杨尺蠖蛹形态观察及球孢白僵菌菌株对其致病力的比较
2015-12-03姚红青谢映平王二虎
姚红青,谢映平*,王二虎
(1.山西大学生命科学学院,太原 030006;2.洪洞县林业局森林保护站,山西洪洞 041600)
杨尺蠖Apocheima cinerarius Erschoff 又名沙枣尺蠖、榆尺蠖、春尺蠖,在分类上属鳞翅目Lepidoptera 尺蛾科Geometridae,是一种暴食性的林木食叶害虫,危害林木有杨、榆、沙枣、沙柳、胡杨、桑、梨、苹果等。杨尺蠖幼虫的食量很大,虫口密度大时可在短时间内将成片的林木叶片全部吃光,导致树木长势衰弱或大面积死亡。该虫在国外分布于中亚地区,国内广泛分布于新疆、宁夏、甘肃、内蒙古、山西、河南、山东等省份(张执中,1997;马建军等,2010;闫祺等,2011)。近年来,该虫在山西临汾地区连续暴发,使大面积的杨树丰产林在5、6月份叶片被全部吃光,危害十分严重。
以往对杨尺蠖的防治主要在幼虫期采用树冠喷洒化学农药的方法,由于树冠高大,喷药困难且污染严重。在生物防治方面有利用飞机低容量喷雾苏云金杆菌(Bt)制剂和杨尺蠖核型多角体病毒(AciNPV)制剂进行杨尺蠖防治(杨升炯,1985;张永安等,2002)。但是,Bt 的芽孢伴晶体易被紫外线杀死,加上空气干燥,飞机喷洒出的雾化液滴极易在短时间内挥发而使Bt 失去附着力与致病力。AciNPV 制剂生产中的技术瓶颈问题是昆虫病毒的繁殖需要用寄主昆虫的活体幼虫,而杨尺蠖一年一代的特性极大限制了AciNPV 的室内连续生产和林间大面积应用(曲良建等,2011)。
白僵菌是重要的昆虫病原真菌,在我国已用于松毛虫等多种森林害虫的生物防治(蒲蛰龙和李增智,1996;Li,2001;汤陈生,2004),取得了较好的效果。但是,在野外应用白僵菌制剂存在的主要问题是紫外线照射、空气温度和湿度无法控制会降低白僵菌孢子活力和对害虫的感染力。在林间应用时,由于树冠高大,所以也需要更先进的喷雾和喷粉工具。为了避免这些弊端,本文结合杨尺蠖幼虫老熟后下树入土化蛹,蛹在土壤中要经历漫长的越夏和越冬的生物学习性,选择杨尺蠖的蛹期作为用白僵菌防治的时期,在山西省临汾地区的洪洞县杨树林布点采样,用球孢白僵菌的3个菌株进行染菌试验,检查杨尺蠖蛹的染病症状和死亡率,为在树冠下表土层中施用白僵菌粉剂,进行生物防治提供依据。
1 材料与方法
1.1 供试昆虫
供试昆虫的调查与采集:杨尺蠖在临汾地区洪洞县的杨树丰产林已连续发生数年,本研究观察和采样林地选在洪洞县大槐树镇(117.7E,36.3N),杨树为15年生,树高4.9-6.3 m,胸径15-20 cm,株行距2 m×2 m,从2013年3月至2014年8月,在林内定期观察杨尺蠖各虫态出现的始、盛、末期,并对越夏虫蛹的数量进行了调查。调查样树100 株,计算虫口密度。根据杨尺蠖化蛹和蛹的发育进度,确定在蛹期的前、中和后期即分别在5月(化蛹后5 d)、7月(化蛹后65 d)、10月(化蛹后165 d)采样。围绕树根基部,翻土,在表土层中采集虫蛹,带回实验室备用。
