吉林延龄草营养器官解剖结构和光合生理特性对环境适应性研究
2015-11-30刘继永王振兴
邵 财,刘继永,张 浩,王振兴
(中国农业科学院特产研究所,长春 130112)
吉林延龄草营养器官解剖结构和光合生理特性对环境适应性研究
邵 财,刘继永,张 浩,王振兴
(中国农业科学院特产研究所,长春 130112)
采用解剖显微观察和光合仪测定等方法研究吉林延龄草根、茎、叶营养器官的解剖结构及光合生理特性,揭示其形态解剖结构、光合特性与环境适应性的关系,为吉林延龄草保护及驯化栽培提供理论基础和科学依据。结果表明,吉林延龄草只具有初生构造的根、茎及维管组织中没有形成层,初生木质部导管数量少,输导组织不发达;叶片表皮无表皮毛,表皮细胞凸凹不平;叶肉细胞由海绵薄壁细胞组成,排列疏松,细胞之间存在较大间隙;叶片上表皮无气孔,气孔分布在叶下表面,气孔密度为18.69个·mm-2,气孔表现“外凸”特征。叶片叶绿素a/b比值为1.90;其具有较低光补偿点(LCP)和光饱和点(LSP),分别是1.23和415 μmol·m-·2s-1,在光饱和点可达最大净光合速率为4.44 μmol·m-·2s-1。研究表明,吉林延龄草在形态结构、光合生理特性上具有明显适应阴生潮湿环境特征,是典型的阴生植物。
吉林延龄草;解剖结构;光合特性;环境适应
网络出版时间2015-4-30 14:31:00 [URL]http://www.cnki.net/kcms/detail/23.1391.S.20150430.1431.006.html
邵财,刘继永,张浩,等.吉林延龄草营养器官解剖结构和光合生理特性对环境适应性研究[J].东北农业大学学报,2015,46(5): 32-37.
Shao Cai,Liu Jiyong,Zhang Hao,et al.Study on environment adaptation of vegetative organs anatomical structure and photosynthetic physiological characteristic ofTrillium kamtschaticumPall.ex Pursh[J].Journal of Northeast Agricultural University,2015,46(5):32-37.(in Chinese with English abstract)
吉林延龄草(Trillium kamtschaticum Pall.ex Pursh)为百合科延龄草属植物,分布于亚洲东北部地区,中国主要分布于吉林,生长于海拔500~1 400 m的林下、林缘潮湿处[1],根茎含有甾体皂甙、偏诺皂苷、延龄草皂苷、薯蓣皂苷、克里托皂苷、甾酮皂苷、倍半萜苷等多种有效成分[2-4],以干燥根茎入药,具有镇静、止痛、活血、止血功效,是治疗高血压、神经衰弱、眩晕头痛、腰腿疼痛、月经不调、外伤出血、跌打损伤等症的珍贵中药材[5]。
