苦草对水-底泥-沉水植物系统中氮素迁移转化的影响
2015-11-18孔祥龙李春华张来甲李永峰
孔祥龙,叶 春,李春华,张来甲,3,李永峰
(1.东北林业大学林学院,黑龙江 哈尔滨 150040;2.中国环境科学研究院,环境基准与风险评估国家重点实验室,湖泊工程技术中心,北京 100012;3.常州大学环境与安全工程学院,江苏 常州 213100)
苦草对水-底泥-沉水植物系统中氮素迁移转化的影响
孔祥龙1,2,叶 春2,李春华2,张来甲2,3,李永峰1*
(1.东北林业大学林学院,黑龙江 哈尔滨 150040;2.中国环境科学研究院,环境基准与风险评估国家重点实验室,湖泊工程技术中心,北京 100012;3.常州大学环境与安全工程学院,江苏 常州 213100)
通过模拟水-底泥-沉水植物(苦草)系统,检测了苦草整个生命周期内总氮及各形态氮含量的变化,以反映N在该系统内的迁移转换.结果表明,在整个研究阶段,空白组和苦草组系统氮含量(水体+底泥+苦草中氮含量)均持续降低,但苦草组(实验始末)氮含量降低幅度明显高于空白组,其中苦草组系统中TN、-N、-N含量分别减少了41.68%、81.96%、93.34%,分别比空白组提高了11.39%、31.90%、0.28%;苦草组底泥中TN、-N、-N含量分别减少了43.45%、87.41%、96.50%,分别比空白组提高了13.78%、37.26%、1.68%.苦草的存在促进了底泥氮的释放,显著提升了底泥微生物活性及氮循环菌的数量,从而加快了系统内的氮素循环,并在其生命周期的不同阶段明显改变各形态氮的迁移及转化方式.2012年7~10月,苦草组系统总氮(TN)减少幅度最大,到10月份,水体中氮素含量达到最少.
苦草;生命周期;氮素;迁移转化;水生植物
湖泊富营养化是世界范围内普遍存在的环境问题[1],其中氮循环的不平衡是导致湖泊富营养化的一个重要因素[2-3].近年来,恢复与重建沉水植物作为控制湖泊富营养化的重要途径,已经得到了广泛应用[4-5].但是由于沉水植物生长的周期性,其衰亡凋落的植物残体会腐烂分解,并释放出氮、磷等营养盐而对湖泊造成二次污染[6-8].因此,需要在沉水植物修复的过程中对其生物量进行调控.研究沉水植物生长周期的不同阶段对水-底泥-沉水植物系统氮素的迁移转化规律,确定最佳的沉水植物生物量及调控时机能为湖泊中沉水植物的生态管理提供基础资料.
苦草是我国最为常见的沉水植物之一,广泛分布于湖泊、河流等水域.在自然条件下,苦草的整个生命周期大致可分为萌芽期、快速生长期、稳定期、枯萎衰亡期及腐败分解期,每年3~4月份,苦草种子或者球茎开始萌发;6~7月份是苦草分蘖生长的快速期;9~10月份苦草稳定生长,生物量达到最大值;10月份以后,苦草逐步枯萎衰亡,至来年3~4月份,苦草迅速腐解,并向水体及底泥释放大量营养盐[7-8].目前国内外对于苦草净化富营养水体,修复污染环境方面的研究大多集中在苦草生命周期的某一个阶段(如生长期、腐解期等)或对系统的某一部分(如水体、沉积物或者泥水界面等),而系统研究苦草的整个生命周期对水环境的影响却很少[9-12].本研究从苦草种子培育开始,历经了春、夏、秋、冬再到次年春季这一完整的苦草生命周期,以水体-沉积物-沉水植物组成的系统为研究主体,全面探究沉水植物生命周期内该系统氮素的迁移转化规律,以期为湖泊中沉水植物的生态管理提供参考.