蛹的形态观察:将采集的杨尺蠖蛹在体视显微镜(OLYMPUS SZ-ST)下观察、测量和拍照,拍照用显微镜数码相机OLYMPUS C5050Z。虫蛹解剖和内部结构观察是将虫体用昆虫针固定于蜡盘上,在体视显微镜下进行。蛹的形态特征名称采用徐玉芬(1959)的术语。
1.2 菌种
菌种来源:本研究选用球孢白僵菌的三个菌株作为试验菌种,分别为TST05 菌株、FDB01 菌株和SYNO1 菌株。TST05 菌株是本实验室从采自山西省襄汾县桃小食心虫Carposina niponensis Walsingham 越冬幼虫的自然染病虫体上分离的病原菌,经过一系列试验已经证明其是桃小食心虫的优良致病菌(熊琦,2013),并于2011年1月保存于中国普通微生物菌种保藏管理中心,保藏号为CGMCC 4526;FDB01 菌株是由本实验室从采自吉林省伊通县红松林帕克阿扁叶蜂Acantholyda parki Shinohara & Byun 入土老熟幼虫自然染病虫体上分离的病原菌(Liu et al.,2012);SYN01 菌株购买于中国普通微生物菌种保藏管理中心,保藏号为CGMCC 3.4428,寄主为松毛虫Dendrolimus kikuchii Matsumura。上述三个菌株均用PDA 培养基连续培养,保存备用。
菌株分生孢子悬浮液的制备:将3 株球孢白僵菌分别接种于PDA 培养基试管中,在25℃±1℃,RH75%±5%,光周期15L∶9D 恒温箱中培养7 d,然后用0.5% 吐温80 溶液洗脱分生孢子,在显微镜下以血球计数板计数,检测分生孢子浓度,将其配置成1×108孢子/mL 浓度的悬浮液,供杨尺蠖蛹染菌试验使用。
1.3 虫体染菌
采用浸虫法进行染菌试验。将虫体大小基本一致的杨尺蠖蛹在浓度为1×108孢子/mL 的孢子悬浮液中浸10 s 后取出,每组处理试虫30 头,放入养虫盒中。空白对照组用同样方法浸0.5%吐温80 无菌水。各处理重复3 次。染菌处理后,处理组与空白组同时置入恒温培养箱内培养,条件为25℃±1℃,RH75%±5%,光周期15L∶9D。每天观察感染情况,并记录死亡虫数,持续观察8 d。
1.4 数据分析
试验数据的统计分析采用SPSS 13.0 软件处理,线性回归分析采用Excel 2003 软件处理。
2 结果与分析
2.1 杨尺蠖入土化蛹的生物学习性
