烤烟内生高温菌YYFG3的分离鉴定及所产蛋白酶活性
2015-11-23孙会忠王小东朱金峰李广良孙明辉许自成宋月芹
孙会忠,王小东,朱金峰 ,李广良,孙明辉,许自成,宋月芹
1 河南科技大学农学院,河南洛阳 471003;
2 河南省烟草公司漯河市公司,河南漯河市 462000;
3 河南农业大学烟草学院,郑州 450002
烤烟内生高温菌YYFG3的分离鉴定及所产蛋白酶活性
孙会忠1,王小东1,朱金峰2,李广良2,孙明辉2,许自成3,宋月芹1
1 河南科技大学农学院,河南洛阳 471003;
2 河南省烟草公司漯河市公司,河南漯河市 462000;
3 河南农业大学烟草学院,郑州 450002
通过固体分离培养基的分离培养和牛奶琼脂鉴别培养基的鉴别初筛,从烟草(Nicotiana tabacum L.)的初烤烟叶中分离到一株嗜热产蛋白酶菌株,编号为YYFG3,其生长温度范围是30℃-65℃,最适生长温度55℃左右,对pH的耐受范围是5-9,最适范围是6-7;在发酵产酶培养基中,56℃、180 r/min条件下发酵32 h的蛋白酶活力达到最大值35.3 U/mL,故将YYFG3定性为产蛋白酶高温菌株。通过光学显微镜和扫描电子显微镜对该菌株形态特征的观察、相关生化特性的测定、16S rDNA的克隆测序以及对菌株分子遗传进化树的构建,确定菌株YYFG3隶属于土芽孢杆菌属Geobacillus,暂将其命名为Geobacillus sp.YYFG3。菌株YYFG3可作为产蛋白酶高温微生物诱变育种和全基因组育种的良好材料,具有良好的开发应用潜力。
烟草;高温菌;分离;蛋白酶;16S rDNA
高温菌株往往具有与常温菌株明显不同的生物学特性,它们一般具有高度的特异性,尤其是高温菌所产的胞外酶活性在高温条件下能够保持很好的稳定性,克服了在应用过程中常温酶化学性质不稳定的局限性,能够更好的适应和满足工业化开发利用的需求,已经开发成功的具有嗜热特性高温酶已达几十种,如蛋白酶、脂肪酶、淀粉酶、糖苷酶、漆酶、硫酸化酶等[1]。蛋白酶占世界酶制剂市场的65%以上,广泛应用于制药、纺织、食品、皮革等领域[2]。近年来高温菌及其所产酶在应用领域扩展迅速,如烟料发酵[3]、菌肥开发[4]、垃圾处理[5]等领域。因为高温蛋白酶的热稳定性一般与其产酶菌株的耐热能力成正比,不管是开发应用还是理论研究,首要条件是分离获得高温菌株,而且来源于微生物的蛋白酶都是胞外酶,比动植物来源的蛋白酶易于加工,因此,从不同材料分离高温菌有着重要的现实意义,一直是功能微生物的研究热点之一。