目前,由于森林过度采伐,严重破坏林下吉林延龄草生长环境,分布范围日趋缩小,加上根茎入药、过量挖掘等因素,致使其种群数量逐渐减少,如不进行合理保护利用,将会面临濒危风险。近年来,我国部分地区有小规模驯化栽培,虽能成活但生长不良。Nott等研究认为,植物营养器官形态解剖结构及光合生理特征最能体现环境因子的影响及植物对环境的适应[6-7],迄今为止,国内外对吉林延龄草的研究主要集中在化学成分、药理作用及其外部形态特征上。为此,本文对吉林延龄草营养器官根、茎、叶解剖结构进行观察,光合特性分析,深入探讨其与所处环境的关系,为吉林延龄草保护及驯化栽培提供理论基础和科学依据。
1 材料与方法
1.1 吉林延龄草形态解剖结构观察
吉林延龄草材料采自吉林左家镇,在取样区域内随机选取生长良好的植株,分别选取吉林延龄草根、茎、叶,采用FAA固定液[38%甲醛∶冰乙酸∶70%乙醇=1∶1∶18(体积比)]固定,采用石蜡切片法制片[8-9],翻红-固绿对染,Nikon Eclipse 80i光学显微镜观察并拍照。
采用无色指甲油印痕法对吉林延龄草成熟叶上、下表皮气孔分布进行观察,NIS-Elements AR3.2软件进行相关数据测量分析。
1.2 叶片叶绿素含量测定
2014年6月,随机选取3株长势良好、朝向一致的植株,每株选取大小基本一致叶片,采用张志良等方法测定叶绿素含量[10]。
1.3 光合作用-光响应的测定
2014年6月吉林延龄草生长旺盛期,选择晴天9:00~12:00测定净光合速率(Pn)对不同光合有效辐射(PAR)的响应。在取样区域内选取3株长势良好、朝向一致、叶片大小相似的植株,应用英国Hansatech公司生产的CIRAS-2型便携式光合测定系统对其叶片中部进行光合作用-光响应的测定。测定时使用大气CO2浓度,利用人工光源,光强设定800、700、600、500、400、300、200、100、0 μmol·m-2·s-1共9个梯度。每次测定时间均为180 s,仪器自动记录净光合速率(Pn),绘制光合作用光响应曲线(Pn-PAR曲线),参照张保玉方法利用二项式拟合方程求出吉林延龄草光合作用光饱和点(LSP)、光补偿点(LCP)和最大净光合速率(Pmax)[11]。
2 结果与分析
2.1 吉林延龄草形态解剖结构
吉林延龄草根形态及营养器官解剖结构见图版Ⅰ。
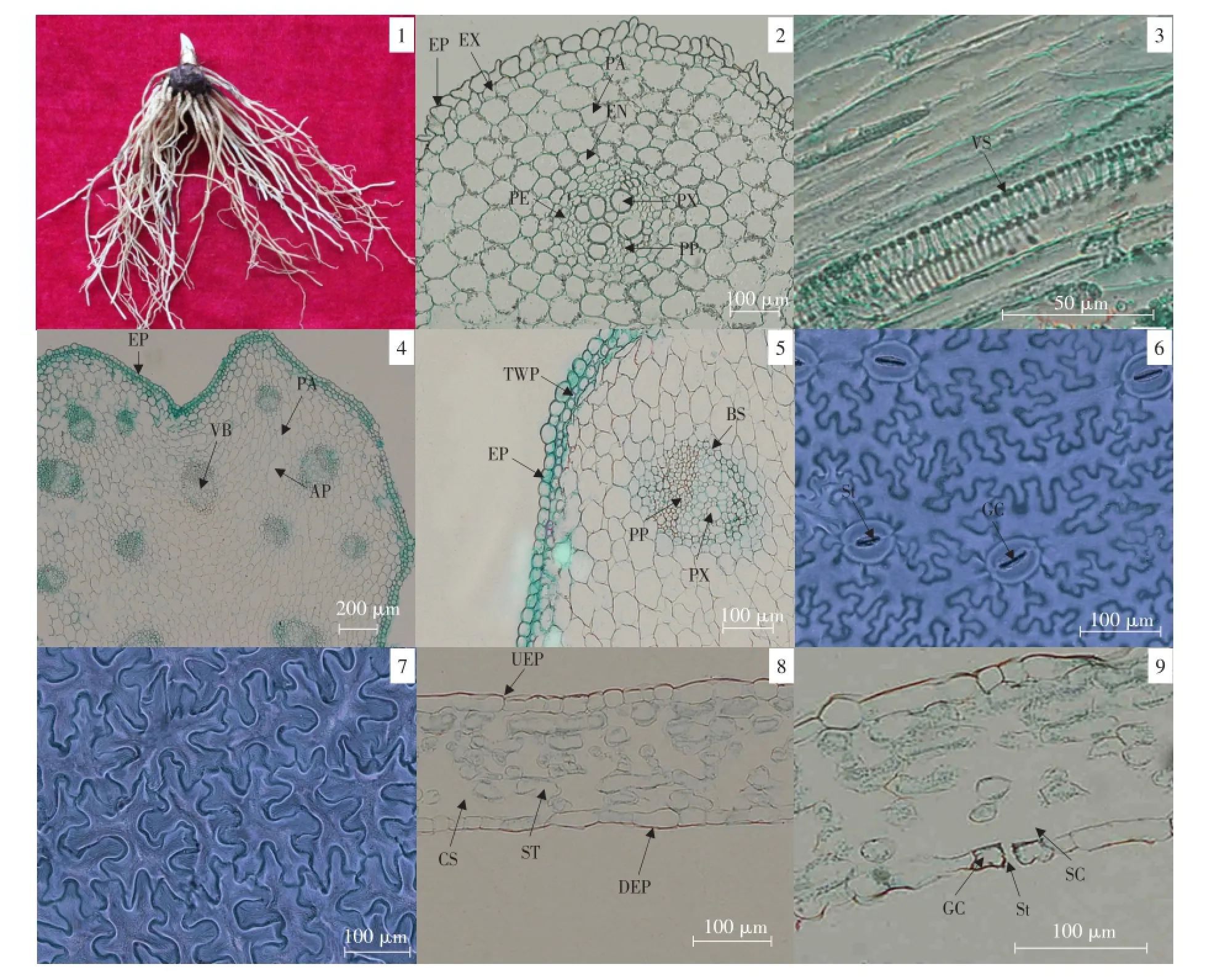
图版Ⅰ 吉林延龄草根形态及营养器官解剖结构PlateⅠ Root morphological and anatomical structure of nutritive organs of Trillium kamtschaticum
2.1.1 吉林延龄草根形态解剖结构
吉林延龄草根具有典型单子叶植物根的结构(见图版Ⅰ-1、2),包括表皮、皮层、中柱三部分,无次生结构。表皮细胞1层,细胞形状不规则、排列紧密。由根外部形态可知,其须根侧根较少,根尖成熟区未发现根毛。根横切面整体上呈现“车轮形”,即外皮层与内皮层的联系是靠多层薄壁细胞叠加起来的细胞群连接,皮层在整个横切面占有比例较大,有7层细胞,外皮层有1层细胞且细胞壁明显加厚,细胞排列紧密,无细胞间隙,成为连续的一层,形成根的保护层,内皮层未见明显凯氏带构造。中柱包括中柱鞘、初生木质部、初生韧皮部。初生木质部与初生韧皮部成辐射式排列,初生木质部包括原生木质部和后生木质部两部分,原生木质部在木质部的最外面,其中导管孔径较细,后生木质部束生达中柱中心,无髓结构,导管数量较少,排列紧密,孔径大小不一,后生木质部薄壁组织细胞体积较小,排列于导管周围;木质部具有四个木质部束,为四原型木质部;由根的纵切可知(见图版Ⅰ-3),木质部导管类型为螺纹导管。