据观察,杨尺蠖在山西临汾地区一年发生一代,2月15日前后越冬蛹开始羽化,成虫从树冠下的表土层钻出,沿树干爬行上树。雌雄成虫在地面或树干上交尾,交尾后雌成虫在树干上的裂皮缝隙产卵。3月20日前后卵开始孵化,3月25日前后部分初孵幼虫已经爬行到树冠上,开始取食叶片。幼虫历经5 龄,5月初老熟幼虫开始下树,5月10日进入下树末期。下树的老熟幼虫在树冠下钻入表土层,随即开始衔土粒,并吐丝、分泌粘液,将土粒黏在一起,逐步构建成一椭圆形、内壁光滑的蛹室,蛹室长度13.4-17.6 mm,粗度4.2-6.5 mm(图1)。约70%的蛹室分布于树干周围表土层15 cm-30 cm 深度范围内,最深可达50 cm。虫口密度最高达到150 头/株。

图1 蛹及蛹室Fig.1 Pupae and pupa cell
2.2 杨尺蠖蛹的外部形态和内部解剖特征
杨尺蠖蛹的外部形态:杨尺蠖的蛹属于被蛹(图2A 和2B),长13.0-17.0 mm,宽4.0-6.0 mm,分为头、胸、腹三部分,前期虫体颜色黄褐色,后期颜色变深,为灰褐色。头部:具复眼(Om)1 对。触角(An)1 对,栉状,紧贴体壁,伸到腹部第4 腹节后缘,末端与后足(Hl)末端相齐。口器的上颚发达,下颚(Mx)较长,末端止于中足末端稍前方,下唇须小,五角形;胸部:胸部分前、中、后胸3 节,中胸最大。中胸气门(Me)呈圆形突起。胸足3 对,紧贴体壁。前足末端伸长于下颚末端之前,中足末端超过下颚末端,后足(Hl)略显露。翅芽(Wb)发达;腹部:腹部具有许多刻点,10 节,第2-8 节各具1 对气门,气门椭圆形。第5、6、7 节可以扭动。雄蛹生殖孔位于第9 腹节,裂缝状,两侧呈半圆形瘤状突起。雌蛹生殖孔位于第8 腹节和第9 腹节之间,也呈裂缝状,周围内陷。肛门位于第10 节腹面中央,雄蛹肛门两侧有隆起,雌蛹则无。蛹的腹部末端具1个臀棘(Cr),分叉。
蛹的内部解剖特征:5月份虫蛹处于发育的前期,体色鲜亮,黄褐色,复眼与体色同。解剖开蛹的体壁,可见体内充满淡黄色脂肪体(Fb),直肠囊(Rc)位于中央,储存化蛹期间的代谢产物,充满红色液体。中肠和马氏管结构并未分化(图2C)。7月份虫蛹已经发育到中期,体色没有显著变化,但复眼的颜色变黑。剖开蛹的体壁,发现内部已发育出成虫,其表皮透明,触角,胸足和翅(Wg)均已分化,(图2E)。将该成虫解剖,虫体内的脂肪体(Fb)较多,淡黄色。中肠(Mg)呈扁椭圆形。观察到回肠(Ⅱ)为细管状,盘曲在腹腔内。消化道末端有一个大的直肠囊(Rc),储存蛹期的代谢产物呈砖红色(图2F)。马氏管(Mt)为淡黄色。此期雌虫体内还未发育出卵。
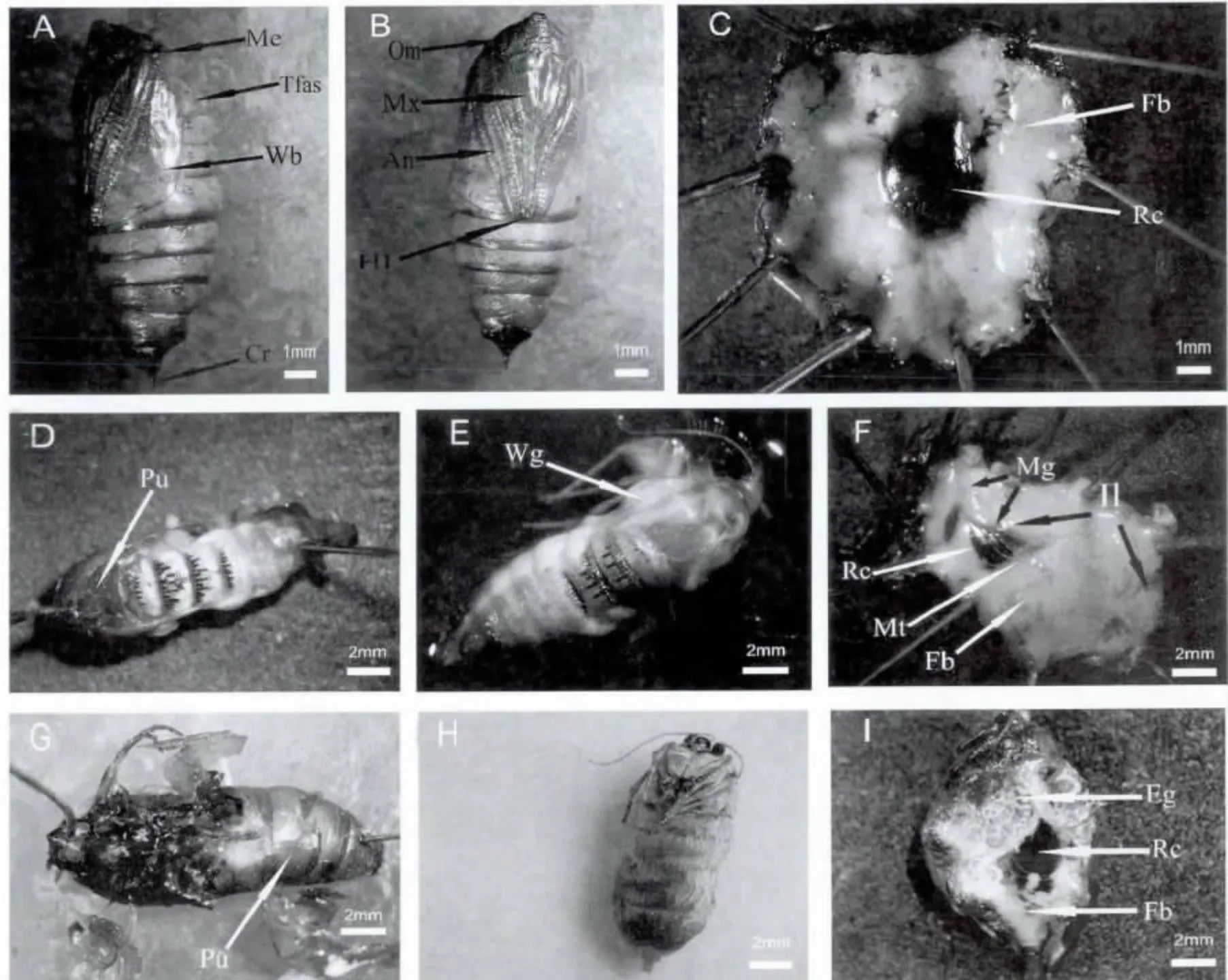
图2 杨尺蠖蛹外部形态及内部结构Fig.2 Morphological characteristics and inner structures of Apocheima cinerarius pupae
