塔里木河下游胡杨树腐烂病病原鉴定
2015-12-18郭开发吴彩兰方晓翠何丽赵思峰
郭开发,吴彩兰,方晓翠,何丽,赵思峰
(新疆绿洲农业病虫害治理与植保资源利用自治区高校重点实验室/石河子大学农学院,新疆 石河子 832003)
胡杨Populus euphratica Oliv是塔里木河下游干旱荒漠区天然分布的唯一乔木树种,对维护塔里木河下游地区的生态系统功能极为重要[1]。近50 a由于塔里木河流域上游水资源的大量开发,塔里木河下游断流320 km,胡杨林因严重干旱胁迫而大面积衰败[2],许多胡杨树发生腐烂病。庄文颖曾报道在中国西北地区胡杨腐烂病是由金黄壳囊孢Cytospora chrysosperma引起,该菌还可以导致灰胡杨、小叶杨、黑杨、欧美杨等几十种杨属植物的腐烂病[3]。另有研究依据形态学、ITS-rDNA gene和LSU gene测序结果认为,在新疆的杨属植物上还存在C.germanica,且该菌具有引起严重经济损失的潜在威胁[4]。为了明确塔里木河下游胡杨腐烂病的病原菌种类,以便进行准确检测与有效防控,本研究对2013年10月采自新疆巴音郭楞蒙古自治州铁门关市周边的胡杨腐烂病菌进行了鉴定。
1 材料与方法
1.1 材料2013年10月从新疆巴音郭楞蒙古自治州铁门关市周边采集胡杨树腐烂病样品12份,且病样有典型的黄褐色卷须状分生孢子角(图1-A)。
马铃薯葡萄糖培养基(PDA):去皮马铃薯200 g,葡萄糖20 g,琼脂18 g,水1000 mL,pH 6.8;
马铃薯葡萄糖液体培养基(PD):去皮马铃薯200 g,葡萄糖20 g,水1000 mL,pH 6.8;
水琼脂培养基(WA):琼脂20 g,水1000 mL。
1.2 病原分离与致病性测定
1.2.1 病原分离与纯化 采用常规组织分离法[5]在PDA平板上分离病原菌。待分离物长出后,在WA平板上单孢纯化,将纯化后获得的菌株保存在PDA斜面上,置4℃冰箱中储存备用。
1.2.2 致病性测定 采用离体枝条接种法[6]进行致病性测定。选取2年生健康的胡杨和新疆杨枝条,截取20 cm长的小段,自来水冲洗后用0.1%HgCl2浸泡10 min,然后用无菌水冲洗、晾干,枝条顶端用石蜡封口。用烧热的铁钉钉帽(直径7 mm)间隔一定距离烫伤树皮,把病原菌饼(直径7 mm)接种在烫伤处,在接种部位上方1~2 cm处加1块湿的脱脂棉,用保鲜膜将其和接种体同时包扎。每个分离株、每种寄主接5段枝条。每段枝条上3个接种点,随机选取1个伤口以接无菌PDA圆饼为空白对照。于10 d后观察、记录枝条发病情况,并对发病枝条进行菌物再分离。
1.3 病原物的鉴定
1.3.1 形态学鉴定 供试菌株接种在离体胡杨枝条上,待其产生产孢体后,挑取产孢体,用双面刀片直接切片,制成玻片后在生物显微镜下观察纵、横切面形态特征并测量分生孢子大小,记录并拍照。
1.3.2 分子鉴定 依据形态学鉴定结果,挑选TMG3,TMG5和TMG9这3株代表菌株在PDA平板上培养3 d后用打孔器打取直径为7 mm菌块接种至装有300 mL PD培养基的500mL的三角瓶中,28℃,200 r/min摇床培养3 d。用双层纱布过滤菌丝,灭菌水冲洗菌丝体至澄清,晾干,采用Lee等人的方法提取菌丝基因组DNA[7]。借鉴Peever等人[8]的方法,用通用引物ITS1和ITS4扩增代表菌株的ITS和5.8SrDNA,用引物Beta-F和Beta-R扩增代表菌株的β-tubulin基因。PCR反应体系:PCR TaqMix 10μL,10μmol/L ITS1/Beta-F 1μL,10μmol/L ITS4 /Beta-R 1μL,基 因 组DNA 0.5μL,ddH2O 12.5μL至终体积为25μL。