桔小实蝇与木瓜实蝇杂交后代产卵管长度的表型遗传特征
2015-12-09武目涛何日荣赵菊鹏任竞妹
马 骏,武目涛,梁 帆,何日荣,赵菊鹏,林 莉,任竞妹,张 灿
(广东出入境检验检疫局检验检疫技术中心,广州 510623)
桔小实蝇Bactrocera dorsalis s.s.和木瓜实蝇Bactrocera papayae 同属桔小实蝇复合种,两者均为为害水果的重要害虫(Clarke et al.,2005)。桔小实蝇复合种起源于东南亚地区,其中以桔小实蝇为代表种。1969年Hardy 在进行桔小实蝇及相关种的分类鉴定及其分布情况研究时指出,桔小实蝇与其它16种果实蝇在形态上属于近缘种,这些类群在形态上很难分辨,但彼此之间的生物学特性及各自寄主范围存在很大差别,提出过去被定为桔小实蝇的一些实蝇其实是由很多近缘种组成的复合种。1994年Drew 和Hancock 根据已收集的种类整理出桔小实蝇40 个复合新种,其中传统分类中的桔小实蝇被称之为“桔小实蝇狭义种”B.dorsalis s.s.。随后Allwoo 等(1999)记录了有害和无害种的寄主植物种类。每一新种根据传统的形态特征加以描述,同时根据同工酶、地理分布、寄主范围以及对性诱剂反应的差异性将其视为独立的生物单元,有的种类若仅仅依靠形态标准则很难区别(Clarke et al.2005)。自从上世纪80年代开始,随着一系列额外新种不断得到描述并加入,到目前桔小实蝇复合种达到75种。
木瓜实蝇根据其与桔小实蝇(狭义种)的差异性由Drew & Hancock 于1994年将其独立定名。两者依靠的主要鉴别特征为:(1)木瓜实蝇的产卵管明显比桔小实蝇长;(2)两者的地理分布不同,即桔小实蝇分布在亚洲大部分地区及东南亚,南至泰国/马来半岛上的克拉地海峡。木瓜实蝇从克拉地海峡开始,向南和向东扩展到印度尼西亚群岛。系统发育及种间条形码的研究显示这两个种实蝇在遗传上表现出了很近的遗传关系(Krosch et al.,2012;Schutze et al.,2012,2013)。有关桔小实蝇与木瓜实蝇是否为同一个种存在争议,至今也未得出明确的定论。根据实验观察,在室内条件下桔小实蝇和木瓜实蝇能正常交配并产生可育后代,两者之间并不存在生殖隔离。而根据Ernst Mayr (2005)对生物物种的概念 (the Biological Species Concept BSC):物种是一群实际发生交配或具有潜在交配能力的自然种群,该群体与其他群体存在生殖隔离。因此,鉴于桔小实蝇和木瓜实蝇的杂交现象,对两者的分类地位有必要回答的科学问题:(1)这两个种实蝇其本身是否属于生物学上的单一种?假如属于生物学上的单一种,其杂交后代种群繁殖的延续性如何?(2)即使按照此前所分的两个不同种实蝇,那么其分类所依据的产卵管长度经过杂交后是否还具有种间鉴别的形态特征?为验证在杂交条件下以产卵管长度作为上述两种实蝇形态鉴别特征的有效性,本研究对来源于各自地理分布区的上述两种实蝇经杂交后,对杂交后代实验种群连续饲养至第33 代并观测了杂交后代产卵管长度的表型遗传特征。