10月份采集的虫蛹已经发育为后期,蛹体颜色变深,呈灰褐色。解剖开蛹的体壁,可以看到内部发育的成虫的表皮已经变黑。触角,胸足和翅均变黑(图2H)。将该虫体解剖,看到腹部脂肪体(Fb)较多,中肠扁平呈椭圆形,回肠相对于中期蛹较短,消化道末端也有一个大的直肠囊(Rc),马氏管乳白色。雌蛹体内有大量的卵(Eg)(图2I)。
2.5 球孢白僵菌对杨尺蠖蛹的致病特点
球孢白僵菌TST05、FDB01、SYN01 均能侵染杨尺蠖蛹。染病的杨尺蠖蛹症状表现相似,感染初期表现为,身体开始变黑,受到外界刺激,腹部体节活动减缓。将死亡的虫体继续保湿培养,发现虫体表面长出白色菌丝(图3)。

图3 前期蛹被球孢白僵菌TST05 感染的症状Fig.3 Symptoms ofpupae in early stage infected by Beauveria bassiana TST05
2.6 不同时期三株球孢白僵菌对杨尺蠖蛹的致死率比较
杨尺蠖蛹接种3 株球孢白僵菌后,逐天检查死亡率,结果显示(图4),这3 株球孢白僵菌菌株对处在前期、中期、后期杨尺蠖蛹均有一定的致病力,蛹的校正死亡率随着感染时间的增加而上升。三个时期,虫体从第1 天出现了死亡。对前期蛹(4A),1-3 d,三菌株对杨尺蠖蛹的校正死亡率相比,FDB01 菌株>TST05 菌株>SYN01 菌株。3 d 后,TST05 菌株对杨尺蠖蛹的校正死亡率最高,FDB01 菌株次之,SYN01 菌株最低。到此后第8 天,TST05 菌株对杨尺蠖蛹的校正死亡率一直保持最高。在中期(4B),1-2 d,TST05 菌株对杨尺蠖蛹的校正死亡率最高,FDB01 菌株次之,SYN01 菌株最低。2 d 后,FDB01 菌株的校正死亡率略高于TST05 菌株,SYN01 菌株还是最低。从第4 天以后,TST05 菌株对杨尺蠖蛹的校正死亡率最高。在后期(4C),前4 d,三株菌对杨尺蠖蛹的校正死亡率相比,FDB01 菌株>TST05 菌株>SYN01 菌株。4 d 后TST05 菌株对杨尺蠖蛹的校正死亡率最高,并且到此后第8 天一直保持最高的趋势。

图4 不同时期TST05、FDB01 和SYN01 菌株对杨尺蠖蛹的校正死亡率Fig.4 Corrected mortalities of Apocheima cinerarius pupae after inoculated with TST05,FDB01 and SYN01,respectively