1 材料与方法
1.1 实验材料
本实验苦草种子取自宜兴市新庄街道农业生态园,在步入式模拟实验室内培育成苦草幼苗(株高约为5cm)作为实验用苦草(因为在自然条件下苦草种子的萌发率不易控制,直接在装置内播种,不能保证苦草的成活率,所以选择先在温室中利用苦草种子培育幼苗,再将幼苗移栽至实验装置中,移栽时选择基本大小一致的苦草幼苗,最大程度上保证实验组初始氮含量一致);底泥采自太湖竺山湾湖区,使用彼得逊采泥器采集,鲜泥经过冷冻干燥、去除粗粒及动植物残体、研磨过筛(100目筛)后,充分混匀作为实验用底泥;实验用水采用超纯水.
1.2 实验设计
实验装置选用圆柱形聚乙烯塑料桶(直径:15.00cm,高:50.00cm,预先经过5%的HCl 处理后用超纯水冲洗干净),30个.将充分混匀后的等量底泥(1647.20g)装入塑料桶内,泥厚约8cm,加入1L超纯水浸泡3d,以使底泥充分湿润,与水均匀混合;而后将培育好的苦草幼苗(株高5cm左右)以簇为单位移植至塑料桶内(每簇8株,共5簇),移植过程中先将苦草幼苗用超纯水洗净,避免外部营养盐的带入,移植完成后,向塑料桶内添加超纯水至刚没过苦草幼苗,稳定3d后加水至45cm处.共移植15桶苦草,剩余15桶作为空白对照,其加水流程与苦草组相同.苦草幼苗移植及装置加水工作于2012年4月2日完成.实验周期内每隔2周向实验装置添加超纯水,以保证装置内水平面稳定在45cm左右.实验在太湖研究基地内进行,实验装置置于一大玻璃棚罩下,该棚四周通透,棚顶透光性良好,且能避免降雨对实验造成的破坏.
1.3 取样与分析
2012年4月5日进行第一次采样,测定各项指标作为实验初始值,之后每隔3个月采集一次样品,至2013年4月,共采集5次样品,采样时间均为上午10时.每次采样均随机选取3桶苦草及3桶空白,将实验装置内的水、泥、苦草全部采出.采用虹吸法将上覆水吸出并精确计量其体积;苦草分离出底泥后用少量超纯水洗净植株上附着的底泥(洗净水倒入实验装置内);底泥采集后充分混匀,留取20g用于检测微生物及含水率,其他全部冷冻干燥,研磨过筛(100目筛)用于营养盐的测定.
数据用SPSS19.0软件进行线性回归分析,用Origin8.0作图.
2 结果与讨论
2.1 苦草生物量及株高变化
由图1可见,苦草生物量及株高均是先上升后下降,于2012年10月份取得最大值.从2012年4月~7月苦草快速生长,其生物量及株高分别增长了10.47倍和14.90倍.2012年7月~10月间苦草株高增速明显放缓,主要原因是由于实验装置内水体深度有限,限制了苦草株高的增长.2012年10月以后,苦草逐渐进入衰亡期,部分苦草被分解,其生物量相应减少,至2013年4月苦草生物量仅剩24.64g,其腐解率为67.20%.

图1 苦草生物量及株高的变化Fig.1 Changes of biomass and Plant height of submerged macrophytes
2.2 水体温度和光照的变化
温度是影响植物季节性生长的重要环境因子[16-17].实验周期内水温先上升后下降,分别在2012年7月及2013年1月取得最大值32.21℃和最小值7℃.另外,温度对沉水植物残体的腐解也有很大的影响[6-8,18].光照能够为植物光合作用提供必需的能源,是影响植物生长的重要环境因素之一[39].水环境中的光因子对沉水植物的生存、生长及分布有十分显著的影响[40],而大部分沉水植物的光补偿点均较低,约26~137lx[41];也有实验表明,在强光照和高营养盐水平下,苦草生长受到抑制[38].由图2可以看出,在实验周期内,光照强度随季节有较明显的变化,夏季光照强度(2012年7月)是冬季光照强度的(2013年1月)1.73倍.

