施肥和种间竞争对黄顶菊入侵土壤微生物的影响
2015-11-25郭艾英陈凤新张风娟
郭艾英, 杨 晴, 陈凤新, 叶 红, 张风娟*
(1. 河北科技师范学院生命科技学院, 秦皇岛 066600; 2. 河北大学生命科学学院,保定 071002; 3. 华中农业大学生命科技学院, 武汉 430070)
施肥和种间竞争对黄顶菊入侵土壤微生物的影响
郭艾英1, 杨 晴1, 陈凤新2, 叶 红3, 张风娟2*
(1. 河北科技师范学院生命科技学院, 秦皇岛 066600; 2. 河北大学生命科学学院,保定 071002; 3. 华中农业大学生命科技学院, 武汉 430070)
田间条件下建立了入侵植物黄顶菊(Flaveriabidentis)和伴生草本植物马唐(Digitariasanguinalis)、益母草(Leonurusjaponicus)单独种植(以下简称单种)和混合种植(以下简称混种,比例1∶1)小区,设置施氮肥和不施氮肥处理,研究两者对黄顶菊入侵土壤微生物的影响。结果表明:黄顶菊单种小区,土壤增施氮肥黄顶菊的生物量显著升高,且促进了真菌的生长,抑制放线菌、有机磷细菌和钾细菌的生长。黄顶菊与马唐竞争生长,施肥显著抑制了放线菌、有机磷细菌和钾细菌的生长,黄顶菊的竞争攻击力系数(aggressivity,以下简称AG)小于0,在竞争中黄顶菊不占优势;不施肥时细菌、放线菌、有机磷细菌的数量与黄顶菊生物量显著正相关(P<0.05),且AG>0,在竞争中黄顶菊占优势。黄顶菊与益母草竞争生长,施肥与否仅有机磷细菌数量显著高于裸土,且微生物数量与黄顶菊生物量都无显著相关性(P>0.05),黄顶菊的AG都小于0,在竞争中不占优势。由此,黄顶菊与马唐或益母草竞争生长,增施氮肥改变了土壤微生物数量,降低了黄顶菊的种间竞争能力,不利于实现黄顶菊的入侵。
黄顶菊; 马唐; 益母草; 竞争能力
外来植物入侵是导致世界范围内生态系统退化和生物多样性丧失的主要原因之一。早期对入侵植物与入侵地生态系统各组分间相互作用的研究主要集中在地上部分,对土壤生态过程影响的研究比较少[1-2]。现在人们认识到外来植物会改变入侵地土壤的生物(微生物群落)和非生物(养分循环)特性,这种改变会反过来影响外来植物的入侵进程,使其更适合自身生长,不利于入侵地本地植物的生长。已有研究表明,外来入侵植物紫茎泽兰、加拿大一枝黄花、薇甘菊、三叶鬼针草等入侵地土壤微生物群落结构发生了改变,影响了土壤微环境,为其进一步入侵提供有利条件[3-5]。
原产于南美洲巴西、阿根廷等国的黄顶菊[Flaveriabidentis(L.) Kuntze.]是我国近年来发现的一种一年生入侵杂草,截至2009年底,仅河北省的黄顶菊疫情已扩大至89县(市),受灾面积2.4万 hm2[6],且呈现以河北省中南部为中心,向周边其他省市扩散的趋势[7]。黄顶菊释放的化感物质能抑制植物种子的萌发和幼苗生长,严重影响入侵地生物多样性,威胁农业生产安全,目前对黄顶菊入侵研究主要集中在生物学特性[8]、传播途径[9]及对其他植物的化感作用[10-11]等方面,与本地植物的竞争也有报道,但仅局限在植物生物学特性[12-13]、土壤养分与酶活性[14-15]、土壤细菌多样性[16-18]、土壤真菌多样性[19]等方面。