2.1.2 吉林延龄草茎形态解剖结构
吉林延龄草茎初生结构由表皮、皮层、维管束三部分组成(见图版Ⅰ-4)。表皮细胞1层,细胞近似砖形,排列紧密,没有细胞间隙。表皮层之内有1层厚壁的薄壁组织(硬化薄壁组织),硬化薄壁组织之内为薄壁组织,皮层薄壁细胞体积较大,呈不规则形状,细胞间存在较小细胞间隙,细胞间分布有少量气道(通气组织);维管束分散分布在茎中,在茎的外周分布较密集,在茎的中心分布较疏松,维管束包含初生木质部和初生韧皮部两部分,没有形成层,但茎中维管束被一圈厚壁组织鞘(束鞘)包被,韧皮部细胞小,排列紧密,不同维管束中木质部导管数量不等,成环形排列,导管孔径大小有差异(见图版Ⅰ-5)。
2.1.3 吉林延龄草叶形态解剖结构
由图版Ⅰ-6、7可见,吉林延龄草上、下表皮细胞呈不规则形状,犬牙交错,排列紧密,无细胞间隙,上表皮无气孔,气孔仅分布在下表皮,保卫细胞肾形,气孔口长椭圆形;下表皮气孔密度为18.69个·mm-2。由叶横切面观察(见图版Ⅰ-8、9),上、下表皮均由单层细胞构成,上表皮有较薄的角质层,无表皮毛,表皮细胞凸凹不平;下表皮气孔略微凸起,气孔下有较大的气孔下室;上、下表皮之间的叶肉由海绵薄壁细胞组成,无明显栅栏组织分化,叶肉细胞形状不规则,细胞排列疏松,细胞间有较大间隙,形成叶肉中较为发达的通气组织。
2.2 吉林延龄草叶片光合生理特性
叶片中叶绿素含量是维持植物正常光合作用以及叶片色泽、彩化度的主要指标,与生境条件和植物种的特性相关。由表1可见,吉林延龄草叶片中叶绿素含量较高,其叶绿素a/b比值为1.90。光补偿点(LCP)是植物进行光合作用所需的最低光强,光强低于LCP,植物生长不能正常进行,光补偿点越小,表明植物利用弱光的能力越强;光饱和点(LSP)是植物光合作用最大时的光强,是植物利用强光能力的指标,光饱和点越高则表明植物适应强光能力越强。吉林延龄草光合作用-光响应曲线(Pn-PAR曲线)表明(见图1),其具有较低的LCP,仅为1.23 μmol·m-2·s-1,同时,具有较小的LSP,为415 μmol·m-2·s-1,在光饱和点可达最大净光合速率为4.44 μmol·m-2·s-1,由此可知,吉林延龄草利用弱光的能力较强,适应强光的能力较弱。

图1 吉林延龄草叶片净光合速率对不同光合有效辐射的响应Fig.1 Light response curves of the net photosynthetic rate (Pn)to the photosynthetically active radiation(PAR)of the leaves of the Trillium kamtschaticum
3 讨论与结论
3.1 解剖结构对环境的适应
生长在不同生态环境中的植物,表现出不同的形态结构,是植物对特定生境的演化适应[12]。吉林延龄草生于海拔500~1 400 m的林下、林边阴湿处。本研究发现,吉林延龄草具备阴生植物结构特征,体现出对阴生环境的适应性。