植物内生菌是一个巨大的遗传多样性宝库,植物为内生菌提供了多元复杂的生境,内生菌所具备的独特的生物学特性[1],为高温菌的分离和开发提供了更多可能[6]。烟草(Nicotiana tabacum L.)富含生物碱[7],有着比较特殊的微生物内生环境,而烟叶烘烤是在较高温度下进行的一个与物理变化相伴随的复杂的生理生化过程[8],该过程会促使具有特殊功能内生菌的存在。迄今为止,尚未见关于烤烟产蛋白酶高温内生菌株分离鉴定的研究报道。基于以上背景,本文拟以烤烟为试验材料探索分离培养具有产蛋白酶活性的高温菌株,旨在丰富产功能酶高温菌资源。
1 材料与方法
1.1 实验材料
烤烟品种:中烟100,由河南省烟草公司漯河市公司提供。
固体分离培养基:蚕蛹干粉3 g,酵母粉5 g,吉兰糖胶2 g,海藻糖0.5 g,菊糖0.3 g,NaCl 9 g,牡丹根际土壤浸汁200 mL,琼脂粉18 g,蒸馏水1 000 ml,自然pH。
牛奶琼脂鉴别培养基:牛肉膏3 g,蛋白胨5 g,NaCl 5 g,脱脂奶粉15 g,琼脂粉20 g,蒸馏水1 000 mL,pH7.0。
LB液体培养基:蛋白胨10 g,酵母粉5 g,NaCl 5 g,蒸馏水 1 000 ml,pH7.0。
发酵产酶培养基:牛肉膏6 g,大豆蛋白胨15 g,KH2PO41.3 g,K2HPO43 g,CaCl22 g,NaCl 2 g,MgSO4·7H2O 0.05 g,FeSO40.001 g,蒸馏水 1 000 mL,pH7.0。
1.2 实验方法
1.2.1 菌株的分离及筛选
烤烟材料的预处理:将新鲜烟叶用无菌水充分水洗后,常规烘烤。烘烤完毕,立即装入无菌袋中备用。
取2片备用叶材料,用无菌水冲洗,之后用75%乙醇进行叶表面浸泡消毒3min,紧接着再次用0.1 mol/L升汞浸泡消毒3min,消毒完成后用无菌水漂洗3次,每次3min,漂洗完成后,在无菌操作条件下用研钵研碎,转入灭菌三角瓶后,加入100 mL无菌水,充分混匀。分别取 50 µL、100 µL、150 µL 不同体积上清液涂布于固体分离培养基,每个体积涂6个平板,恒温培养箱中55 ℃培养48 h后,挑取菌落连续进行3次平板划线纯化,并将纯化菌株甘油-20℃保存。取80 µL最后一次漂洗液涂板同步培养作为对照,无杂菌生长视为消毒成功。
将纯化菌株接种于牛奶琼脂鉴别培养基进行产蛋白酶初筛,55℃恒温培养48 h,将具有透明圈者初步定性为具有产蛋白酶活性高温菌株。
1.2.2 菌株生长温度范围及最适温度测定
菌株在LB液体培养基中活化培养后(OD600=0.4),以1%的接种量转接至新鲜LB液体培养基,分别在20℃、25℃、30℃、35℃、45℃、50℃、55℃、60℃、65℃和70℃温度下,180 r/min摇床培养36 h。通过测定菌液OD600确定其生长温度范围和最适生长温度。设3次重复。