图2 水体温度和光照强度的变化Fig.2 Changes of water temperature and light intensity
2.3 水体pH值及ρ(DO)的变化
由图3可见,苦草组水体pH值及ρ(DO)在实验周期内均先升高后下降,空白组水体pH值前期有所上升,2012年7月之后变化不大,空白组水体ρ(DO)一直较为稳定.实验开始的3个月内,温度开始升高,苦草幼苗迅速生长,其光合作用消耗了水体中的CO2,并释放出大量O2,促使水体pH值及ρ(DO)明显上升.2012年7~10月,苦草生长放缓,进入10月份后苦草逐渐开始衰亡,光合作用减弱,导致水体ρ(DO)略有下降.2012年10月至2013年1月间,苦草大量衰亡,其体内部分不稳定有机物进入水体[19],被微生物快速分解,并释放出CO2,造成水体pH值及ρ(DO)的显著下降;随着来年春天水温的升高,苦草残体大规模腐解,致使水体pH值及ρ(DO)继续下降.
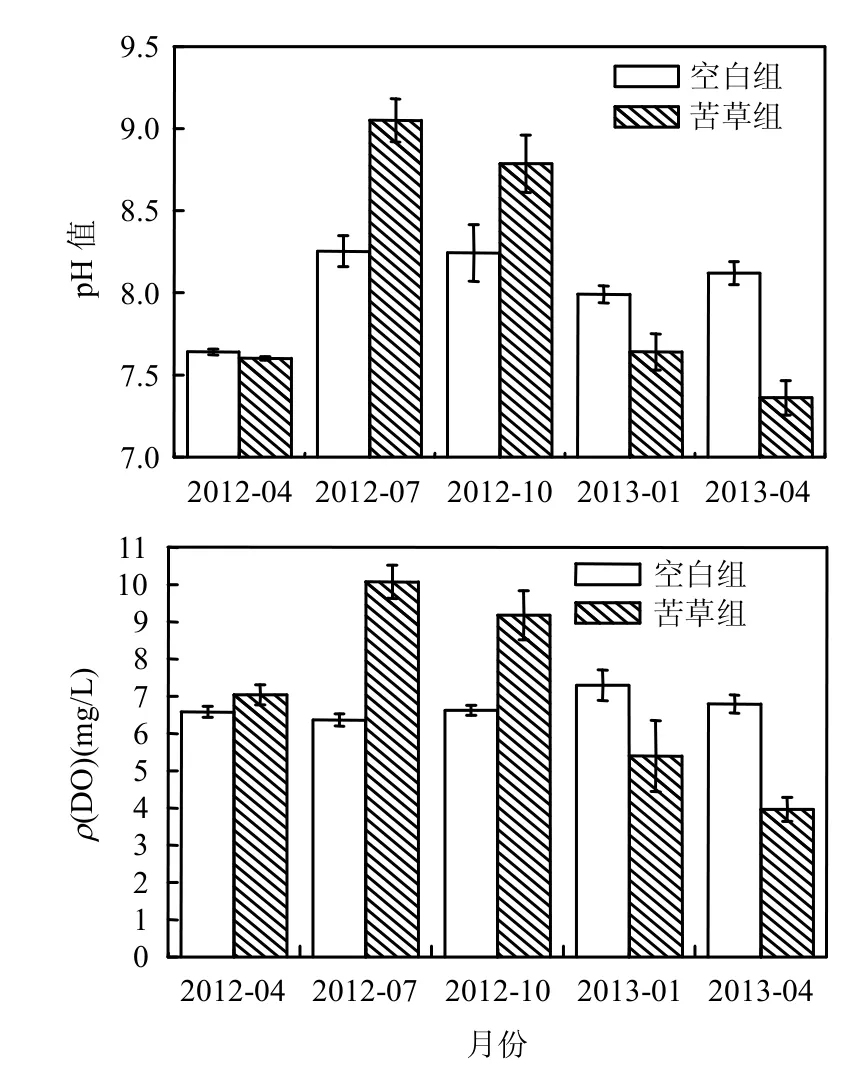
图3 水体pH值及ρ(DO)的变化Fig.3 Changes of water environmental physical factors
2.4 水体中叶绿素a含量的变化
由图4可见,实验周期内空白组水体叶绿素a的含量在实验初期有所增长,2012年10月份后基本维持稳定,苦草组水体叶绿素a的含量一直升高,2012年10月份后,苦草组水体叶绿素a的增量较大.