ITS基因PCR扩增反应程序为94℃预变性4 min,然后94℃变性40 s,55℃退火1 min,72℃延伸1 min,共30个循环,最后72℃延伸10 min;β-tubulin基因PCR扩增反应程序为:94℃预变性3 min,然后94℃变性30 s,60℃退火30 s,72℃延伸1 min,共30个循环,最后72℃延伸10 min。反应完成后,1%琼脂糖凝胶电泳检测PCR反应产物。将不同引物的PCR反应产物进一步纯化后委托北京六合华大基因科技股份有限公司进行测序。测序后的序列经校正后,在核酸序列数据库GenBank中进行同源序列搜索(Blast搜索),比较测试菌株同现有数据库中相应序列的相似程度,用DNAMAN软件进行比对并构建系统树,应用自展法(Bootstrap)检验系统树,自展数据集为1000次。
2 结果与分析
2.1 病原菌的致病性测定人工接种2年生健康的胡杨和新疆杨枝条后,12个分离菌株均导致胡杨和新疆杨枝条发病。接种7 d后接种部位出现褐色、水浸状病斑。随后病斑纵向横向扩展至绕枝条一周。枝条病斑部位表皮皱缩,病健交界明显(图1-B,C)。接种10 d后,病斑表面产生大量黑色小点,室温较为干燥的条件下,产孢体孔口会溢出橘黄色的分生孢子角。对发病组织进行再分离,均可获得相应的接种菌株。对照枝条上未出现腐烂病症状。

图1 胡杨树腐烂病症状及致病性测定结果
2.2 病原物形态鉴定室内培养,菌落较紧密,菌落白色至乳白色,菌落背面米黄色。培养后期褐色产孢体随机在菌落分布(图2)。子座埋生在树皮中,突出,褐色,圆形至椭圆形,直径1.5~1.9 mm;孔口1个,黑色,与顶盘同高;分生孢子器由大量内陷的小腔室组成,共用壁,不规则排列呈玫瑰花座状;分生孢子透明、香蕉形,大小为(3.5~5.0)μm×(0.9~1.4)μm;分生孢子角橘黄色至橘红色(图3)。依据形态特征将病原菌鉴定为金黄壳囊孢Cytospora chrysosperma。
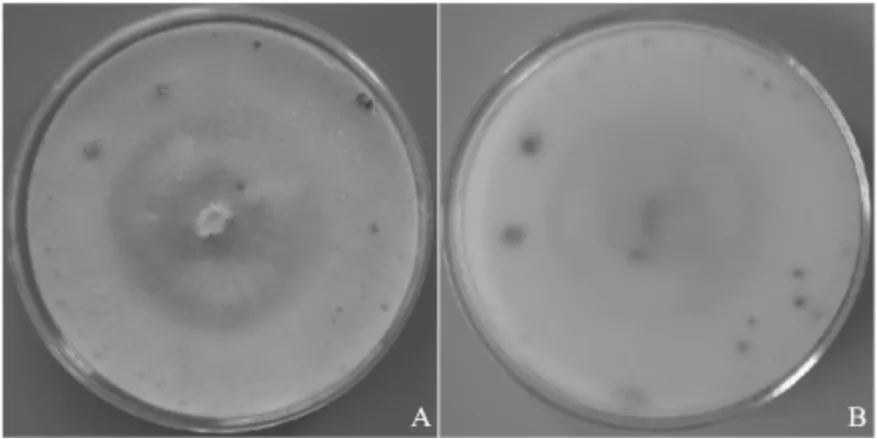
图2 C.chrysosperma在PDA培养基上菌落特征(A,正面;B,背面)
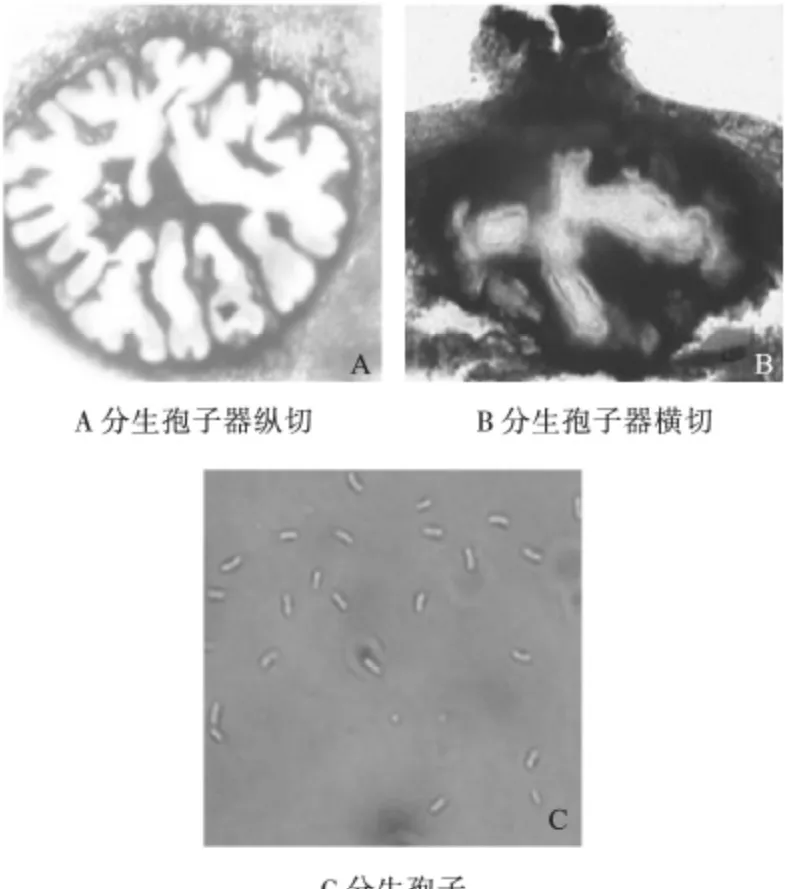
图3 Cytospora.chrysosperma形态特征
2.3 病原物分子鉴定分离菌株的DNA提取后经电泳检测,所得DNA片段约为2000 bp。利用ITS和β-tubulin两个基因对该病原菌进行PCR扩增,经电泳检测分别获得一条大小约为550 bp、500 bp的清晰条带(图4),将所测得的序列登陆NCBI(http://www.ncbi.nlm.nih.gov/blast)进行Blast比对。通过DNAMEN建立系统发育树(图5,6)。ITS和β微管蛋白序列分析:通过DNAMAN进行系统发育分析,通过最大简约法构建系统发育树。3个代表菌株的rDNA ITS序列(登录号分别为KP114124.1,KP114125.1和KP114126.1)与已登录的菌株C.chrysosperma(KJ739472)具有很高的Bootstrap支持率,相似率达到99%(图5)。3个代表菌株的β微管蛋白基因(登录号分别为KP117037.1,KP117038.1,KP117039.1)与已登录的菌株C.chrysosperma(JQ900372)达到98%以上的相似率(图6)。
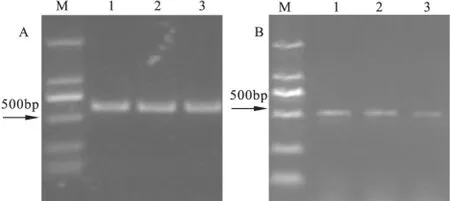
图4 胡杨腐烂病菌代表菌株PCR扩增结果(A,ITS;B,β-tubulin)

图5 代表菌株基于ITS序列构建的系统发育树

图6 代表菌株基于β-tubulin序列构建的系统发育树
3 讨论与结论