从吉林延龄草石蜡切片来看,吉林延龄草只具有初生构造的根、茎,维管组织中无形成层,不能够加粗,其根、茎无次生生长,初生木质部导管数量少,孔径较细,输导组织不发达。在这种情况下,根尖吸水能力不能超过输导组织的运输能力。因此,吉林延龄草不能生出太多侧根,根外部形态观察根尖成熟区没有根毛,减少水分吸收面积,这与阴生植物黄连研究报道一致[13]。
栅栏组织发达、表皮细胞排列平直是阳生叶典型特征,而海绵组织发达、表皮细胞凸透形则是叶片对于弱光、水分充足的一种适应特性[14-16]。本研究中吉林延龄草叶肉组织无明显栅栏组织,叶肉细胞仅由海绵薄壁细胞组成,排列疏松、胞间隙较大,可减少光量子投射损失,使植物更充分地利用有限光能进行光合作用,积累有机物质,提高阴生环境适应性[17]。此外,Ehlreinger等研究报道,表皮是控制叶片吸收光量子重要因素,叶片表皮附属物,如表皮毛、短柔毛等,可减少叶片对光量子吸收[18-19]。吉林延龄草生长在阴生环境中,无高光强照射,其叶片无表皮毛,可减少光量子损失,利于吸收更多光能,提高光合产量,适应林下弱光生长环境。
气孔是植物与外界环境进行气体交换重要通道,气孔密度关系到植物光合作用、呼吸作用和蒸腾作用等重要生理过程。王荣等研究发现,生长在高光强下的植物通常比低光强下的植物具有更高的气孔密度[15,17]。林植芳等对大量植物叶片的气孔分布研究后发现,生长迅速、光合能力较强的植物有较高的气孔上下表皮比,生长较缓慢,光合效率较低的阴生植物则在叶片上表面只有少数或没有气孔,与本研究结果一致[20]。有利于缓解在较高光强下由于具有较大叶肉表面积而造成CO2扩散受阻情况[21]。本研究吉林延龄草叶气孔下有较大气孔下室,可能是对叶片气孔少的适应,防止CO2扩散受阻;叶片上表皮无气孔分布、下表皮气孔数量低于阴生植物平均水平(40~100个·mm-2)[22]、气孔密度小(18.69个·mm-2)等结构特征,可减少蒸腾作用[23]。表明植株适宜生长在阴湿环境中,避免强光下由于维管组织不能为叶片提供充足水分及因气孔数量较少,无法达到正常蒸腾降温目的,造成叶片局部温度过高而干枯,甚至导致植株死亡;叶片下表皮气孔“外凸”特征适应潮湿阴生的环境[13]。
3.2 光合生理特性对环境的适应
叶绿素是植物光合作用吸收光能的主要光合色素,一般来说阳地植物叶片叶绿素a/b比值约为3∶1,而阴生植物叶绿素a/b比值约为2.3∶1[22],从本研究结果可知吉林延龄草叶片含有高叶绿素含量,但其叶绿素a/b比值仅为1.90,小于阴生植物比值标准,表明吉林延龄草叶片中叶绿素b相对含量较高,可更有效利用漫射光中较多蓝紫光,适应阴生环境生长。
光合-光响应参数提供植物对环境条件变化适应能力的光合特性信息,反映植物对光照强度适应能力,在一定程度上决定植物分布模式。植物LSP表示对光照强度需求量,其值越高表示对光强需求越大。植物LCP反映其对弱光适应能力,LCP值越小,植物利用弱光能力越强[16,24]。白伟岚等通过对50种植物光合特性曲线分析,指出光补偿点和光饱和点是评价植物耐阴能力指标,一般草本阴生植物光补偿点和光饱和点值域分别是1~5和400~850 μmol·m-2·s-1,草本阳生植物光补偿点和光饱和点值域分别是10~40和1 200~2 500 μmol·m-·2s-1[25]。本研究表明,吉林延龄草光补偿点和光饱和点值域分别是1.23和415 μmol·m-2·s-1,具有较低光补偿点和光饱和点,表明利用弱光能力较强,对光强需求量较小,与胡天印等研究结果一致[26]。