施肥影响土壤微生物种群的数量和活性,影响土壤矿物质、有机质含量,进而影响作物产量及土壤肥力[20]。张天瑞等研究表明黄顶菊入侵地土壤全氮、铵态氮、硝态氮含量随入侵程度的增加而增加[21],而土壤氮含量增加会进一步影响黄顶菊和本地植物的竞争平衡关系,在黄顶菊与本地植物竞争生长中这是否有利于黄顶菊快速生长而形成单优群落呢?本试验通过人工施氮肥提高土壤含氮量,田间建立黄顶菊与伴生草本植物马唐[Digitariasanguinalis(L.) Scop.]、益母草(LeonurusjaponicusHoutt.)的单种和混种小区,研究黄顶菊与本地植物竞争生长过程中氮肥对黄顶菊根区土壤微生物的影响,旨在从微生物学角度研究黄顶菊入侵机制,为利用种间竞争特性对黄顶菊进行有效防控提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为黄顶菊和入侵域2种草本植物:马唐和益母草。
1.2 试验设计
1.2.1 田间试验
田间试验在河北科技师范学院昌黎校区农场进行(北纬39°42′,东经119°09′),土质为潮土,pH 7.6,电导率 531.0 μS,有机质含量1.70%,水解性氮108.35 μg/g,有效磷 26.21 μg/g,速效钾90.65 μg/g。本试验于2012年5月中旬至9月中旬进行,采用随机区组试验设计,试验处理分别为裸土(不种植任何植物,简称CK)、黄顶菊单独种植(以下简称单种,简称Fb)、马唐单种(简称Ds)、益母草单种(简称Lj)、黄顶菊与马唐混合种植(以下简称混种,简称Fb+Ds)、黄顶菊与益母草混种(简称Fb+Lj);每个处理分为施氮肥和不施氮肥两个水平,肥料为尿素,添加量为30 g/m2,于播种前一次性加入。每个处理种植面积为3.0 m×1.0 m,种植密度为80 株/m2,混种处理中,2种植物的种子进行混合然后播种,待植物长出幼苗,再进行间苗处理,使每个小区1种或2种植物共32株,混种比例为1∶1,随机排列,小区之间为宽度0.2 m的隔离带,每个处理5次重复。试验期间,及时人工拔除其他杂草。
于2012年9月中旬植物处于结实后期采样,每个小区采用对角线采样法采集土壤,除去落叶层后,每个小区对角采集5~10个点,单种区在主根周围向下垂直取5~20 cm土壤混合,混种区选取对角线上距两种植物的中心部位土壤混合,分别装袋后放入冰盒内带回实验室。新鲜土样过0.5 cm筛后测定土壤微生物种群与数量。
1.2.2 土壤的微生物群落测定
土壤微生物数量采用固体平板稀释法进行分离测定。细菌、真菌、放线菌分别用牛肉膏蛋白胨琼脂培养基、马丁氏培养基和改良高氏1号培养基培养;有机磷细菌和钾细菌分别采用有机磷细菌培养基和硅酸盐细菌培养基培养。选取菌落数(colony-forming unit, cfu)在30~300之间的平板进行计数。
1.2.3 生物量测定
