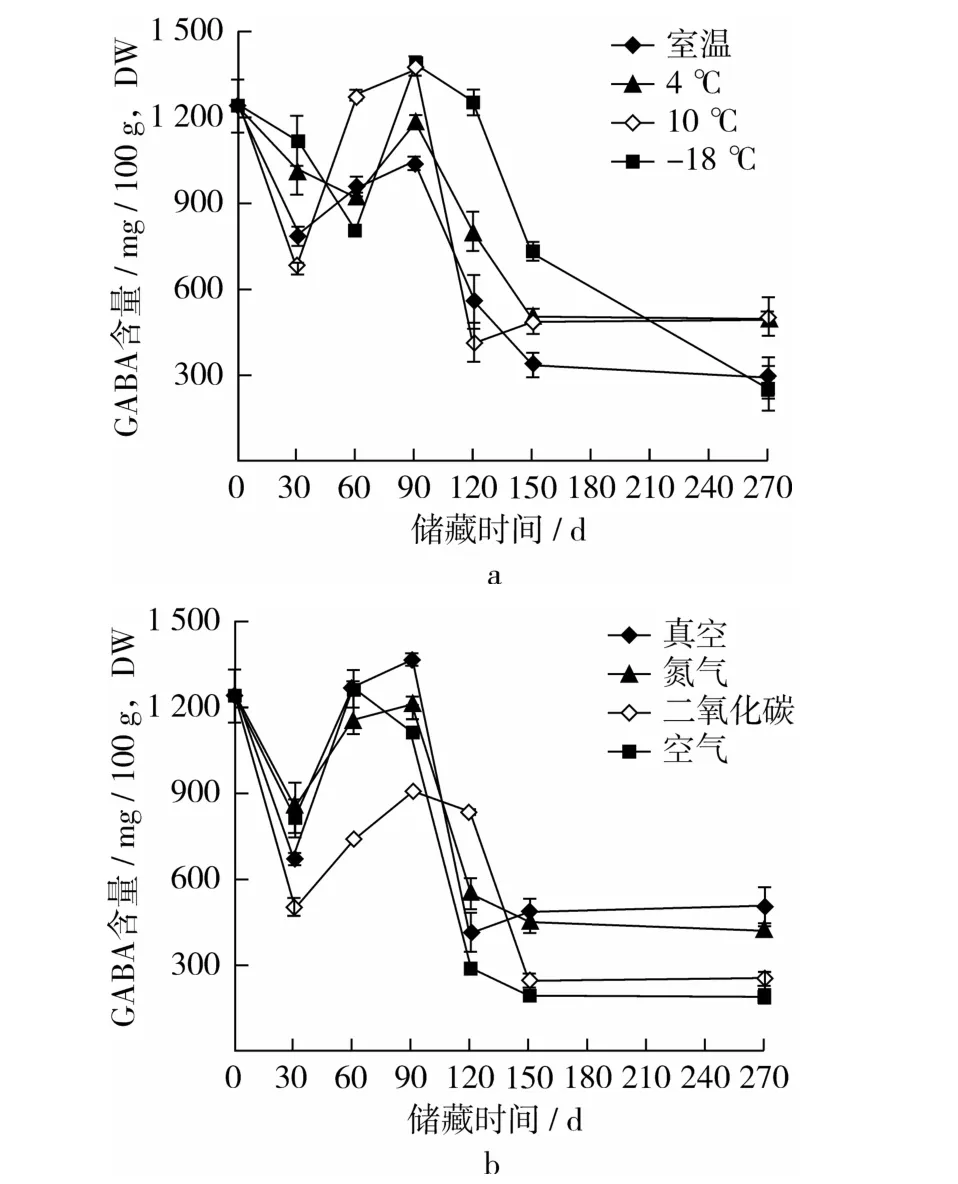
图4 储藏温度和气体环境对糙米发芽72 h后GABA含量的影响
2.2.2 GAD活力
如图5a所示,在10、4和-18℃下储藏可显著促进糙米发芽后GAD活力的上升(P<0.05),10℃下的增长发生在储藏30~60 d期间,4和-18℃下则发生在90~150 d期间。如图5b所示,低氧环境显著优于空气环境(P<0.05),在空气条件下储藏30 d后 GAD活力回升较弱,而在真空、N2、CO2等低氧条件下都有明显回升。整体而言,低温及低氧环境可以较好促进糙米发芽后GAD活力的升高。

图5 储藏温度和气体环境对糙米发芽72 h后的GAD活力的影响
2.2.3 蛋白酶活力
如图6a所示,糙米在4和-18℃低温环境下储藏30 d内,发芽后蛋白酶活力显著升高(P<0.05),在-18℃下储藏增幅最大,但在储藏30 d后开始回落,至60 d后保持平稳。在储藏60 d后,-18℃环境下储藏的发芽糙米的蛋白酶活力显著高于其他温度环境(P<0.05);如图6 b所示,低氧环境显著高于空气环境(P<0.05),在CO2和N2环境下,在储藏的前30 d内糙米在发芽后蛋白酶活上升,随后下降,其中在CO2环境下储藏60 d后蛋白酶活降幅较大。

图6 储藏温度和气体环境对糙米发芽72 h后蛋白酶活力的影响
2.3 低温与低氧储藏对糙米种子生命力及淀粉粒形态的影响
2.3.1 糙米发芽率和脂质氧化程度的影响
通常情况下糙米的发芽率与GABA在发芽后的积累能力成正相关性,本试验以储藏360 d后的糙米为材料研究不同温度和气体环境对糙米生命力的影响,测定结果见表1。在室温条件下储藏(1号样品)的脂肪酸值和电导率升值显著高于其他储藏环境(P<0.05),在空气有氧环境下储藏(7号样品)的MDA含量相对较高。
2.3.2 糙米胚的组织结构及近胚端储藏淀粉粒形态
使用扫描电镜观察了不同条件下储藏360 d的糙米胚表面、胚乳中复合淀粉粒形态,分别见图7中a1~a8、b1~b8。
由a1~a8可知,糙米胚由于受到致密层保护,在不同条件下储藏糙米胚表面形态差异不大,胚部均保持致密完整的表面形态,并未发生明显破损和穿孔。新鲜大米的淀粉粒多以复合粒形式存在,通过比较b1~b8,不同条件下储藏的胚乳内复合淀粉粒的形态差异并不明显,由b8可观察到饱满的淀粉粒均为棱角分明的多面体,且粒间排列致密,但b1~b7经过长期储藏后糙米胚乳内均出现了裂缝。这是由于胚乳细胞中赋予新米细胞壁较好弹性的多酚类化合物如阿魏酸在储藏过程中游离出来,以及细胞内部活性氧和自由基造成的破坏,致使大米细胞壁原有的富有弹性的网络结构被破坏,而陈化过程中产生的游离酚酸又是木质素的主要前体物质,这些物质可能聚合在一起,构成复杂的聚合体,形成典型的、具有相当硬度的陈米细胞组织,使细胞壁变脆。在不利储藏环境下稻米陈化后,胚乳中复合淀粉粒排列变疏松,折断面胚乳细胞破裂程度加大[13-14]。
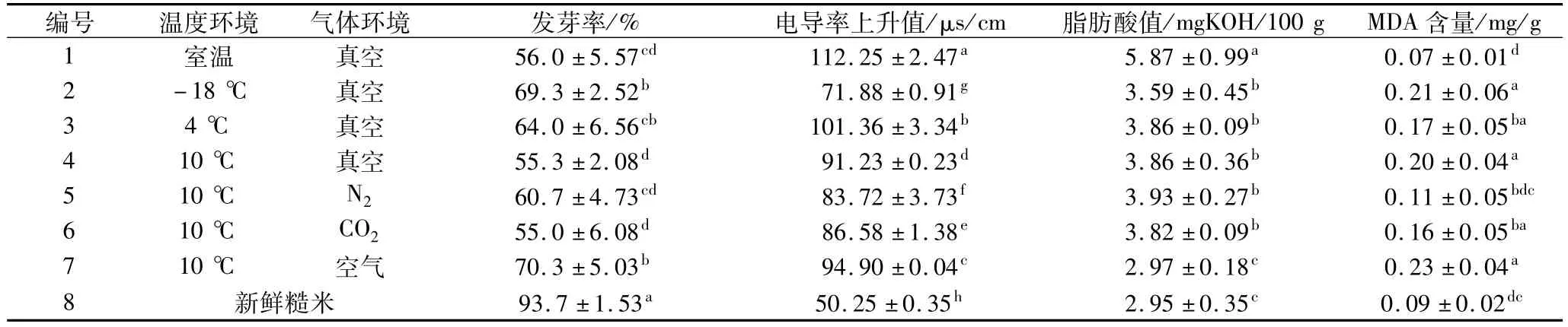
表1 温度和气体环境对储藏后糙米发芽率、浸出液电导率、脂肪酸值、MDA含量的影响
糙米的糊粉层和盾片是维持种子的重要物质库,含有大量的脂类和蛋白质,能为种子萌发提供必需的内源酶和内源激素,在高温和有氧环境下极易发生脂质氧化及过氧化反应,导致细胞的膜通道坏损[15]。由图7中c1~c4可知,室温条件下盾片附近淀粉粒上的微孔结构会因为陈化的发生而增多,加剧了氧化反应和种子发芽力的丧失。
3 讨论