以上研究表明,从吉林延龄草形态、解剖结构特点均表现出其对阴生环境的适应;从叶绿素a/b比值和光合-光响应曲线分析,吉林延龄草是典型阴生植物。因此,吉林延龄草在原产地就地保护时,应注意调光,避免树木茂盛郁闭度过大或清林过度郁闭度过小,在引种驯化栽培过程中,进行适当遮阴处理,调节最适光强,达到迁地保护目的,实现资源可持续利用。
[1] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1978:97.
[2] Masateru O,Yoshihiro Y,Tsuyoshi I.Steroids from the under⁃ground parts of Trillium kamtschaticum[J].Chemical and Pharma⁃ceutical Bulletin,2003,51:1328-1331.
[3] Masateru O,Chika T,Fumie S.Two new steroid glycosides and a new sesquiterpenoid glycoside from the underground parts of Tril⁃lium kamtschaticum[J].Chemical and Pharmaceutical Bulletin, 2007,55:551-556.
[4] Masateru O,Fumie S,Syoko S.Three new steroid glycosides from the underground parts of Trillium kamtschaticum[J].Chemical& Pharmaceutical Bulletin,2007,55:1093-1096.
[5] 国家中医药管理局.中华本草[M].上海:上海科技出版社, 1999:174-175.
[6] Mott K A,Gibson A G,O'Leary J W.The adaptive significance of amphistomatic leaves[J].Plant Cell Environ,1982(9):455-460.
[7] 庞杰,张凤兰,郝丽珍,等.沙芥幼苗叶片解剖结构和光合作用对干旱胁迫的响应[J].生态环境学报,2013,22(4):575-581.
[8] 赵曼容,贾恢先.植物学实用技术[M].兰州:甘肃民族出版社, 1993:22-42.
[9] 李正理.植物组织制片学[M].北京:北京大学出版社,1996: 130-139.
[10] 张志良,瞿伟菁,李小芳.植物生理学实验指导[M].北京:高等教育出版社,2009:58-60.
[11] 张保玉.葡萄光合作用光响应曲线的研究[D].杨凌:西北农林科技大学,2009:22-28.
[12] 闫成才,刘晓娟,田青,等.藏荠营养器官解剖结构及其与环境的关系[J].西北植物学报,2013,33(5):946-951.
[13] 袁王俊,张维瑞,尚富德,等.黄连营养器官解剖结构与其阴生环境相关性研究[J].河南大学学报,2007,33(2):184-186.