2012年9月,分别选取每个处理中长势良好的4~8株植物,将其分别分为根、茎、叶三部分,除去表面灰尘及杂质后于85 ℃恒温干燥箱烘干至恒重,并用10-4的电子天平称各部分干重。
1.2.4 数据分析
选择相对竞争强度 (relative competition intensity,RCI)[22]和竞争攻击力系数(aggressivity,AG)[23]来评判本试验中的物种竞争能力。计算公式分别为:
(1)
(2)
式中,Y表示每种植物的生物量;a、b代表入侵种、本地种;p、q表示物种a、b的混栽比例,p+q=1;Ya表示物种a单独种植时的生物量;Yb表示物种b单独种植时的生物量;Yab表示物种b存在的情况下物种a的生物量;Yba表示物种a存在的情况下物种b的生物量;(1)中的a,b可以互换。
RCI=0表明两物种间不存在竞争,0
试验数据采用SPSS 13.0软件对微生物数量和生物量进行统计方差分析,对数据进行显著性检验,再对两组数据作相关性分析。
2 结果与分析
2.1 不同处理对不同植物根区土壤微生物总量的影响
由表1可得,试验小区各种植方式(F=17.91,P=0)和施肥方式(F=106.23,P=0)对不同植物根区土壤微生物总量的影响达到了显著差异,说明种植方式及施肥方式都显著影响根区土壤微生物总量。

表1 种植和施肥对土壤微生物总量的影响
施肥后入侵植物黄顶菊单种小区土壤微生物总量显著降低,且低于裸土,而马唐单种、黄顶菊与马唐混种小区微生物总量显著升高(表2),说明施氮肥不利于黄顶菊单种根区土壤微生物的生长,促进了马唐单种和黄顶菊与马唐混种小区土壤微生物的生长,对益母草小区微生物的生长无显著影响。
土壤增施氮肥黄顶菊单种根区的微生物总量显著低于马唐单种、黄顶菊与马唐混种小区,不施肥时黄顶菊单种根区微生物总量高于其他种植小区(表2),说明施氮肥对马唐根区土壤微生物的生长更有利,不施氮肥时黄顶菊根系分泌物有利于土壤微生物的生长。

表2 施肥处理对黄顶菊和草本植物根区土壤微生物总量的影响1)
1) CK为裸土对照,Fb、Ds、Lj分别表示黄顶菊、马唐和益母草单种群落、Fb+Ds表示黄顶菊与马唐混种群落、Fb+Lj为黄顶菊与益母草混种群落,数据表示为平均值±标准误;数据后小写字母表示在0.05水平上的差异显著性。下表同。
Fb, Ds and Lj indicateF.bidentis,D.sanguinalisandL.japonicusplanted singly, respectively. Fb+Ds indicatesF.bidentisplanted withD.sanguinalis; Fb+Lj indicatesF.bidentisplanted withL.japonicus.Data in the table are mean±SE; data followed by different lowercase letters indicate significant difference at 0.05 levels, and the same below.
2.2 不同处理对黄顶菊、马唐和益母草根区土壤微生物菌群的影响
