科尔沁沙质割草地植物群落种间联结研究*
2015-12-02李晓兰相吉山秦立金赵雪梅
李晓兰,相吉山,赵 岩,秦立金,赵雪梅
(赤峰学院)
0 引言
种间联结是指不同物种在空间分布上的相互联结性,通常是由于群落生境的差异影响了物种的分布而引起的[1-2].种间联结是生态群落的重要特征之一,是群落形成和演化的基础,是重要的数量和结构指标,是种群间相互作用的一种表现形式,也是群落分类的依据.对不同种个体在空间联结程度的客观测定在研究群落水平格局的形成、种群进化和群落演替动态方面具有重大的意义[3].根据群落内共同出现的种对生境选择和要求上的异同以及相互间的吸引或排斥状况,可将种间关系分成正相关、负相关或不相关三种类型[4].
测定种间联结通常用样方抽样调查两个种存在或不存在的二元数据,排成2×2联列表,再将联列表内的数据代入有关数学公式进行计算,测定种间的联结程度.在国外Fisher(1943)[5]、Krglov(1968)[6]和 Yate 等(1972)[7]运用 χ2检验的方法测定了植物群落中种间的联结关系.Schluter(1984)[8]改进了前人的方法,提出了反映多种间关联的方差比值法,为测定多种间的复合相关性提供了新的途径.
我国关于植物种间联结性研究始于上世纪70 年代末,王伯荪、彭少麟[9-10]、李育中[11]等学者在种间相关测定取样技术、种间关联的理论和应用实践等方面作了大量的开创性工作,使我国在植物种间联结性的研究方面得到进一步发展.该文主要以科尔沁草地植被调查为基础,分析各物种之间的相关关系,进而为研究科尔沁沙地草地植物群落的结构、特性和演替提供依据.
1 研究区自然概况
研究地点位于科尔沁沙地西部,以多年围栏封育的沙质草地植物群落为研究对象,其地理位置位于东经 119°39',北纬 43°02',平均海拔494.6 m.该地区属于温带大陆性半干旱气候类型,平均年降雨量311 mm左右,主要集中在5~9月,占全年降水量的 70.1%;年蒸发量约2500 mm,年均温度为5.8 ~6.4 ℃;≥10℃的积温为3000~3200℃ ,无霜期140~160 d.
2 研究方法
2.1 取样方法
在围栏封育草地内随机设置1 m×1 m的样方160个.在每个样方内记录植物的种名、盖度、多度、高度、生活型等指标.
2.2 测量方法
根据样地调查资料,对调查数据进行整理,剔除盖度和频度小于5%的物种,建立2×2联列表,分别计算 a,b,c,d 值.其中,a 为两个种都出现的样方数;b为A种出现而B种不出现的样方数;c为A种不出现而B种出现的样方数;d为两个种均不出现的样方数.
(1)多物种间的总体关联性检验
Schluter(1984)[8]提出基于出现 -不出现数据的方差比率进行检验,其计算公式为:
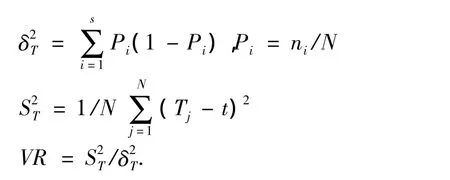
式中S为总物种数,N为总样方数,Tj为样方j内出现的研究物种的总数,ni为物种i出现的样方数,t为样方中种的平均数.
VR>1表示物种间表现出正的关联,VR<1表示物种间存在负的关联,VR=1表示复合种间关联性是独立的.值得注意的是在具体分析时,种间的正负关联是可以抵消的.
采用统计量W(=N×VR)来检验VR值偏移1的显著程度.例如种间不显著关联,则W落入由下面χ2分布给出的界限内的概率有90%,
(2)成对物种间联结性检验 -χ2检验
由于取样的不连续性,因此,非连续性数据的χ2值用Yates的连续校正公式计算:

式中N为样方数,其他意义同前.