1 材料与方法
1.1 虫源和虫种鉴定
桔小实蝇采自广州番石榴果园,实验室饲养3 代。木瓜实蝇从马来西亚进境木瓜中截获得到,实验室饲养10 代以上。两种实蝇饲养于广东检验检疫技术中心植物检疫实验室。实蝇幼虫均用人工饲料饲养,由酵母、蔗糖、麦胚、水等配制。实蝇成虫性成熟后,用含有新鲜番石榴汁的产卵器收集虫卵,并转移至铺有滤纸的人工饲料上,于25℃±1℃,RH65%的条件下培养。继代繁殖中,每一代随机收集虫卵500 粒左右。幼虫老熟后用水洗出置于湿润的细沙中化蛹,蛹接近羽化时将蛹从沙中筛出并转移至养虫笼让其羽化。成虫提供食料(酵母∶糖=1∶3)和水。光源由40 W 白炽灯管提供,光照时间为上午8∶00 至晚上8∶00。
1.2 杂交方法与产卵管长度测定
两种实蝇种类鉴定根据Dew and Hancock(1994)描述的形态特征确认。F1 代由桔小实蝇与木瓜实蝇亲本杂交获得,由桔小实蝇(d)雌性与木瓜实蝇 (p)雄性产生的杂交一代标记为Fdp1,由木瓜实蝇雌性与桔小实蝇雄性产生的杂交一代标记为Fpd1。自F1 代开始,分别将交杂交种和其亲本持续繁殖至第33 代并随机收取该代杂交种及其亲本雌成虫36-186 头置于冰箱冷冻,采用显微镊在显微测微尺下解剖产卵针并测定其长度。
1.3 产卵管长度统计分析
采用DPS 数据处理系统(2013)统计分析杂交后代及其亲本产卵管长度的频次分布特征以及差异性。
2 结果与分析
2.1 杂交代与亲本产卵管长度的差异性
对测量的杂交种与其亲本产卵管长度进行方差分析,经方差齐性检验表明测量数据方差齐性一致(卡方值=2.13,df=3,P=0.5453)。4种遗传种系的产卵管长度均存在明显差异 (P<0.01),其中以木瓜实蝇的产卵管长度为最长,桔小实蝇的为最短,第33 代正反交杂交自交系产卵管长度介于两种亲本之间,并且正反交后代(来源于不同杂交亲本)之间的产卵管长度也具有明显差异(P<0.01)(表1)。4种系产卵管长度在95%的置信区间下互不重叠(表1)。
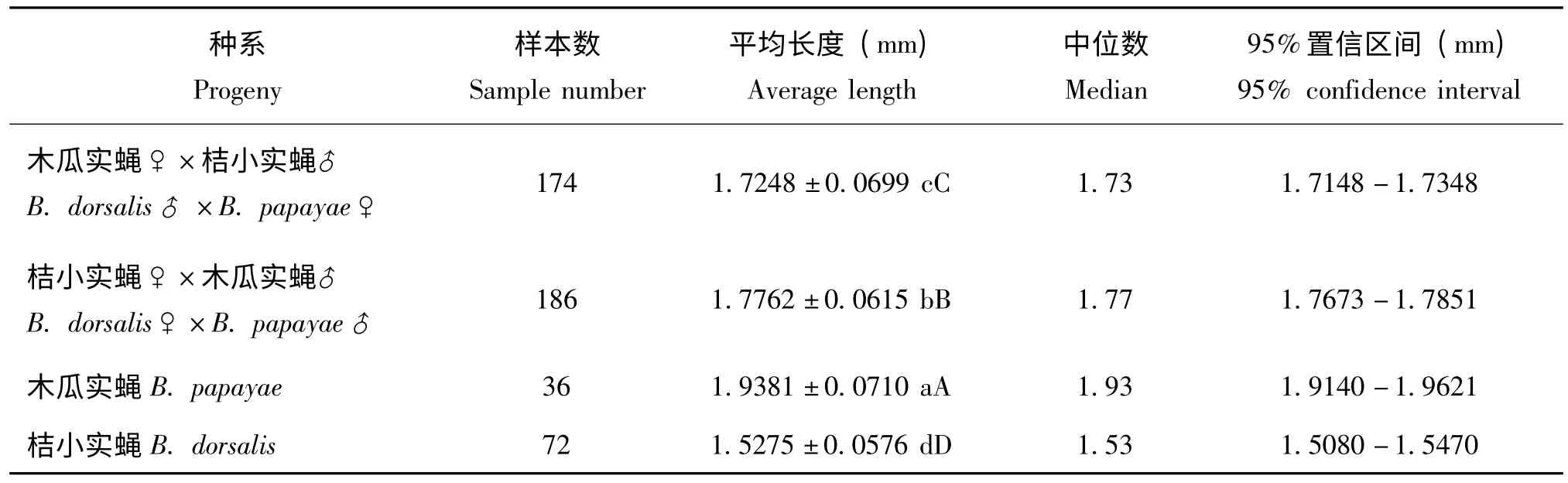
表1 桔小实蝇和木瓜实蝇及其杂交第33 代产卵管长度差异性分析Table 1 Length of aculeus for hybrid progeny between Bactrocera dorsalis s.s.and B.papayae after reproducing thirty-three generations
2.2 产卵管长度的频次分布特征
将第33 代杂交种及其亲本产卵管长度的分布频次进行分布函数拟合,经卡方检验均符合正态分布(图1),表明无论亲本还是杂交种其产卵管长度在一定的范围内为正态分布,产卵管长度的性状表现为完全随机性事件。
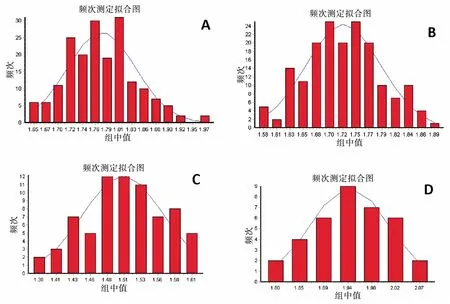