1.2.3 菌株生长pH耐受范围测定
菌株在LB液体培养基中活化培养后(OD600=0.4),以1%的接种量转接至新鲜LB液体培养基,培养基pH分别为3.0、4.0、5.0、6.0、7.0、8.0、9.0和10.0等8个梯度,在55℃、180 r/min摇床培养24 h,通过测定菌液OD600确定其pH耐受范围及最适pH。设3次重复。
1.2.4 菌株生长与产蛋白酶的关系
菌株在LB液体培养基中活化培养后(OD600=0.4),以4%接种量接种于发酵产酶培养基,55℃、180 r/min摇床连续培养60 h,期间每隔4 h取样一次,分别测定发酵液OD600和蛋白酶活力。设3次重复。蛋白酶活力采用Folin-酚法测定,步骤参考文献[9]进行。
1.2.5 菌株的鉴定
①表型及生化特征
应用光学显微镜和扫描电子显微镜对菌株进行形态观察[10];生理生化指标测定方法参考文献[11]、[12]进行。
②菌株的16S rDNA分析
将目标菌株接种于LB培养基,37 ℃,180 r/min摇床培养12 h,获取菌液。采用TaKaRa MiniBEST Bacterial Genomic DNA Extraction kit Ver. 2.0 试剂盒提取YYFG3基因组DNA。
扩增引物:正向引物27F:5′ -AGAGTTTGATCCT GGCTCAG-3′ ,反向引物:1492R:5′ -TAGGGTTA CCTTGTTACGACTT-3′ 。 扩 增 体 系(20 µL):10×Ex Taq PCR buffer 2 µL,dNTP(10 mmol/mL)1.6 µL,正向和反向引物(20 µm/mL)各 1 µL,DNA 模板 1 µL,Ex Taq DNA 聚合酶 0.2 µL,超纯水13.2 µL。扩增程序:94 ℃预变性5min;94 ℃变性 50 s,50 ℃退火 30 s,72 ℃延伸 1min 30 s,循环35次;72 ℃最终延伸10min。目的片段长度约为1.5 kb,PCR产物经1 %琼脂糖凝胶电泳后,采用TaKaRa MiniBEST Agarose Gel DNA Extraction kit Ver. 3.0对目的条带进行胶回收,回收产物与TaKaRa pMDTM18-T Vecter质粒载体连接,并将重组质粒转化于大肠杆菌DH5α感受态细胞,37 ℃培养12 h,蓝白斑筛选后,选取阳性克隆摇菌送测(由北京奥克鼎盛生物科技有限公司完成)。
将测序获得的16S rDNA序列提交NCBI数据库在线进行相似性比对,采用MEGA6.06软件构建Neighbor-Joining分子遗传进化树。菌株形态学和生化鉴定参考文献[13]、[14]和[15]进行。
2 结果与分析