(3)种间联结系数 AC(association coefficient)
关联系数AC在[-1,+1]之间,其最大值为+1,也就是两个物种共同出现的最大可能性,最小的可能性是 -1.计算关联系数时常采用下列公式[9-10,12]:
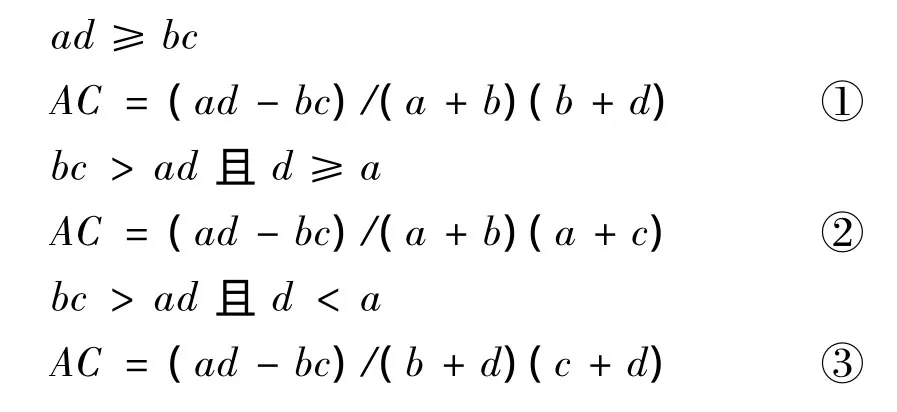
(4)共同出现百分率PC(percentage cooccurrence)
共同出现百分率PC是用来测度物种间正联结程度的,其计算公式为:
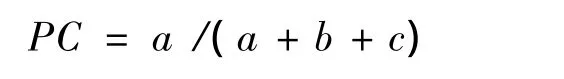
PC的值域为[0,1],其值域越趋近于1,则表明物种间的正联结性越强[9,13,14].
3 结果与分析
3.1 种间的总体关联分析
对于调查样方中盖度和频度≥5%的物种间的整体关联进行分析,结果表明:群落内种间的总体关联性的方差均值VR=2.624,表现为多物种间存在正相关.检验相关性的统计量W=419.84,W 不落入之间,说明所有调查38个种群之间在总体上表现出显著的正相关.
种间的总体关联性反映了群落的稳定性[15].一般来说,进展演替的植物群落,群落结构及其种类组成将逐渐趋于完善和稳定,种间关系也逐渐趋向于正相关,以求得物种间的稳定共存[16-17].上述结果说明,科尔沁割草草地群落种物种间表现出显著的正联结性,反映了该群落处于较稳定的阶段.
3.2 种间联结的χ2检验
χ2检验能比较准确的表示种对间联结的显著程度[13,18](图 1).从图 1 可以看出,38 个物种两两组合构成703个种对,共有230个种对的种间关联达到显著水平;其中,正、负关联分别为124个和106个,正、负关联达到极显著水平(P<0.01)的种对分别为90个和34个.其余种对的关联性都不显著,占总种对的67.3%.