死亡率和LT50都可以反映出菌株对目标害虫的致病力,死亡率越大,LT50越小,说明致病力越强。表1 显示出第8 天不同时期三株菌对杨尺蠖蛹的校正死亡率和致死中时LT50。前期,TST05 菌株对杨尺蠖蛹的校正死亡率为71.05%±1.56%,LT50为4.76 d,各菌株间差异显著(P<0.05)。死亡率最高,LT50最小,对杨尺蠖蛹的致病力最高。中期,TST05 菌株对杨尺蠖蛹的校正死亡率为70.73%±5.23%,LT50为4.91 d,与其他菌株差异显著(P<0.05)。死亡率最高,LT50最小,对杨尺蠖蛹的致病力最高。后期,TST05 菌株对杨尺蠖蛹的死亡率最高,为61.18%±2.12%,与其他菌株差异显著(P<0.05)。LT50最小,为5.60d,对杨尺蠖蛹的致病力最高。同一菌株不同时期的校正死亡率和LT50不同。TST05 菌株对前期蛹和中期蛹的致死率较高,在70%以上,对后期蛹的致死率偏低,各时期间差异显著(P<0.05)。前期和中期LT50低于5 d,后期LT50为5.6 d。说明TST05 菌株对前期蛹和中期蛹的致病力较高,对后期蛹的致病力偏低。FDB01 菌株对前期和中期蛹的致死率均高于后期,但各时期间差异不显著(P>0.05)。LT50均在5 d 以上。SYN01 菌株对中期蛹的致死率最高,后期蛹次之,前期蛹最低,各时期间差异不显著(P>0.05)。LT50大于6 d。
综上所述,TST05 对杨尺蠖蛹致病力最强的菌株,而前期和中期都可以作为防治的最佳时期。
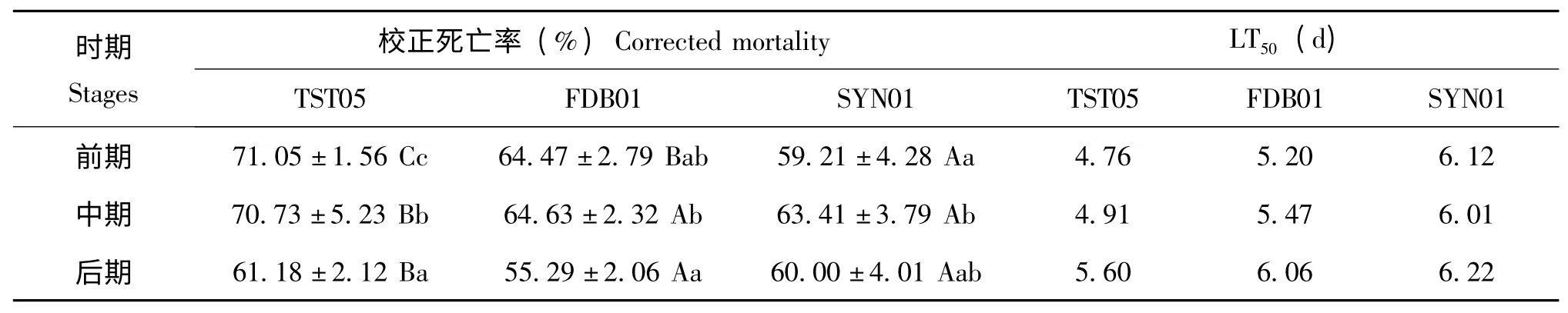
表1 不同菌株对杨尺蠖蛹的致死率和致死中时(LT50)Table 1 Corrected mortalities and LT50of Apocheima cinerarius pupae infected by the three strains of Beauveria bassiana
3 结论与讨论
杨尺蠖是一种暴食性食叶害虫,严重发生时可将杨树叶片吃光,影响树木生长甚至导致树木大面积死亡。目前对杨尺蠖的防治主要在幼虫期,用化学农药、Bt 和核型多角病毒进行防治。化学防治污染严重,Bt 和核型多角病毒的大面积防治需要飞机作业。此外,林间作业除了喷洒不均外,细菌和病毒都避免不了受到多种不稳定因素的干扰。比如:空气温湿度、紫外照射以及雨水冲刷。这些因素都不能使微生物杀虫剂持续发挥作用。
为了避免化学防治的弊端、节约生产成本和提高环境的可控性,可以选择在害虫下树入土期间进行防治。白僵菌在防治桃小食心虫等有地下生活史的害虫方面取得了很好的效果(朱永敏等,2012)。所以,本实验选用球孢白僵菌进行地下防治杨尺蠖。经调查,杨尺蠖的蛹期长达9个月之久,可以在杨尺蠖蛹期施用白僵菌菌粉,减少蛹的出土率。这样就充分发挥了地下防治的优势,既可以为球孢白僵菌孢子萌发提供有利的温度和湿度,又可以避免紫外线照射对孢子萌发的影响。本实验所用三株球孢白僵菌TST05、FDB01 和SYN01 的寄主分别为:桃小食心虫、帕克阿扁叶蜂和松毛虫。其中桃小食心虫和松毛虫与本文研究的杨尺蠖均属于鳞翅目昆虫。而帕克阿扁叶蜂与杨尺蠖的习性相似,在老熟幼虫下树入土后都会经历漫长蛹期。