由2.2.1和2.3.1中试验数据可知,适宜的温度和气体储藏环境可以维持糙米中GABA含量和发芽率。糙米发芽过程中GAD催化L-Glu脱去羧基生成GABA,其酶活力受Ca2+调控,Ca2+在细胞内外的流动对于GAD的影响极显著[16],同时激活后的蛋白酶水解糙米中的贮藏蛋白产生大量Glu为合成GABA提供底物[17]。在GAD催化GABA生成的同时,部分GABA被γ-氨基丁酸转氨酶(GABA-T)降解为琥珀酸半醛,只有当GAD活性大于GABA-T活性时,GABA才能实现有效积累[18]。低温[19]和低氧[20]环境可激发植物抗逆性,细胞内 Ca2+浓度增加,GAD活力上升,GAD与GABA-T的活性比值加大,GABA积累量显著提高,与本试验2.2.1和2.2.2中数据一致。本试验中低温和低氧环境下蛋白酶活力在0~30 d间显著上升,糙米需要30~60 d适应低温和低氧环境,在储藏早期(0~30 d)GABA含量、GAD活力出现小幅下降,在30~60 d间显著上升,并在60~90 d期间出现峰值,之后再缓慢降低。因此,进一步测定细胞内Ca2+浓度、Glu等关键氨基酸的含量,以及GABA-T活力的变化对于深入阐明环境对发芽糙米GABA积累影响规律和机制有重要意义。
糙米表面糊粉层以及内部脂类物质在储藏过程中因脂肪酶的水解作用生成游离脂肪酸,导致脂肪酸值升高[21]。储藏温度越高,其脂肪酸值上升速度越快[22],且在储藏温度超过20℃时,由于霉菌的生长繁殖分泌脂肪酶,会加快脂肪水解速度,脂肪酸值也随之增高[23]。脂肪酸在一定条件下可分解成醛和酮类物质,脂质过氧化反应的主要产物为MDA,MDA能降低生物膜的不饱和度和膜的流动性,使膜透性增大[24]。李宏洋等[11]报道了糙米在高温和有氧环境下长时间储藏,其种子膜系统会逐渐遭到破坏,导致糙米浸出液电导率增大。当胚部受损以及在高温有氧环境下长期贮藏过程中,过强的呼吸作用会导致稻谷种子陈化而使发芽力显著下降[25]。蒋家月等[26]研究报道了发芽率与游离脂肪酸值、电导率、MDA含量间呈负相关,糙米膜功能丧失可能是种子生命力下降的重要原因。
本研究表1中1号样品在室温条件储藏后脂肪酸值和糙米浸出液电导率升值均显著高于其他储藏环境下的样品(P<0.05),发芽率也相对较低;7号样品在低温(10℃)空气环境下储藏后,脂肪酸值相对较低,且具有较高的发芽率,尽管其MDA含量相对较高,但尚未对糙米种子生命力造成显著破坏。低温储藏可以有效抑制脂质氧化和过氧化反应,高温造成脂肪酸值升高和膜通透性下降与发芽活力的下降之间有着更直接的关联,而有氧环境下储藏尽管会导致脂质过氧化反应加剧,使MDA值升高,但10℃低温条件下不会直接造成糙米发芽活力的丧失。扫描电镜观察(图7)表明胚表面致密膜系统并未受储藏环境的影响,高温储藏环境可能加剧了内部糊粉层及盾片组织附近的脂肪氧化反应,对糙米细胞膜系统造成破坏,因而导致种子发芽率降低。
4 结论
在10℃下储藏,糙米中GABA含量可得到较好维持;低温、低氧环境在一定时间内能显著促进糙米发芽后GABA的积累,发芽糙米中GABA含量、GAD活力在储藏60~90 d期间出现峰值,然后逐渐下降,蛋白酶在低温和低氧环境下储藏30 d内显著升高,其中4℃和-18℃储藏更有助于发芽糙米中GABA的积累,且CO2、N2及真空所产生的低氧环境对GABA积累的影响存在差异。低温和低氧储藏环境对糙米胚表面膜系统致密性及胚乳中淀粉粒形态均无明显影响,室温储藏环境会加剧糊粉层及盾片组织内的脂肪氧化,对糙米细胞膜系统和发芽活性造成显著破坏。低温储藏能够有效减缓脂肪酸值升高和细胞膜通透性下降,维持糙米发芽活力,低氧环境可以有效减少脂质过氧化反应,抑制MDA值升高。
参考文献
[1]曹晓虹,温焕斌,李翠娟,等.糙米发芽过程中蛋白酶活性及含氮物质的变化[J].南京农业大学学报,2010,33(2):96-100