由于本实验用水为超纯水,所以实验开始时水体叶绿素a的含量为0.由于底泥中本身有少量残留的植物种子,在环境适宜情况下会萌发生长,进而导致空白组水体叶绿素a含量的增加,但增幅有限.苦草进入10月份后开始衰亡,苦草叶片断裂,造成苦草部分组织进入水体,引起水体叶绿素a的含量的升高;随着苦草残体被大规模分解,更多的植物组织进入水体,促使水体叶绿素a的含量不断增加.对前6个月(2012年4~10月)水体中叶绿素a的含量(Y)与实验时间(X)做线性回归分析,得出空白组:Y = 0.446X - 1.868,R² = 0.989;苦草组:Y = 0.78X - 3.053,R² = 0.998.可见,实验前6个月内水体中叶绿素a的含量与实验时间呈显著线性正相关.

图4 水体中叶绿素a含量的变化Fig.4 Change of Chl-a concentration in water
2.5 水-底泥-沉水植物系统中N的迁移转化
2.5.1 系统及各介质中TN含量的变化 苦草组系统氮含量由水体、底泥及沉水植物中的氮含量组成,空白组系统氮含量由水体及底泥中的氮含量组成.由图5a可以看出,苦草组及空白组系统TN含量在实验周期内一直在降低,截止实验结束,空白组TN较起始降低了30.29%,苦草组降低了41.68%,比空白组提高11.39%,说明苦草的存在促进了系统内氮的去除.2012年4~7月,空白组系统TN含量减少了12.80%,而苦草组系统降幅仅有8.66%,这主要是由于该阶段苦草快速生长,水体及底泥中的无机氮被苦草大量吸收并同化,并以有机氮的形式储存于植物体内,所以系统TN降幅小于空白组;2012年7月~10月的时间段,苦草组TN减少量最大,此阶段,苦草稳定生长,叶片变浓绿、宽大,其生物量及株高达到最大,由于白天强烈的光合作用,使系统DO过饱和,系统内有机氮迅速矿化,夜晚由于强烈的呼吸作用使系统内DO迅速下降,系统反硝化反应剧烈,这种由于沉水植物造成的氧化还原环境的变化,提高了系统TN的去除效率.2012年4~10月,苦草生物量及株高持续增长(图1),植物生长吸收水体及底泥中的氮素,使水体TN含量(图5c))持续降低,到2012年10月份,水体中总氮含量达到了最低值,2013年1月,苦草将衰亡腐解,并向水体释放大量营养盐,导致水体氮含量升高,如果在2012年10月份将部分苦草移出系统,既可以达到净化水体的目的,又能降低秋冬季节苦草的衰亡腐解对水体产生的二次污染.另外,虽然2012年10月份苦草的生物量及株高(图1)均达到了最大值,但是其含氮量却低于2012年7月份,计算2012年7月和10月苦草的单位质量含氮量分别为28.51、20.70mg/g,导致这一现象的原因是苦草从10月份开始逐步进入枯萎衰亡期,其体内部分有机物会游离出苦草个体,进入其他介质[19],Cisternas[20]的研究表明,叶片凋落物N减少的原因主要有3个:最初是死叶片的N滤去,然后是固定N细菌或真菌的迁移作用;最后是由于反硝化细菌、负责分解的中间动物群、氨挥发和淋溶的作用;2012年10月~2013年1月,苦草组及空白组系统TN含量减少率分别为6.14%、0.16%,较前一时期,减少率明显放缓.其主要原因是随着温度的降低,微生物活性及氮循环菌数量均随之降低(图7),氮循环速率减缓;2013年1月~4月,苦草开始大量腐解,导致水体含氮量明显升高.对从2012年10月~2013年4月,植物TN(Y1)及水体TN(Y2)与苦草生物量(X)进行先行回归分析,得出线性回归方程分别为:Y1= 0.75X - 7.447,R² = 0.999,Y2= -0.437X + 29.093,R² = 0.911,说明植物腐解期,生物量与植物TN含量显著正相关,与水体TN含量负相关.