2.1 目标菌株的获得
通过固体分离培养基的分离和牛奶琼脂培养基的鉴别培养,分离筛选出一株水解圈大而显著的菌株,编号为YYFG3,其水解圈直径/菌落直径比为4.6(图1),初步确定为目标菌株。

图1 YYFG3在牛奶琼脂平板上的水解圈Fig. 1 Hydrolysis circle of strain YYFG3 in milk culture medium
2.2 YYFG3的生长特性
由图2可知,菌株YYFG3在LB培养基中培养时,25℃以下和70℃以上均不能生长,30℃和65℃能够生长,但长势较弱,其它温度下菌株均能够较好生长,55℃培养时的OD600值最大,所以,菌株对温度的响应范围在30℃-65℃之间,最适生长温度55℃左右。图3则表明,菌株YYFG3生长的pH耐受范围是5≤pH≤9,最适生长pH6-7。
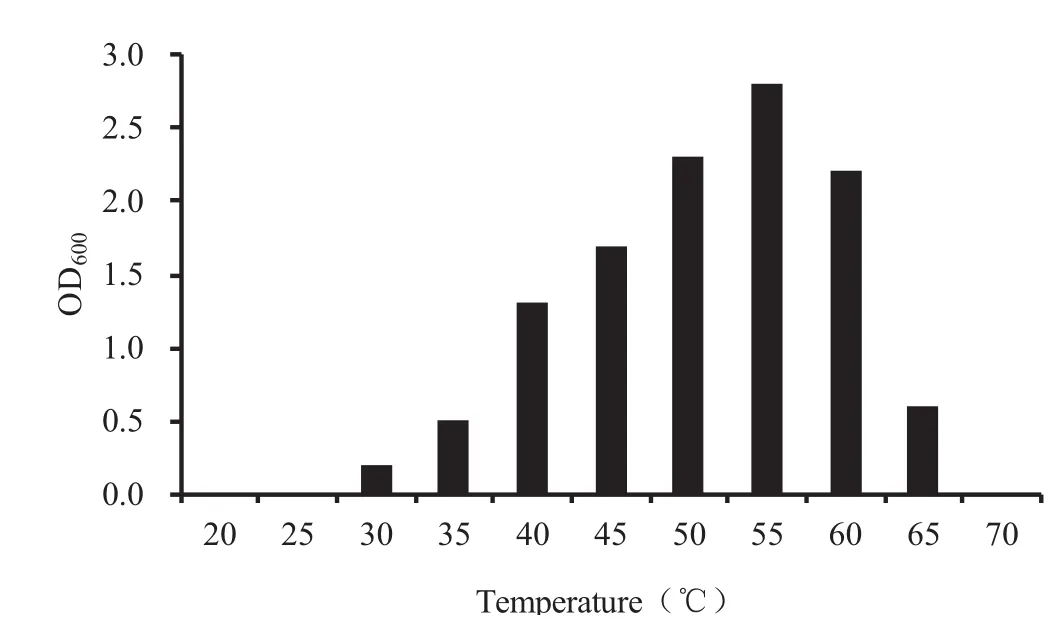
图2 温度对菌株生长的影响Fig.2 Effect of temperature on bacterial growth

图3 pH对菌株生长的影响Fig.3 Effect of pH on bacterial growth
2.3 YYFG3的生长与产蛋白酶曲线
如图4,菌株YYFG3在发酵产酶培养基中连续培养约60 h的历程中,菌株生长曲线呈现较为典型的“S”型曲线,发酵约8 h开始进入对数生长期,产蛋白酶活性在发酵24 h时跃变较为明显,从发酵28 h开始,蛋白酶活性开始进入高点,直至52 h,酶活性始终比较稳定,说明产酶能力主要在发酵后期菌体生物量最大的稳定期。发酵约44 h,菌株开始进入衰亡期,但产酶活性却下降平缓,说明产酶活性不但集中菌株生长曲线的稳定期和衰亡前期,而且酶活性持续时间长且较为稳定。所以,菌株YYFG3蛋白酶产量与生长曲线具有一定相关性。
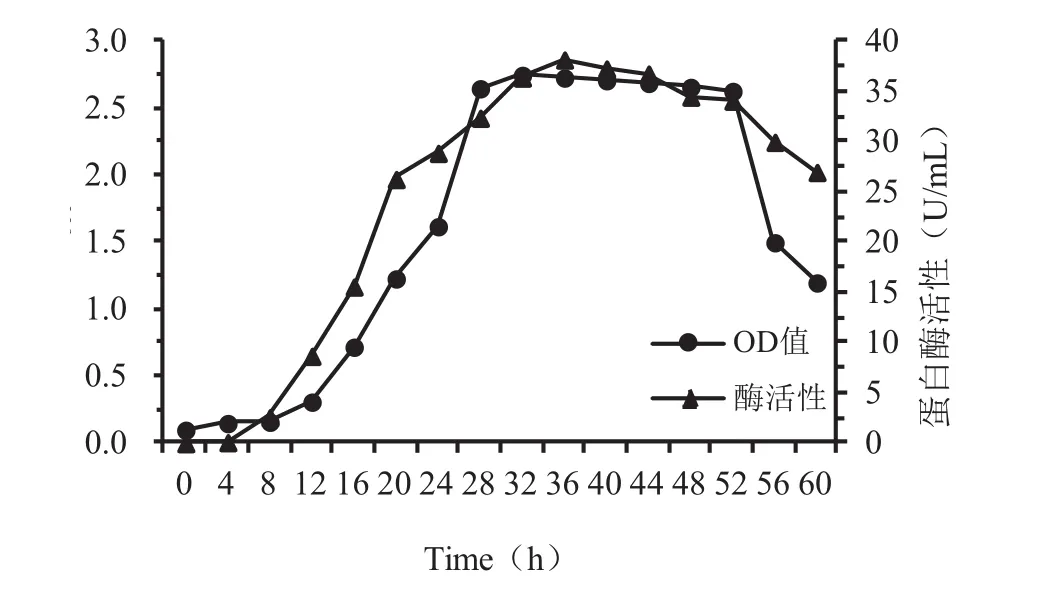
图4 菌株YYFG3的蛋白酶活性Fig.4 Enzyme production of strain YYFG3
2.4 菌株YYFG3的鉴定
2.4.1 表型特征
菌株YYFG3大小 1.1-2.8μm×0.8-1.6μm,无荚膜,能运动;在LB平板上55 ℃培养18 h,可观察到单菌落近圆形,边缘不整齐,直径2-4mm,表面粗糙,隆起不十分明显,扁平状,菌落灰白色(图5-A);芽孢椭圆形,位于中间或次极端(图5-B)。扫描电镜下可见典型直杆型菌体,两端钝圆,具侧生鞭毛(图5-C)。

图5 菌株YYFG3的表型特征Fig.5 The general characteristics of strain YYFG3
2.4.2 生理生化特性