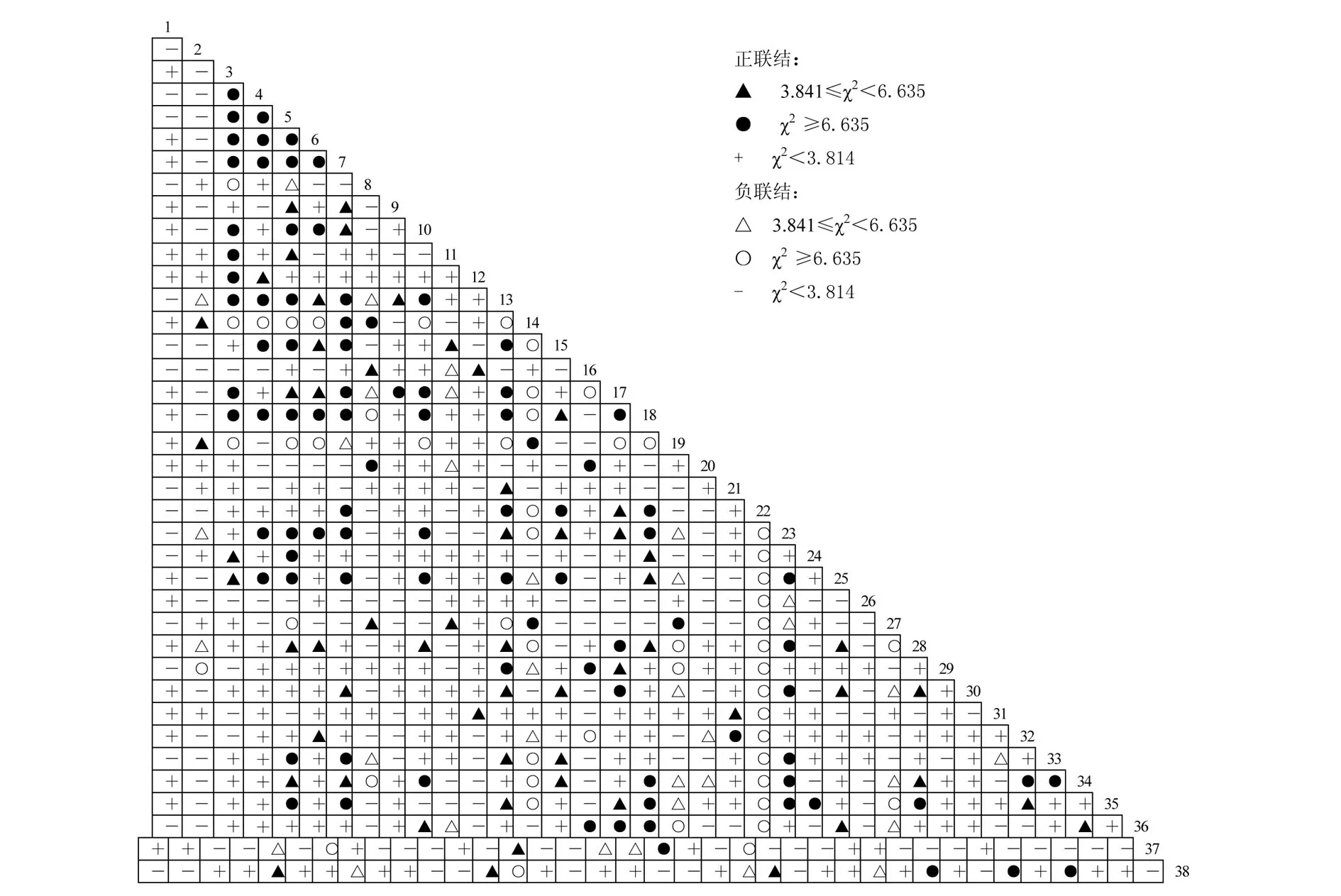
图1 38种主要种群间χ2值的半矩阵图
由于χ2检验只是确定了种间关联性显著与否,并没有对种间关联性的大小作出定量分析,有些种对经χ2检验不显著的种对实际上也存在着一定的关联性,因此,χ2检验法只能对种间关联作出定性的判别.而联结系数AC和共同出现百分率PC则能体现那些由χ2检验证明不显著的联结性的大小[13].
3.3 种间联结系数AC
由联结系数计算公式可知,AC的值域为[-1,1].AC的值越接近于1,表明2个物种共同出现和共同不出现的可能性越大;相反,AC的值越接近于 -1,表明2个物种单独出现的可能性越大.本围封割草地38个主要种群组成的703个种对间的联结系数详见图2.
从图2可知,703对个种对中67.3% 的种对经χ2检验后没有达到显著水平.联结系数AC≥0.6的种对有43对,仅占6%;AC <-0.6的种对有60对,占8.5%;-0.6≤AC < -0.2的种对有105对;-0.2≤ AC <0.2的种对有 287对;0.2≤AC <0.6的种对有208对.
3.4 共同出现的百分率PC
PC值是用来测定物种间联结程度强弱的指标,其值域[0,100%],PC值越接近于100%,则表明2物种间的正联结程度越高,2物种共同出现和不出现的可能性越大,2物种的生态习性和对环境的需求越一致.38种主要种群的703个种对间的PC值详见图3.