[14] 贺金生,陈伟烈,王勋陵,等.高山栎叶的形态结构及其与生态环境的关系[J].植物生态学报,1994,18(3):219-227.
[15] 王荣,郭志华.不同光环境下枫香幼苗的叶片解剖结构[J].生态学杂志,2007,26(11):1719-1724.
[16] 陈模舜,柯世省.天台鹅耳枥叶片的解剖结构和光合特性对光照的适应[J].林业科学,2013,49(2):46-53.
[17] 张泽宏,吴小霞.5种蕨类植物叶片解剖结构及其对阴生环境的适应性研究[J].华中师范大学学报,2013,47(6):840-843
[18] Ehlreinger J,Bjorman O,Mooney H.A leaf pubescence effects on absorptance and photosynthesis in a desert shrub[J].Science, 1976,192:376-377.
[19] Eller B M,Willi P.The significance of leaf pubescence for the ab⁃sorption of global radiation by Tussilago farfara[J].Oecologia, 1981,49:366-370.
[20] 林植芳,李双顺,林桂珠.叶片气孔的分布与光合途径[J].植物学报,1986,28(4):387-395.
[21] 史刚荣,蔡庆生.白三叶叶片解剖可塑性及其对光强的响应[J].草地学报,2006,14(4):301-305.
[22] 王忠.植物生理学[M].北京:中国农业出版社,2002:66,131.
[23] 斯日古楞,王明玖,额尔敦花.虉草营养器官解剖结构与抗旱耐涝性及利用之间的关系[J].植物研究,2013,33(4):391-397.
[24] 柴胜丰,庄雪影,韦霄,等.光照强度对濒危植物毛瓣金花茶光合生理特性的影响[J].西北植物学报,2013,33(3):547-554.
[25] 白伟岚,任建武,高永伟,等.园林植物的耐荫性研究[J].林业科技通讯,1999,2(2):12-15.
[26] 胡天印,李娜,郭水良.珍稀植物延龄草濒危的形态解剖、光合和环境因素分析[J].科技通报,2007,23(4):508-518.
Study on environment adaptation of vegetative organs anatomical structure and photosynthetic physiological characteristic ofTrillium ka⁃mtschaticumPall.ex Pursh
SHAO Cai,LIU Jiyong,ZHANG Hao,WANG Zhenxing(Insti-tute of Special Animals and Plant Sciences,Chinese Academy of Agricultural Sciences,Changchun 130112,China)
The anatomic structure of nutritive organs,such as root,stem,leaf and photosynthetic physiological properties of the leaves ofTrillium kamtschaticumwas studied with paraffin sectioning method and photosynthetic apparatus.The objective of the present study was to explain how the anatomical structuresandphotosyntheticphysiologicalcharacteristicsofTrilliumkamtschaticumtoadaptits environment,and could provide theoretical foundation and scientific basis for its protection and domestication cultivation.The results showed that it had the primary structure of root and stem.There was no cambium invascular tissue.The number of catheter was less and conducting tissue wasn't developed.There was no epidermal hair on the leaf epidermis and the epidermic cells were uneven.The mesophyll cells were only composed of sponge parenchyma cells.It arranged loosely and the intercellular space was large.There was no stoma in upper epidermis of leaf.There was stoma in lower surface of leaf and the stomatal density was 18.69 individual·mm-2.The stomas were convex.The ratio of the chlorophyll a/b was 1.90 and the light compensation point and the light saturation point of leaf was 1.23 and 415 μmol·m-2·s-1,respectively.The maximum net photosynthetic rate was 4.44 μmol·m-2·s-1.These results indicated thatTrillium kamtschaticum had the obviously morphological structure and photosynthetic physiological characteristic to adapt shade and wet environment.It was a typical shade plant.
Trillium kamtschaticum;anatomic structure;photosynthetic properties;environment adaptation
Q944.53
A
1005-9369(2015)05-0032-06
2015-01-12
吉林省科技支撑计划项目(20120909)
邵财(1982-),男,助理研究员,硕士,研究方向为药用植物资源学。E-mail:shaocai2003@163.com