对菌株YYFG3进行的生理生化指标测定结果表明,菌株相关生化特征与文献[13]和[14]和[15]中土芽孢杆菌属描述基本一致(表1)。
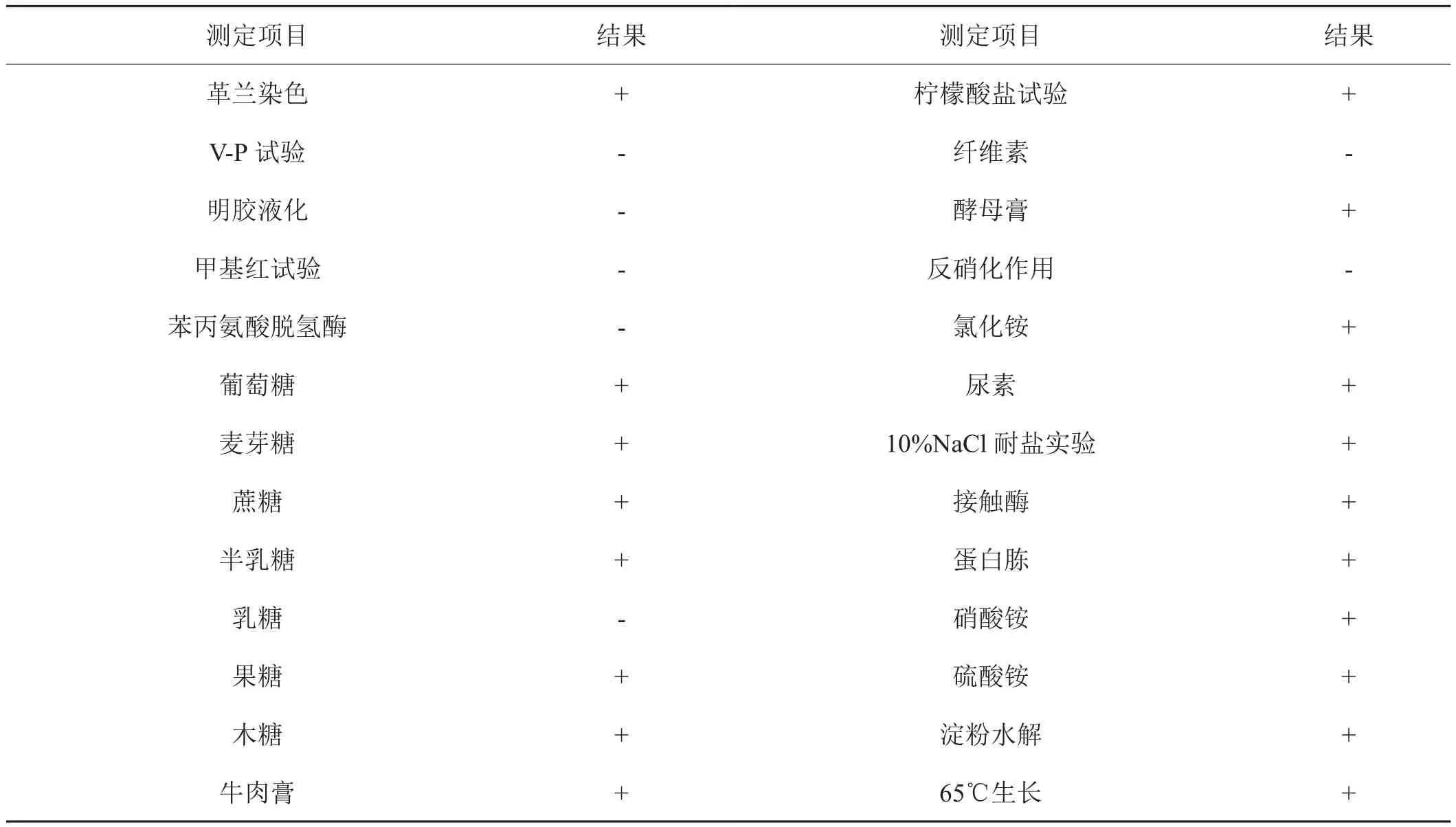
表1 菌株YYFG3的生理生化指标测定结果Tab.1 Determination results of physiological and biochemical index of strain YYFG3
2.4.3 16S rDNA分析
菌株YYFG3的16S rDNA序列PCR扩增结果如图6所示。

图6 菌株YYFG3的16S rDNA序列扩增结果Fig.6 The ampli fi cation results of 16S rDNA sequence of strain YYFG3
将目的条带进行切胶回收,并与质粒连接后,转化DH5α感受态细胞,送测阳性克隆获得的YYFG3菌株16S rDNA序列长度为1517 bp,将结果提交GenBank,序列号为KR362555。将菌株16S rDNA基因序列提交NCBI(National Center for Biotechnology Information)数据库的blast程序进行相似性比对,并选取相似度高的菌株序列构建菌株YYFG3的分子遗传进化树(图7)。结果显示,菌株YYFG3与土芽孢杆菌属的Geobacillus pallidus strain P4和G. Pallidus strain BGSC109A1聚为同一分支,与二者的相似性均达到99%,且bootstrap值也达到99%。综合该菌株的形态特征、生理生化特性和分子遗传进化树分析,将菌株YYFG3归为土芽孢杆菌属Geobacillus,暂命名为Geobacillus sp. YYFG3。

图7 基于16S rDNA基因序列的YYFG3系统发育进化树Fig. 7 Phylogenetic tree based on 16S rDNA gene sequences of YYFG3
3 结论与讨论
通过本研究,主要得出以下结论:(1)对烤烟进行严格的表面消毒后,仍然能够分离到能够耐受65℃高温菌活体纯培养物,说明烟叶经过烘烤后,确实有高温菌的存在;(2)菌株YYFG3在56℃、180 r/min条件下发酵发酵历程中,蛋白酶活力最高达到了35.3 U/mL,说明该菌株在高温条件下具有较高的产蛋白酶活性,在烤制过程中对品质形成可能起着一定的作用;(3)菌株YYFG3经过多项分类鉴定,确定隶属于土芽孢杆菌属(Geobacillus),该属是从原芽孢杆菌属(Bacillus)分离出来的一个新属,具有嗜热的共同特性,从系统归属的角度来看,YYFG3菌株具有产耐高温功能酶的可能。