土壤增施氮肥处理,黄顶菊单种小区土壤放线菌、有机磷细菌、钾细菌数量显著降低,真菌数量显著增多;混种小区中,黄顶菊与马唐混种放线菌、有机磷细菌、钾细菌数量显著降低,细菌和真菌数量显著升高;黄顶菊与益母草混种显著降低了放线菌、钾细菌数量,有机磷细菌数量显著升高(表3)。说明土壤含氮水平显著影响了黄顶菊、马唐、益母草根区微生物生长,对单种和混种小区的影响不同。
与黄顶菊单种相比,施肥显著提高了黄顶菊与马唐混种小区土壤细菌、放线菌、钾细菌的数量,黄顶菊与益母草混种小区有机磷细菌的数量显著提高,而真菌数量降低(表3),该结果说明黄顶菊与马唐混种,施肥对细菌、放线菌、钾细菌的生长有利,与益母草混种施肥利于有机磷细菌的生长,抑制真菌活性。
单种小区中,施肥时黄顶菊单种土壤的细菌数量显著低于马唐,与益母草无显著差异,放线菌、有机磷细菌数量均显著低于马唐、益母草单种,而真菌和钾细菌数量显著高于马唐、益母草单种(表3),这表明与马唐、益母草相比,施肥有利于黄顶菊根区土壤真菌和钾细菌的生长,抑制了放线菌和有机磷细菌数量,马唐根分泌物利于细菌的生长。
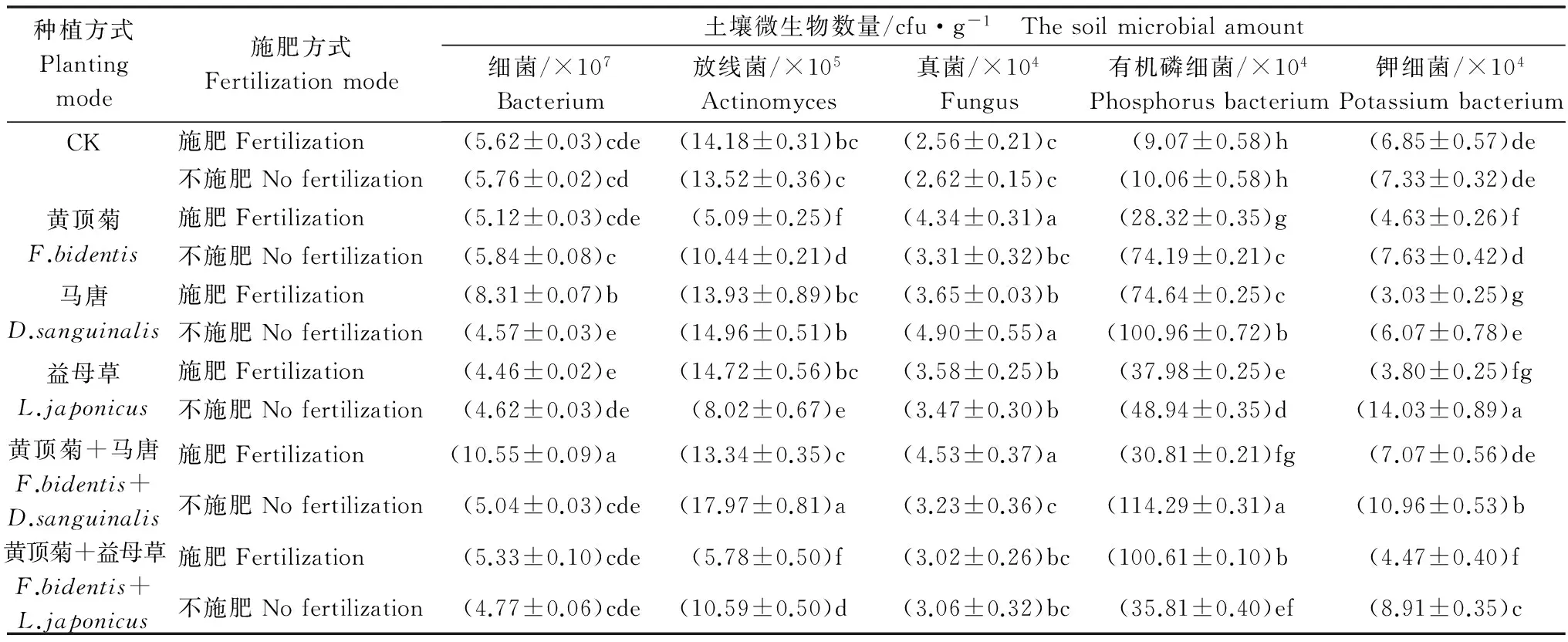
表3 施肥处理对黄顶菊、马唐和益母草根区土壤微生物数量的影响
2.3 不同处理黄顶菊和马唐、益母草竞争能力的比较
由表4可得,黄顶菊与马唐竞争生长,无论土壤是否施肥,黄顶菊和马唐的RCI都大于0,说明两者的生长都受到了抑制。施肥时黄顶菊的AG值小于0,不施肥时AG大于0,说明土壤增施氮肥降低了黄顶菊的种间竞争力,改变了两者的竞争关系,在竞争中黄顶菊不占优势。
黄顶菊与益母草竞争生长,土壤施肥与否,黄顶菊的RCI大于0,施肥时益母草的RCI大于0,不施肥RCI小于0,说明施肥时两者的生长都受到了抑制,不施肥时黄顶菊促进了益母草的生长。施肥与否黄顶菊的AG都小于0,说明在竞争中黄顶菊不占优势。

表4 黄顶菊、马唐、益母草的竞争生长指标
2.4 不同处理条件下黄顶菊的生物量与根区土壤微生物数量之间的相关性
从上述分析得出,试验小区不同的种植方式和施肥方式显著影响了土壤微生物数量和黄顶菊的竞争能力,由此对黄顶菊的生物量和土壤微生物数量之间进行了相关性分析(表5),结果表明:试验各小区黄顶菊的生物量与土壤微生物的数量之间都呈正相关关系,说明土壤微生物数量影响黄顶菊的生长。单种小区中,施肥时黄顶菊的生物量与放线菌、真菌和有机磷细菌数量显著正相关,不施肥时与土壤微生物总量、细菌、放线菌和有机磷细菌数量显著正相关,说明土壤含氮量不同影响黄顶菊生长的微生物数量和种类也不同。混种小区中,与马唐竞争生长,施肥时黄顶菊的生物量与放线菌、有机磷细菌数量显著正相关,不施肥则与细菌、放线菌和真菌数量显著正相关;与益母草竞争生长,施肥与否黄顶菊的生物量与微生物数量之间都无显著相关性。此结果说明黄顶菊与马唐竞争生长,施肥与否小区的放线菌和有机磷细菌的生长对黄顶菊生长都有促进作用,且土壤含氮量低时细菌对黄顶菊的生长也有促进作用;与益母草竞争生长过程中,施肥与否,土壤微生物数量的多少对黄顶菊的生长都无显著影响。