图1 桔小实蝇和木瓜实蝇及其杂交第33 代产卵管长度频次分布Fig.1 Frequency distribution of length of aculeus of hybrid progeny between Bactrocera dorsalis s.s.and B.papayae after reproducing thirty-three generations
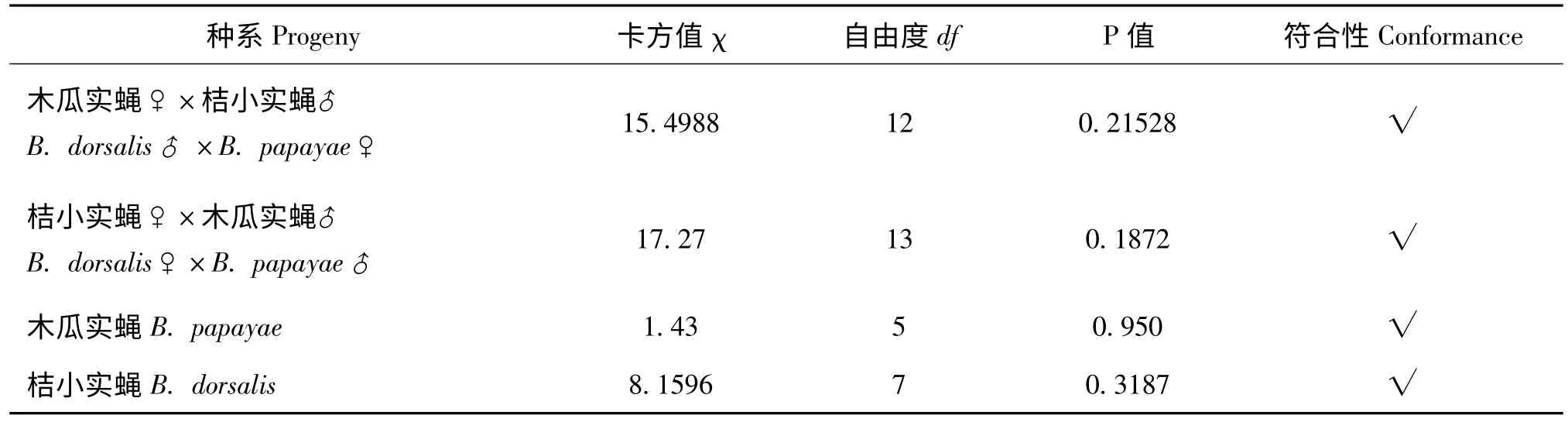
表2 桔小实蝇和木瓜实蝇及其杂交第33 代产卵管长度频次分布分布正态性检验Table 2 Normal distribution tests for length of aculeus of hybrid progeny between Bactrocera dorsalis s.s.and B.papayae after reproducing thirty-three generations
3 结论与讨论
对桔小实蝇和木瓜实蝇杂交代经过33 代的连续观测结果表明,两种实蝇杂交后可正常发育繁殖(种群数量特征另文发表),不存在严格的生殖隔离现象。杂交后代的产卵管长度表现为数量遗传特征,即后代的总体长度总是介于两亲本之间,且不同杂交种系之间(如亲本,杂交和回交代之间)的产卵管长度具有显著差异性,各种系内产卵管长度的频次分布也均表现一致。
传统分类中以产卵长度作为这两种实蝇分类鉴别依据的前提是:两者具有生殖隔离或没有经过基因交流。但从本研究观测结果看,无论是亲本还是杂交种尽管经过33 代的隔离繁殖(即保持种系内自交),其产卵管长度仍然保持其自身种系的特异性,但是,一旦种(系)间出现杂交,这种特异性会随着杂交作用的产生而发生改变,表现为随着杂交后代与亲本(或其他种系间)不断回交,其后代的产卵长度会因数量遗传特征与亲本的差异越来越小,其种(或种群)间的形态区别特征也愈来愈细微。
另外,本研究结果也从另一个侧面验证了桔小实蝇和木瓜实蝇为新近进化的种类,两者在东南亚地区由于地理隔离的产生而发生分化(Matthew et al.,2012),如产卵管长度的差异性,但这种分化还未足以进化成生殖隔离的独立种,两者的分化水平应当属于不同的地理种群。只要两种实蝇存在地理隔离,其产卵管长度的差异性会始终保持,而当地理隔离一旦消除或因人为作用使得两种实蝇相遇杂交,则以产卵管长度作为种间的区别特征将随之改变,这就意味着产卵管长度不能作为这两种实蝇固定的鉴别特征。
References)
Allwood AJ,Chinajariyawong A,Drew RAI,et al.Host plant records for fruit flies (Diptera:Tephritidae)in South East Asia[J].Raffles Bull.Zool.Suppl.,1999,7:1-92.
Clarke AR,Armstrong KF,Carmichael AE,et al.Invasive phytophagous pests arising through a recent tropical evolutionary radiation:The Bactrocera dorsalis complex of fruit flies[J].Annu.Rev.Entomol.,2005,50:293-319.