[2]Yoshimura M,Toyoshi T,Sano A,et al.Antihypertensive effect of a gamma-aminobutyric acid rich tomato cultivar“DG03-9”in spontaneously hypertensive rats[J].Journal of Agricultural and Food Chemistry.2010,58(1),615-619
[3]Palmer M L,Schulz M J,Murphy C S,et al.The Cellular basis of GABAB-mediated interhemispheric inhibition[J].Science,2012,335,989-993
[4]姚森,杨特武,赵莉君,等.发芽糙米中γ-氨基丁酸含量的品种基因型差异分析[J].中国农业科学,2008,41(12):3974-3982
[5]王玉凤,夏吉庆,孙培灵.糙米低温储藏技术的发展及应用前景[J].东北农业大学学报,2009,40(4):136-140
[6]王玉凤,孙培玲.糙米保鲜储藏技术试验研究[J].中国粮油学报,2009,24(5):95-98
[7]张兴亮,李喜宏,霍雨霞,等.温度对气调储藏水稻理化特性的影响[J].中国粮油学报,2010,25(5):77-80
[8]丁俊胄,刘贞,赵思明,等.糙米发芽过程中内源酶活性及主要成分的变化[J].食品科学,2011.32(11):29-32
[9]丁俊胄,刘贞,张璐,等.储藏期对发芽糙米富集γ-氨基丁酸的影响[J].中国粮油学报,2011,26(9):83-86
[10]Bartyzel I,Pelczar K,Paszkowski A.Functioning of theγ-aminobutyrate pathway in wheat seedlings affected by osmotic stress[J].Biologia Plantarum,2003,47,221-225
[11]李宏洋,王若兰,胡连荣.不同储藏条件下糙米品质变化研究[J].粮食储藏,2007(4):38-41
[12]侯福林.植物生理学试验教程[M].北京:科学出版社,2004:91
[13]周显青,张玉荣.储藏稻谷品质指标的变化及其差异性[J].农业工程学报,2008,24(12):238-242
[14]周显青,张玉荣,李里特.不同模拟储藏条件下粳米胚乳显微结构变化[J].农业工程学报,2010,26(5):329-334
[15]宋松泉,程红焱,姜孝成,等.种子生物学[M].北京:科学出版社,2008:145
[16]Zik M,Fridmann-Sirkis Y,Fromm H.C-terminal residues of plant glutamate decarboxylase are required for oligomerization of a high-molecular weight complex and for activation by calcium/calmodulin[J].Biochimica et Biophysica Acta,2006,1764(5),872-876
[17]付蓉.γ-氨基丁酸对作物幼苗生长发育的影响[D].南京:南京农业大学,2010
[18]Akihiro T,Koike S,TaniR,et al.Biochemicalmechanism on GABA accumulation during fruit development in tomato[J].Plant and Cell Physiology,2008,49,1378-1389