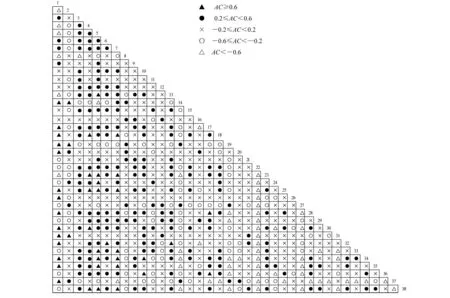
图2 38种主要种群间联结系数的半矩阵图
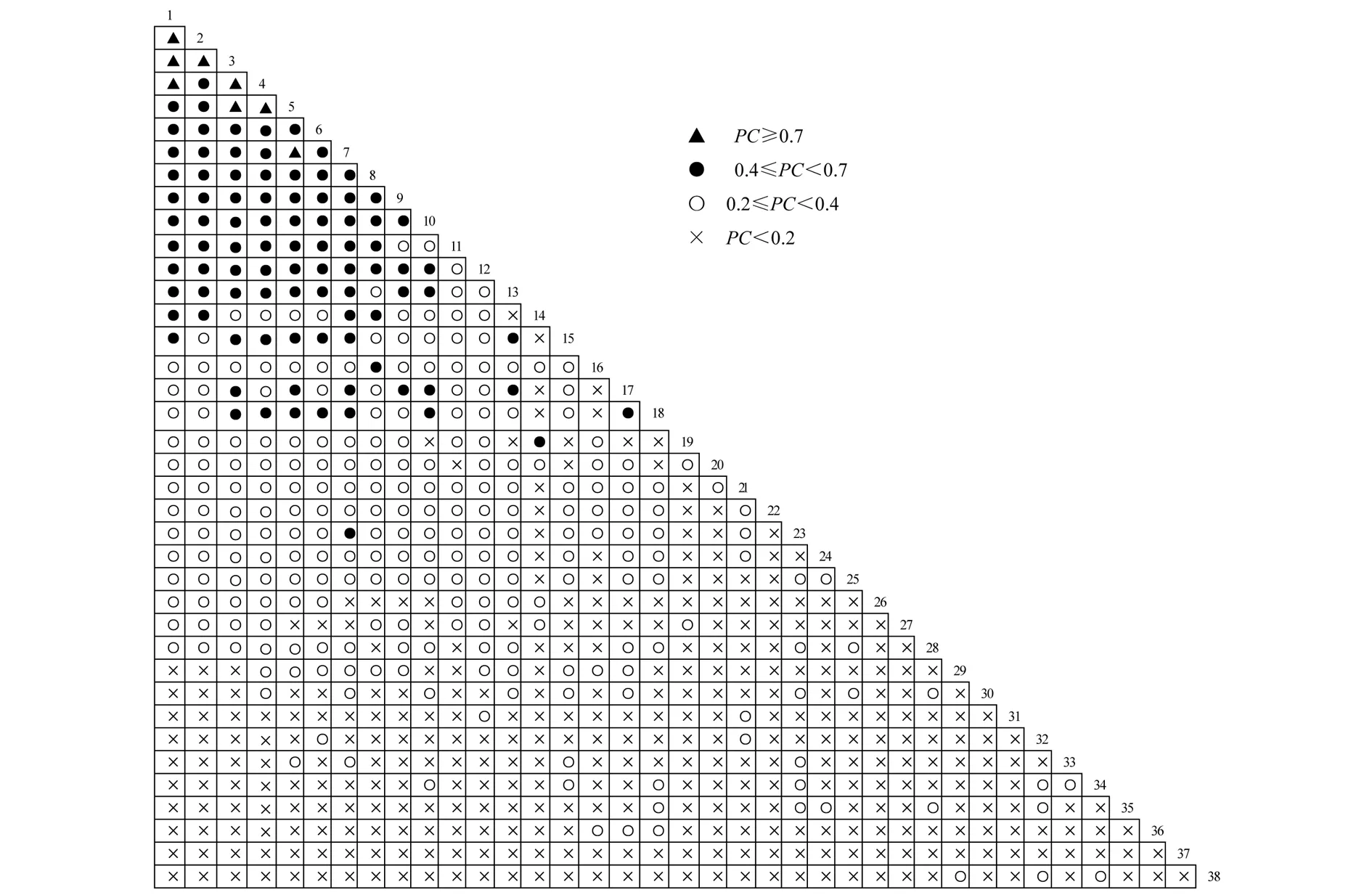
图3 38种主要种群间共同出现百分率的半矩阵图
由图3可知,PC<20% 的种对为358对,所占比例已达51%;20%≤PC<40%的种对为244对,占34.7%;40% ≤PC <70% 的种对为91对,占12.9%;而PC≥70% 的种对仅有8对,仅占1%.由此可见38种主要物种种间的联结性较弱.