De Queiroz K.Ernst Mayr and the modern concept of species[J].Proc.Natl.Acad.Sci.USA,2005,102 (Suppl.1):6600-6607.
Drew RAI,Hancock DL,The Bactrocera dorsalis complex of fruit flies(Diptera:Tephritidae:Dacinae) in Asia [J].Bulletin of Entomological Research,1994,84:2 (i-iii):1-68.
Hardy DE.Taxonomy and distribution of the oriental fruit fly and related species (Tephritidae-Diptera)[J].Proc.Hawaii.Entomol.Soc.,1969,20:395-428.
Matthew N,Krosch MK,Schutze KF,et al.Piecing together an integrative taxonomic puzzle:Microsatellite,wing shape and aedeagus length analyses of Bactrocera dorsalis s.l.(Diptera:Tephritidae)find no evidence of multiple lineages in a proposed contact zone along the Thai/Malay Peninsula [J].Systematic Entomology,2012,38:2-13.
Schutze MK,Jessup AQ,UL-Haq I,et al.Mating compatibility among four pest members of the Bactrocera dorsalis fruit fly species complex(Diptera:Tephritidae) [J].J.Econ.Entomol.,2013,106 (2):695-707.
Schutze MK,Krosch MN,Armstrong KF,et al.Population structure of Bactrocera dorsalis s.s.,B.papayae and B.philippinensis(Diptera:Tephritidae)in southeast Asia:Evidence for a single species hypothesis using mitochondrial DNA and wingshape data[J].BMC Evolutionary Biology,2012,12:130.doi:10.1186/1471-2148-12-130.