[19]Cholewa E,Andrzej JC,Shelp B J,et al.Cold-shockstimulatedγ-amirlobutynic acid synthesis ismediated by an increase in cytosolic Ca2+,not by an increase in cytosolic H+[J].Canadian Journal of Botany,1997,75,375-382
[20]Allan,W L,Peiris C,Bown A W,et al.Gammahydroxybutyrate accumulates in green tea and soybean sprouts in response to oxygen deficiency[J].Canadian Journal of Plant Science,2003,83,951-953
[21]周显青,杨文生,张玉荣.模拟典型储粮环境下大米品质指标的变化及其差异性分析[J].粮食与饲料工业,2009(5):1-5
[22]程建华,张蓉建,晏书明,等.糙米在不同储藏条件下品质变化的研究[J].粮食储藏,2003(3):42-46
[23]杨晓蓉,周建新,姚明兰,等.不同储藏条件下稻谷脂肪酸值的变化和霉变相关性的研究[J].谷物化学与分析品质,2006(5):49-52
[24]Suzuki Y.Screening and mode of inheritance of aricevarie lacking lipoxygenase-3[J].Gramma Field Symp,1995,33,51-62
[25]于勇,廖文艳,王俊.辐照稻谷种子的种胚微观结构特性及其萌发特性研究[J].中国粮油学报,2009,24(3):1-6
[26]蒋家月,宋美,吴跃进,等.水稻种胚脂肪氧化酶Lox-1,Lox-2缺失对种子储藏特性的影响[J].激光生物学报,2008,17(3):395-399
Effects of Low Temperature and Low Oxygen Storage on γ-Aminobutyric Acid Content of Brown Rice During Germination
Ding Junzhou1Zhou Qiang2Yang Tewu3Dong Mengyi1Zhang Lu1Xiong Shanbai1
(College of Food Science and Technology,Huazhong Agricultural University1,Wuhan 430070)
(Huang Gang Academy of Agricultural Science2,Huanggang 438000)
(College of Plant Science and Technology,Huazhong Agricultural University3,Wuhan 430070)
Brown rice has been stored at different temperature(4℃,10℃,-18℃,and room temperature)and gas environment(CO2,N2,air,and vacuum)respectively.The GABA content,GAD activity and protease activity of the brown rice and germinating brown rice have been determined during the storage process.The germination rate,cellmembrane