高温菌一般的温度界定范围是最低生长温度为25℃-50℃,最适生长温度为55℃,最高生长温度为60~70℃[1]。参照这个标准,菌株YYFG3在30-65℃温度范围均可以生长,55℃生长最好,完全符合高温菌对温度的响应特征,可以定性为高温菌菌株。从高温、中温、低温环境中都能够分离获得高温菌,但从高温环境筛选获得的高温菌产功能高温酶的可能性也更大,酶的热稳定性好,具有更大的开发应用潜力[16]。鲜烟叶在烘烤过程中,淀粉酶、蛋白酶、氧化还原酶等对烤烟品质的形成起着重要的作用,而一系列酶的活性及作用时间与烘烤温度密切相关,常温菌在烘烤的高温阶段会死亡或失去活性,烘烤特殊的温度环境则有利于高温菌的活动,但关于烤烟内生高温菌的研究鲜见报道。在本实验的操作过程中,充分关注了无菌操作,对实验材料在烘烤前进行了多次漂洗,尽可能去除叶片表面的附生菌群;烘烤完成后,又进行了二重表面消毒,尽可能的降低了叶表面附生菌群的干扰。
不同来源的微生物往往具有不同的生物学特性,即便是同一种微生物,也会由于生境的不同而表现出某些生理特性上的显著差异[1]。本实验分离到的烤烟内生菌YYFG3的嗜热特性显著,pH耐受范围较广,初始产蛋白酶活性也较高,所以能够更好的适应不同工业上加工工艺的要求,在应用方面具有广阔前景。本实验证实,烟草营养器官内除了具有丰富的常温菌细菌外[17],也蕴藏着宝贵的高温菌资源。微生物的产酶活性与培养基类型和发酵条件密切相关[18],后续将对高温菌株YYFG3所产的蛋白酶学性质及与烤烟品质形成的关系展开深入研究。
[1]徐丽华,娄恺,张华,等. 微生物资源学(第二版)[M].北京:科学出版社,2010:141,163-165,206.
[2]廉立慧,高丽君,王德才,等. 高温蛋白酶产生菌的筛选及其产酶条件和酶学性质分析[J]. 生物技术通报,2011,3:175-179.
[3]李士林,王宜君,汤朝起,等. 耐高温菌的分离及在固态发酵上部烟叶中的应用[J]. 生物加工工程,2015,13(1):35-41.
[4]张询,李晓云,孙丽娜. 污泥堆制发酵过程中多环麝香的降解及微生物群落的演替[J]. 沈阳农业大学学报,2015,46(1):96-100.
[5]李华芝,李秀艳,胡启平,等. 处理厨余垃圾的高温菌剂研制及其降解性质研究[J]. 华东师范大学学报(自然科学版),2011,2011(3):126-133.
[6]刘志恒. 现代微生物学(第二版)[M]. 北京:科学出版社,2008:406-410.
[7]孔雯,先锋,李长影,等. 1株烟碱降解菌的筛选、鉴定及其降解性能的初步研究[J]. 华中农业大学学报,2011,30(1):30-33.