表5 黄顶菊的生物量与根区土壤微生物数量的相关系数1)
1)*表示在0.05水平上显著相关。
*indicate significant correlation at 0.05 level.
3 讨论
土壤微生物是维持土壤质量的重要因素,植物根系分泌物可改变土壤微生物的种群结构[24],其数量与土壤养分含量存在一定的相关性[25],因而间接影响附近植物的生长。本研究表明,土壤施氮肥时黄顶菊、马唐、益母草单种或混种小区根区土壤微生物的总量不同,其中三大类微生物和细菌生理类群的数量存在较大差别,在竞争生长中对黄顶菊、马唐或益母草的生物量的影响也不同。
施肥时黄顶菊单种根区的微生物总量较不施肥处理低,但黄顶菊的生物量较高,这与Mandic 等研究得到的高水平施氮肥对土壤微生物总量与马铃薯产量的影响结果一致[26],这可能是氮肥提高了土壤有机质含量,促进植物生长,致使黄顶菊的生物量增大。与施肥处理相比,不施肥时不同植物小区仅黄顶菊单种根区的微生物总量显著升高,且细菌、放线菌和有机磷细菌数量与黄顶菊的生物量显著正相关,可能是土壤含氮水平较低时,黄顶菊根系分泌物促进了细菌和有机磷细菌、放线菌的生命活动,提高了土壤养分的有效性和利用率,从而促进了黄顶菊生长,这可能是黄顶菊能够在贫瘠的土壤中旺盛生长的原因;在与马唐或益母草竞争生长时,不施肥时黄顶菊的生物量都显著高于施肥处理,可能也与上述原因有关。多数放线菌具有一定的抑菌活性,本研究也发现黄顶菊根区具有抑菌作用的放线菌数量较马唐、益母草单种小区多,样液稀释梯度越小其抑菌活性越强,且无论是否土壤施肥与黄顶菊的生物量均显著正相关,这是否有助于提高黄顶菊拮抗土传病原菌的能力有待进一步研究。
黄顶菊与马唐竞争生长,施肥与否两者都受到了对方强有力的竞争,施肥时这种对黄顶菊的抑制作用强于非施肥处理,这说明两者竞争生长过程中土壤含氮水平的增加有助于提高马唐的竞争能力,吕远等研究结果表明田间试验也发现土壤氮含量为1.42 g/kg时黄顶菊对马唐并未表现出抑制作用[13],而不施肥时黄顶菊较强的竞争力可能得益于其根围的微生物活性;黄顶菊与益母草竞争生长,无论施肥与否黄顶菊都受到益母草的有力竞争,而不施肥时黄顶菊促进了益母草的生长,且在竞争中无论施肥与否益母草都占优势,说明在两者竞争生长的过程中,益母草可有效地抑制黄顶菊的生长,且土壤含氮量较小时,黄顶菊的生长在一定程度上促进了益母草的生长。
牛红榜研究得到紫茎泽兰入侵后土壤微生物各生理功能类群的变化与土壤理化性质的变化显著相关,由此土壤微生物群落的变化可能导致了土壤养分水平的升高[4];王蕾在研究黄顶菊入侵对土壤理化性质影响时得出土壤细菌的变化与土壤有效磷、速效钾的变化极显著相关[27]。本研究表明施肥与否黄顶菊单种小区细菌数量与裸土相比无显著差异,但细菌生理类群中有机磷细菌数量显著升高,与黄顶菊的生物量显著正相关。张天瑞等研究表明黄顶菊入侵土壤全磷、速效磷含量都有所下降[21],这可能是由于黄顶菊根区有机磷细菌数量升高,降解土壤中磷矿物,为作物提供有效磷而导致土壤全磷含量降低,且黄顶菊对速效磷有较强的吸收和利用能力[21],由此导致速效磷含量降低;黄顶菊单种小区根区钾细菌数量施肥时与裸土相比显著降低,与黄顶菊的生物量无显著相关性。王蕾研究得出黄顶菊入侵地土壤速效钾含量降低了[27],这可能与土壤钾细菌数量较小有关,但这些细菌生理类群是怎样参与并影响土壤肥力促进黄顶菊生长有待于进一步考证。