system integrity,free fatty acid value,MDA content,Standard Electronic Modules(SEM)of the brown rice have also been analyzed after storage.The purpose of the study is to understand the appropriate storage conditions for a higher GABA production of brown rice after germination.The results showed that in the early stage of storage(0~90 d),the germinating brown rice from the brown rice stored in low temperature and low oxygen environment had their GABA content and GAD activity to be higher than those in room temperature and air environment.Under 10℃wasmore suitable tomaintain GABA content in germinating brown rice.GABA content will be 64.46%higher than that stored under room temperature for 60 d.Stored under4℃and-18℃were contributed to the accumulation of GABA in the germinating brown rice,with GABA content of124.24%and 43.59%higher than that under room temperature stored for 120 d respectively.Stored in the environment of N2and CO2,decreasing of GABA content could be effectively slowed down.The CO2addition wasmore suitable for short-term storage of brown rice with high GABA content.The GABA contentwas 98.80%higher than that stored in air environment for 30 d.Low oxygen storagewas beneficial for the GABA accumulation of germinating brown rice.On the condition of N2,CO2and vacuum,GABA content in germinating brown rice was79.58%,55.39%and 71.53%higher respectively than that stored in air for 60 d.
brown rice,storage,germination,low oxygen storage,GABA,lipid oxidation
S379.2
A
1003-0174(2015)01-0001-08
2013-11-03
丁俊胄,男,1988年出生,博士,食品科学
熊善柏,男,1963年出生,教授,水产品及淀粉质食品加工保鲜理论与技术