TST05 和FDB01 菌株对桃小食心虫和帕克阿扁叶蜂的致病力较强。所以选择这三个菌株进行试验。在染菌实验中,3 株球孢白僵菌菌株TST05、FDB01、SYN01 随时间的推移,对杨尺蠖蛹的致病性逐渐增强。用1×108孢子/mL浓度处理后,第8 天对杨尺蠖蛹的校正死亡率可达55.29%以上。这说明:球孢白僵菌在杨尺蠖蛹期的地下防治中有较大的发展潜力。我们筛选出对杨尺蠖致病力最高的菌株TST05,而且TST05 菌株对杨尺蠖前期蛹和中期蛹死亡率较高,后期蛹死亡率最低。所以可选择在前期和中期进行防治。
通过三个时期采样发现杨尺蠖蛹到后期颜色变深,说明蛹表皮逐渐几丁质化。通过多次解剖发现前期蛹刚进入发育期,剖开蛹壳,发现没有虫皮,各器官未发育,腹部脂肪体比中期蛹和后期蛹少,而脂肪体在细胞能量储存以及保护机体免受外界胁迫等方面发挥了重要作用(Thompson,2003)。所以前期蛹抗逆性最差,最容易染菌。而中期蛹刚好发育成成虫。剖开蛹壳后,发现虫体表皮透明,腹部脂肪体较多,所以中期蛹的抗逆性比前期蛹好。后期蛹在中期蛹的基础上进一步发育,蛹表皮颜色变深。剖开蛹壳发现,虫体变黑,腹部脂肪体较多,所以抗逆性最好,最不易染菌。刘玉文等(1999)研究表明蚕蛹在幼虫期的脂肪体在蛹期解离后重新形成,所以在前期脂肪体最少,中后期较多,这与本实验结果相似。球孢白僵菌不但能够寄生昆虫,而且在生态系统中寄主昆虫缺乏时,能够在不同的环境中营腐生生活(Studdert and Kaya,1990;Pireira et al.,1993)。这使得白僵菌可以在土壤中延续。本实验室熊琦(2013)在桃小食心虫病原真菌的研究结果中表明:TST05 菌株具有较强的宿存能力和较稳定的产孢水平,有利于起到持续控制害虫数量的作用。在起始浓度为106CFU/土样的土壤中,8个月后仍能达到104CFU。结合实际,我们可以在5月份老熟幼虫刚化蛹时用TST05 菌粉拌入土中,防治杨尺蠖蛹,也可在7、8月湿度较大的雨季施用球孢白僵菌菌粉。
References)
Li ZZ,Li CR,Huang B,et al.Discovery and demonstration of the teleomorph of Beauveria bassiana(Bals.)Vuill.,an important entomogenous fungus[J].Chinese Science Bulletin,2001,46:751-753.
Liu R,Xie YP,Zhao CS,et al.Beauveria bassiana isolated from pine sawfly and its pathogenicity[J].Mycosystema,2012,31(3):450-457.