[1] Ehrenfeld J G.Effect of exotic plant invasions on soil nutrient cycling processes [J].Ecosystems, 2003,6(6):503-523.
[2] Levine J M, Vilà M, Antonio C M D, et al.Mechanisms underlying the impacts of exotic plant invasions [J].Proceedings Biological Sciences, 2003, 270: 775-781.
[3] 李国庆. 入侵植物加拿大一枝黄花对根际土壤微生物群落多样性的影响研究[D].福州: 福建农林大学,2009.
[4] 牛红榜. 外来植物紫茎泽兰入侵的土壤微生物学机制[D].北京: 中国农业科学院, 2007.
[5] 陈亮, 李会娜, 杨民和, 等. 入侵植物薇甘菊和三叶鬼针草对土壤细菌群落的影响[J].中国农学通报,2011,27(8):63-68.
[6] 皇甫超河, 王楠楠, 陈冬青, 等. 增施氮肥对黄顶菊与高丹草苗期竞争的影响[J].生态环境学报,2010,19(3):672-678.
[7] 刘玉升,刘宁,付卫东,等.外来入侵植物-黄顶菊山东省发生现状调查[J].山东农业大学学报:自然科学版,2011,42(2):187-190.
[8] 刘全儒. 中国菊科植物新归化属黄菊属[J].植物分类学报,2005, 43(2):178-180.
[9] 陆秀君, 董立新, 李瑞军, 等. 黄顶菊种子传播途径及定植能力初步探讨[J].江苏农业科学,2009(3): 140-141.
[10]任艳萍, 江莎, 古松, 等. 外来植物黄顶菊根、茎、叶的化感作用初探[J].植物保护, 2009, 35(3):36-40.
[11]张风娟,徐兴友,陈凤敏,等.黄顶菊茎叶浸提液对白菜和水稻幼苗化感作用的初步研究[J].西北植物学报,2008,28(8):1669-1674.
[12]许文超, 徐娇, 陶哺, 等. 外来入侵植物黄顶菊的化感作用初步研究[J].河北农业大学学报,2007,30(6):63-67.
[13]吕远, 王贵启, 郑丽, 等. 入侵植物黄顶菊与本地植物的竞争[J].生态学杂志, 2011, 30(4): 677-681.
[14]皇甫超河, 张天瑞, 刘红梅, 等. 三种牧草植物对黄顶菊田间替代控制[J].生态学杂志, 2010, 29(8):1511-1518.
[15]马杰, 皇甫超河, 易津, 等. 4种替代植物对黄顶菊入侵土壤养分和酶活性的影响[J].生态环境学报,2011,20(5):805-812.