4 结论与讨论
结果与分析已表明,χ2检验只是对种间联结性给出了一个概括性的结论,AC值和PC值虽能反映种间联结性的相对强弱,但就AC值而言,由于计算公式中分子为(ad-bc),当所研究群落中物种数目较多,而调查样方面积较小仅能体现较少的物种数(人为地减小了b值或c值)时,它一方面夸大了负联结性,另一方面则由于高d值(2个物种在调查样方中均不出现)导致AC值正值偏高.PC值由于忽视了d值的作用,无形中夸大了a、b、c值在联结测定中的作用(参见PC的计算公式).因此,将χ2检验、AC值和PC值三个指标结合起来分析,结果更为可靠.
科尔沁围封割草草地群落内38种主要草本种群构成了703个种对,经χ2检验、AC值和PC值的比较分析可知,有近一半的种对间的联结性不显著.经χ2检验种间联结关系显著、AC值和PC值相对较高的种对是:大油芒—掐不齐、狗尾草—野韭菜、狼毒—黄金菊、节节草—黄金菊、芦苇—野韭菜、洽草—黄金菊、狼毒—山莴苣、芦苇—轮叶沙参8个种对.表明这些种对的生态习性和对环境的需求较一致.
近些年来,科尔沁草地正处于不同程度的退化状态,所以为了实现可持续发展,应该对科尔沁地区的原有植被进行分析.根据该文的种间联结分析,揭示了植物群落种间关系,进而为植被恢复过程中的种群配置和植被管理提供重要的科学依据,对指导生产实践具有重要的意义.
[1] 王伯荪.植物种群学[M].广州:中山大学出版社,1989.
[2] Greig-Smith P.Quantitative Plant Ecology[M].3rd ed.Blackwell Scientific publications,1983.
[3] Cox G W.Laboratory Manual of General Ecology.8th ed Wm C Brown Co,Dubuque,2002.167-169.
[4] Hubalek Z.Coefficients of association and similarity basedon binary data:an evaluation.Biological Reviews,1982,57:669-689.
[5] Fisher R A,et al.The relation between the number of species and the number of individuals in a random sample of an animal population [J].J Animal Ecol,1943,12(1):42-58.
[6] Krglov V V.Species association in plankton[J].Oceanology,1968,8(2):243-251.
[7] Yates F E,et al.Integration of the whole organism-A foundation for a theoretical biology.Challenging biological problem:direction towards their solutions[M].Oxford Univ Press New York,1972.
[8] Schluter D.A variance test for detecting species associations,with some example applications.Ecology,1984,65:998-1005.
[9] 王伯荪、彭少麟.南亚热带常绿阔叶林种间联结测定技术研究.Ⅰ.种间联结测式的探讨与修正[J].植物生态学与地植物学丛刊,1985,9(4):274-285.
[10]彭少麟,王伯荪.南亚热带常绿阔叶林种间联结测定技术研究.Ⅱ.种间联结研究的取样技术[M].热带亚热带森林生态系统研究,1985.
[11]李育中.三种类型草地植物种间关联的测定与比较[J].生态学杂志,1991,10(6):6-10.
[12] Hurlbert S H.A coefficient of interspecific association.Ecology,1969,50:1-9.
[13]郭志华,卓正大,陈洁,等.庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究[J].植物生态学报,1997,21(5):424-432.
[14]王伯荪译.植物群落排序[M].北京:科学出版社,1986.
[15]周先叶,王伯荪,李鸣光.广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J].植物生态学报,2000,24(3):332-339.
[16]杨一川,庄平,黎系荣.峨眉山峨嵋栲、华木荷群落研究[J].植物生态学报,1994,18(2):105-120.
[17]杜道林,刘玉成,李睿.缙云山亚热带栲树林优势种群种间联结性研究[J].植物生态学报,1995,19(2):149-157.
[18]阳小成,钟章成.绵阳宽司河流域防护林优势种群间的联结性研究[J].西南师范大学学报:自然科学版,1994,19(3):290-297.