壳囊孢属Cytospora种类较多,分布范围广,种间的形态特征具有较大的相似性。按照传统的病原菌分类鉴定十分困难。ITS和β-tubulin序列分析用于壳囊孢属的分类和系统发育研究已被证明是可行的[9]。金黄壳囊孢危害杨树、柳树、桉树等多种林木树种,在西北、华北及东北地区是危害杨树生产的主要病原因子[10]。杨明秀 等认为金黄壳囊孢在我国培养性状上存在明显的遗传多样性,遗传多样性与地理来源有一定关系,但致病性多样性与地理来源无明显关系[11]。孙祥瑞对河北省白杨树腐烂病病原鉴定也为金黄壳囊孢[12]。目前已报道的能引起杨属主要林木腐烂病的病菌有金黄壳囊孢[3,12]和C.germanica。本研究表明新疆塔里木河下游胡杨树腐烂病的病原物为金黄壳囊孢,该菌与新疆已报道的杨树等树木上的腐烂病菌为一个种。致病性测定结果也表明,该菌既可侵染胡杨,也可侵染新疆杨,但该病原菌在两种林木上能否传播以及不同寄主来源的病菌是否存在致病性差异还有待于进一步研究。
[1] Pei Zhiqin,Xiao Chunwang,Dong Dan,et al.Comparison of the fine root dynamics of Populus euphratica forests in different habitats in the lower reaches of the Tarim River in Xinjiang,China,during the growing season[J].Journal of Forest Research,2012,17(4):343-351.
[2] 陈亚鹏,陈亚宁,徐长春,等.塔里木河下游地下水埋深对胡杨气体交换和叶绿素荧光的影响[J].生态学报,2011,31(2):0344-0353.
[3] Zhuang Wenying.Fungi of northwestern China[M].Ithaca:Mycotaxon Ltd,2005:1-430.
[4] Zhang Q T,He M,Lu Q,et al.Morphological and molecular identification of Cytospora germanica causing canker on Populus spp.in China[J].Plant disease,2013,97(6):846.
[5] 方中达.植病研究方法[M].3版.北京:中国农业出版社,1998:122-124.
[6] 臧睿,黄丽丽,康振生,等.陕西苹果树腐烂病菌(Cytospora spp.)不同分离株的生物学特性与致病性研究[J].植物病理学报,2007,37(4):342-251.
[7] Lee SB,Milgroom M G,Taylor J W.A rapid,high yield miniprep method for isolation of total genomic DNA from fungi[J].Fungal Genetics Newsletter,1988,35:23-24.
[8] Peever TL,Su G,Carpenter-Boggs L,et al.Molecular systema-tics of citrus-associated Alternaria species[J].Mycologia,2004,96(1):119-134.
[9] 杜琴,孔利,赵思峰,等.新疆库尔勒香梨腐烂病病原鉴定[J].新疆农业科学,2013,50(12):2258-2265.
[10] 张星耀,陈海燕,梁军,等.金黄壳囊孢菌(Cytospora chrysosperma)的培养性状和营养体亲和性[J].西北农林科技大学学报:自然科学版,2007,35(3):99-105.
[11]杨明秀,宋瑞清.中国金黄壳囊孢菌的致病性分化及遗传多样性[J].林业科学,2013,49(6):115-121.
[12]孙祥瑞.几种树皮腐烂病菌鉴定和生物学特性的研究[D].河北 保定:河北农业大学,2010.