2.5.2 系统及各介质中形态氮含量的变化 各形态氮可在植物及微生物的作用下相互转化,而这种转化是N在各介质之间迁移的主要驱动力[21-22].另外,氨挥发、N沉降、底泥粘粒的吸附和解析等方式也促进了N在各介质间的迁移.由图6a可以看出,空白组系统内含量在实验周期内一直减少,苦草组系统在实验周期内一直降低,而含量在2013年1月之前一直减少,2013年1月~4月有小幅增加.总体来看,苦草组系统减少量明显高于空白组无显著差别.
2012年4月~7月,温度逐渐升高,苦草快速生长,底泥中不断有氮素释放出来,一部分在氮循环菌作用下以气体形式迁出系统,另一部分进入水体或被植物同化进入植物体内.水体及底泥中的-N和-N均减少.2012年7月~10月,温度较高,微生物活性剂及数量均达到最大,底泥中氮素的矿化速率也最大,同时反硝化菌n(MPN)也达到最大(图7d),氮素迁出系统的速率最高,底泥中-N和底泥及水体中-N均有所减少,水体中略有增加是由底泥中氮素矿化生成的-N释放到水体中所致,而此时的硝化菌n(MPN)比氨化菌n(MPN)及反硝化菌n(MPN)低2~3个数量级(图7)也能解释这一现象.2012年10月~2013年1月,温度逐渐降低,微生物活性也逐渐减弱,底泥和水体中的-N均没有明显变化,说明此阶段-N的产生与消耗处于平衡阶段,而与空白组相比,苦草组底泥的矿化速率扔明显高于空白组.2013年1~4月苦草大量腐解,向水体释放大量的营养盐,导致水体中-N及-N迅速升高,部分迁移至底泥中,进而底泥中-N含量有所提高,而-N的减少数量与空白组相比也有所降低.

图6 系统及各介质中形态氮含量的变化Fig.6 Changes of-N and-N content in the system, sediment, water and plant
2.5.3 底泥中微生物活性及氮循环菌数量的变化 电子受体活性(ETS)主要是通过测定微生物细胞内脱氢酶(DHA)的活性来间接的反应微生物群落的活性[23-24].实验初期,由于底泥经过冷冻干燥,其微生物大部分处于休眠状态,活性较低;随着温度升高,苦草组及空白组微生物在2012年7~10月均处于较高活性,且苦草组微生物活性明显高于空白组;进入秋冬季节以后,苦草组及空白组活性明显减弱,主要是1月份水温较低,微生物活性受到抑制[25],至来年春季,底泥微生物活性又有所回升.苦草组及空白组氨化菌n(MPN)在2012年4~7月有明显的增长,苦草组增长了近5个数量级;随后氨化菌n(MPN)较为稳定;2013年1月有所下降,而后在4月份又逐步回升.硝化菌n(MPN)及反硝化菌n(MPN)的变化趋势与氨化菌n(MPN)基本类似,但是反硝化菌n(MPN)是在2012年10月份取得最大值,而且,硝化菌n(MPN)在实验周期内偏少,较氨化菌n(MPN)及反硝化菌n(MPN)小了2~3个数量级.
氮的迁移转化离不开氮循环菌群的作用[26],相关研究表明,氮循环微生物对外界环境的要求不一[25],由于敞水区水质及底泥的匀质性,氮循环微生物很难同时形成优势种群并发挥作用[27];水生高等植物的存在能够在其根部形成“根际氧化带”,又由于植物的覆盖及其呼吸代谢作用,氧化带之外往往处于缺氧状态,从而在水生高等植物根区形成了这样一个富氧-缺氧的氧化还原微环境,因而,反硝化菌、硝化菌、氨化细菌等微生物能够同时出现并发挥其作用[27-29].空白组和苦草组氮循环菌的变化规律明显,两系统均在2012年7月~10月份氮循环菌达到较大值,到2013年一月份达到最低,然后到2013年4月份有所回升,但苦草组氮循环菌的数量明显高于空白组,说明苦草的存在促进了氮循环菌群的繁殖,进而促进了氮的迁移转化的力度与速率.在苦草生长周期的不同阶段,氮循环菌群处于不同的动态平衡状态,该状态随温度、DO、pH值及植物生长状况等外界条件不断发生改变,从而导致各形态氮在水、底泥及植物之间的相互迁移转化.