[16]闫素丽, 皇甫超河, 李刚, 等. 四种牧草植物替代控制对黄顶菊入侵土壤细菌多样性的影响[J].植物生态学报,2011, 35(1):45-55.
[17]纪巧凤, 宋振, 张国良, 等. 黄顶菊入侵对土壤磷细菌多样性的影响[J].农业资源与环境学报,2014,31(2):175-181.
[18]赵晓红,皇甫超河,曲波,等. 黄顶菊入侵对土壤氨氧化细菌群落结构及多样性的影响[J].生态学杂志,2014,33(6):1-9.
[19]常瑞恒,皇甫超河,杨殿林,等.生物替代对黄顶菊根际土壤真菌群落多样性的影响[J].中国农学通报,2011,27(33):60-66.
[21]张天瑞, 皇甫超河, 白小明, 等. 黄顶菊入侵对土壤养分和酶活性的影响[J].生态学杂志, 2010, 29(7):1353-1358.
[22]Grace J B.On the measurement of plant competition intensity[J].Ecology,1995, 76: 305-308.
[23]McGilchrist C, Trenbath B.A revised analysis of plant competition experiments [J].Biometrics, 1971,27:659-671.
[24]Klironomos J N.Feedback with soil biota contributes to plant rarity and invasiveness in communities [J].Nature,2002,417(6884):67-70.
[25]湛方栋, 何永美, 陈建军, 等. 施氮量对丘北辣椒土壤微生物数量的影响[J].云南农业大学学报,2010, 25(6):854-861.
[27]王蕾. 外来植物黄顶菊入侵对土壤理化性质及生物活性的影响[D].合肥: 安徽农业大学,2008.
(责任编辑:田 喆)
Effects of fertilization and interspecific competition on the microbes in rhizosphere soils invaded byFlaveriabidentis
Guo Aiying1, Yang Qing1, Chen Fengxin2, Ye Hong3, Zhang Fengjuan2
(1. College of Life Science & Technology, Hebei Normal University of Science & Technology, Qinhuangdao 066600, China; 2. Institute of Life Sciences, Hebei University, Baoding 071002, China; 3. College of Life Science and Technology, Huazhong Agricultural University, Wuhan 430070, China)
The effect of nitrogen fertilizer on the microbes in rhizosphere soils ofFlaveriabidentisin competitive growth with two native species,Digitariasanguinalis(L.) Scop. andLeonurusjaponicusHoutt., planted singly or mixed withF.bidentis(1∶1), respectively, was studied in the field. The results showed that nitrogen addition increased the biomass ofF.bidentisand promoted the growth of fungi in the monoculture ofF.bidentis, but decreased actinomycetes, phosphorus bacteria and potassium bacteria. WhenF.bidentiscompeted withD.sanguinalis,nitrogen addition was significantly inhibited the growth of actinomycetes, phosphorus bacteria and potassium bacteria. The aggressivity of the invasive species was less than 0, so the competition was disadvantageous forF.bidentis. A positive significant correlation existed between the counts of bacteria, actinomycetes, phosphorus bacteria and the biomass ofF.bidentistreated with non-fertilizer control. The aggressivity of the invasive species was more than 0, andF.bidentiswas dominant in the competition. WhenF.bidentiscompeted withL.japonicus, the microbes were less than in bare soils except for phosphorus bacteria, and no matter fertilizer added or not, there was no significant correlation between soil microbial quantity and the biomass ofF.bidentis. The aggressivity of the invasive species was less than 0, indicating that the competition was advantageous toF.bidentis. To sum up, nitrogen addition changed the quantity of soil microorganisms and decreased the competitive ability of the invasion species.
Flaveriabidentis;Digitariasanguinalis;Leonurusjaponicus; competitive ability
2014-04-26
2014-07-28
国家自然科学基金项目(31372000,31171906);河北省自然科学基金项目(C2015201021)
S 45
A
10.3969/j.issn.0529-1542.2015.03.011
* 通信作者 E-mail: fengjuanzhang@126.com