[8]陈建军,吕永华,王维. 烟草品质生理及其调控研究[M].广州:华南理工大学出版社,2009:219-222.
[9]蒋咏梅. 微生物育种学实验[M]. 北京:科学出版社,2012:46-47.
[10]谢家仪,董光军,刘振英. 扫描电镜的微生物样品制备方法[J]. 电子显微学报,2005,24(4):440.
[11]范俐. 微生物学基础与实验技术[M]. 厦门:厦门大学出版社,2012:167-181.
[12]刘国生. 微生物学实验技术[M]. 北京:科学出版社,2007:141-155.
[13]陶天申,杨瑞馥,东秀珠. 原核生物系统学[M]. 北京:化学工业出版社,2007:375.
[14]陈瑜. 临床常见细菌、真菌鉴定手册[M]. 北京:人民卫生出版社,2009:190-193.
[15]Nazina T N, Tourova T P, Poltaraus A B, et al. Taxonomic study of aerobic thermophilic bacilli: descriptions of Geobacillus subterraneus gen[J]. International journal of systematic and evolutionary microbiology,2001,51:433-446.
[16]戴玄,唐兵,陈向东,等. 产高温蛋白酶微生物菌种资源的研究[J]. 微生物学杂志,1997,17(3):25-29.
[17]朱泓,王一明,林先贵. 响应面模型分析高温蛋白酶菌株增殖和产酶关系[J]. 微生物学通报,2014,41(5):1020-1027.
[18]陈泽斌,夏振远,雷丽萍,等. 烟草内生细菌种群特征分析[J]. 中国烟草学报,2014,20(3):102-107.
Isolation and identi fi cation of endophytic thermophilic bacteria YYFG3 from fl ue-cured tobacco and its protease activity
SUN Huizhong1, WANG Xiaodong1, ZHU Jinfeng2, LI Guangliang2, SUN Minghui2, XU Zhicheng3, SONG Yueqin1
1 College of Agriculture,Henan University of Science and Technology, Luoyang 471003, Henan, China;
2 Henan Luohe Municipal Tobacco Corporation, Luohe 462000 Henan, China;
3 College of Tobacco Science, Henan Agricultural University, Zhengzhou 450002, China
Thermophilic bacteria strain producing protease YYFG3 was isolated from fl ue-cured tobacco (Nicotiana tabacum L.) by using solid culture medium and milk agar medium. The growth temperature was in the range of 30-65℃, and the optimum growth temperature was 55℃ . The tolerance of pH was in the range of 5-9, and the optimum pH was 6-7. After 32h fermentation under the condition of 55℃ , 180 r/min, protease activity reached the maximum value of 35.3 U/mL. Therefore, the YYFG3 was identi fi ed as a thermophilic strain producing protease. Morphological characteristics of strain YYFG3 were observed by optical microscope and scanning electron microscope, and related biochemical properties were measured. 16S rDNA was cloned and sequenced, and its phylogenetic tree was established. YYFG3 was classi fi ed into the genus Geobacillus, and temporarily named Geobacillus sp. YYFG3. It was concluded that strain YYFG3 can be used as good material for conventional mutation breeding and genome breeding of thermophilic bacteria producing proteinase and enjoyed a good prospect for further development and application.
Nicotiana tabacum; endophytic bacteria; isolation; protease; 16S rDNA
孙会忠,王小东,朱金峰,等. 烤烟内生高温菌YYFG3的分离鉴定及所产蛋白酶活性[J]. 中国烟草学报,2015,21(6)
河南省烟草公司科技项目(HYKJ2012M04;HYKJ201302);上海烟草集团责任有限公司项目(SZBCW2014-00830);河南科技大学博士科研启动基金(4024-13480045;4026-13480047)
孙会忠(1976—),博士,副教授,主要从事植物内生菌研究,Email:huizhong66@163.com
朱金峰(1979—),硕士,高级农艺师,主要从事烟草生产技术研究与推广,Email:lylhsys@126.com
2015-03-21
:SUN Huizhong, WANG Xiaodong, ZHU Jinfeng, et al. Isolation and identi fi cation of endophytic thermophilic bacteria YYFG3 from fl ue-cured tobacco and its protease activity[J]. Acta Tabacaria Sinica, 2015,21(6 )