Liu YW,Liu ZG.Morphology and anatomy of development of Chinese tussah incocoon heated stage[J].Journal of Shenyang Normal University,1999,3:38-43.[刘玉文,刘治国.中国柞蚕暖茧期蛹体发育的形态解剖学研究[J].沈阳师范学院报,1999,3:38-43]
Ma JJ,Yao H,Qi ZL.Emergence characteristics of overwintering Apocheima cinerarus and its larval spatial distribution pattern[J].Forest Pest and Disease,2010,29(1):10-12.[马建军,姚虹,齐志利.春尺蠖越冬蛹羽化特性及幼虫空间分布研究[J].中国森林病虫,2010,29(1):10-12]
Pireira RM,Alvas SB,Stimac JL.Growth of Beauveria bassiana in fire ant nest soil with amendments[J].J.Invertebr.Pathol.,1993,62:9-12.
Pu ZL,Li ZZ.InsetMycology[M].Anhui:Anhui Science &Technology Publishing House,1996,405-424.[蒲蛰龙,李增智.昆虫真菌学[M].安徽:安徽科学技术出版社,1996,405-424]
Qu JL,Wang YZ,Tao J,et al.Biological characteristics and multiplication of Apocheima cinerarius Erschoff nucleo polyhedro virus in lab[J].Forest Pest and Disease,2011,30(6):35-37.[曲建良,王玉珠,陶婧,等.春尺蠖NPV 生物学特性及室内增殖研究[J].中国森林病虫,2011,30(6):35-37]
Studdert JP,Kaya HK.Water potential,temperature and soil type on the formation of Beauveria bassiana soil colonies[J].J.Invertebr.Pathol.,1990,56:380-386.
Tang CS.Study on Excellent Strains and Controlling by Different Formulations of Beauveria bassiana in Forest[D].Fuzhou:Master Dissertation,Fujian Agrieulture and Forestry University,2004.[汤陈生.白僵菌优良菌株及林间使用剂型的研究[D].福州:福建农林大学硕士学位论文,2004]
Thompson NS.Trehalose-the insect blood sugar[J].Advance Insect Physiology.2003,31:203-285.
Xiong Q.Study on Beauveria bassiana strain TST05—An entomopathogenic fungus for Carposina sasakii[D].Taiyuan:PhD Dissertation,Shanxi University,2013.[熊琦.桃小食心虫病原真菌—TST05菌株的研究[D].太原:山西大学博士学位论文,2013]
Xu YF.Identification of main pest of Lepidoptera[J].Acta Entomol.Sin.,1959,9(5):395-416.[徐玉芬.鳞翅目主要害虫蛹的鉴别[J].昆虫学报,1959,9(5):395-416]
Yan Q,Hou XL,Ma JX,et al.Extraction and GC-MS identification of active components of sex pheromone from Apocheima cinerarius Erschoff[J].Acta Entomol.Sin.,2011,54(3):368-372.[闫祺,侯雪玲,马纪萱,等.春尺蠖性信息素活性成分的提取和GC-MS 鉴定[J].昆虫学报,2011,54(3):368-372]
Yang SJ.Low-volume aerial spray of Bt for the control of Apocheima cinerarius[J].Bulletin of Biological Control,1985,4:19.[杨升炯.飞机低容量喷雾Bt 乳剂防治杨尺蠖[J].生物防治通报,1985,4:19]
Zhang YA,Wang GC,Xu BM,et al.Report of controlling Apocheima cinerarius by AciNPV virus in Hetian region of Xinjiang[J].Forestry of Xinjiang,2002,2:20-30.[张永安,王贵成,徐保泯,等.新疆和田地区利用AciNPV 病毒防治春尺蠖试验报告[J].新疆林业,2002,2:20-30]
Zhang ZZ.Forest Entomology[M].Beijing:China Forestry Publishing House,1997,232-233.[张执中.森林昆虫学[M].北京:中国林业出版社,1997,232-233]
Zhu YM,Xue JL,Xie YP,et al.Virulence of the five strains of entomopathogenic fungi infected on the larvae of Carposina sasakii[J].Acta Phytophylacica.Sin.,2012,39(6):549-555.[朱永敏,薛皎亮,谢映平,等.五株病原真菌对桃小食心虫的致病力[J].植物保护学报,2012,39(6):549-555]