图7 底泥中微生物活性及氮循环菌数量的变化Fig.7 Changes of activity and amount of nitrogen cycle bacteria in sediment
2.5.4 底泥及水体中各形态氮比例的变化 由图8a,图8b可以看出,空白组和苦草组底泥中不同形态氮含量的顺序为有机氮>NH4+-N> NO3--N,即使沉水植物的存在也未能改变这种格局.空白组和苦草组底泥中NO3--N含量总体上来说一直相对较低,所占比例变化不大,主要变化体现在2012年7月份,此时温度较高,菌活性较强,底泥中NO3--N释放较快;空白组底泥NH4+-N比例在实验周期内变化不大,说明空白组底泥中的NH4+-N的生成与消耗处于动态平衡状态,苦草组底泥NH4+-N比例在实验前期(2012年4月~2013年1月)一直降低,是因为底泥通过矿化进入上层水中的-N含量较高[30-31],2013年1~4月,苦草组底泥中-N比例有所回升,是因为初春植物腐解向水体中释放大量的氮素,由于底泥中无机颗粒对-N具有较强的吸附作用[32],使得底泥中-N含量升高.这也证明了苦草的存在打破了空白组状态下的动态平衡,明显改变了-N的迁移及转化.
水生植物主要通过影响pH值和DO等水质参数[33],吸收营养盐[34-35],为浮游动物提供栖息地[34],分泌抑制藻类生长的物质[34]和改变底泥理化性质[36]等而影响水环境,改变水体水质.空白组水体(图8c)中由于没有植物,无机氮不能被吸收,有机氮则不断被分解,只是有机氮比例逐步降低,同时,空白组水体ρ(DO)一直处于较高水平,为硝化作用提供了条件,所以空白组水体中-N比例稳步增加.苦草组水体(图8d)各赋存形态的氮所占比例均变化较大,这也表明,沉水植物的存在影响了各赋存形态氮的分配比例,且这种影响在其不同的生长阶段不同.

图8 底泥及水体中各赋存形式氮的比例变化Fig.8 Changes of the proportion of various forms of nitrogen in the sediment and water
实验周期内,空白组底泥中各形态氮所占比例基本变化不大,说明底泥中各形态氮等比减少,整个阶段处于平衡状态.释放到水体中各赋存形式的氮,受环境温度、pH值、DO、光照及微生物等因素的影响,不断发生变化,趋向于平衡.苦草组由于苦草的加入,打破了原有平衡.在2012年4月~7月,底泥和水体中-N所占比例均不断降低,关于利用水生物作为有机碳源,降低水体硝酸盐含量的报道[37]证实了反硝化作用强度与有机碳源的供给密切相关.底泥中-N比例不断下降,而水体中却不断上升,说明该阶段底泥中氮素不断矿化,并以-N形式不断释放到水体中.在2012年10月~2013年4月,苦草经历了衰亡期和腐解期,有研究[18]认为沉水植物在秋、冬寒冷季节虽然进入衰亡期,但低温环境下植物体腐解比较缓慢,营养物质释放量也较为有限,大量的营养物质保留在植物残体中,短时间内不会对水质产生显著影响,待翌年初春天天气回暖,才向水体大规模释放营养物质.2012年10月~2013年1月份,水体和底泥中-N比例基本无变化-N比例下降,有机氮比例升高,这是因为此阶段温度较低,氨化菌n(MPN)减少,其活性变弱,有机氮的矿化作用减少.2013年1~4月,由于温度回升,苦草开始大量腐解,导致水体中-N和-N含量增加,图8d中-N比例并没有增加是因为底泥对水体中-N的强吸收作用[32],使水体中-N比例没有增加而底泥中-N比例升高.
2.6 底泥中C/N的变化
为更深入的分析实验周期内N的变化,计算底泥中TOC和TN的比值作图9.试验周期内,空白组底泥C/N比略有波动,但基本稳定在8~10之间,苦草组底泥C/N一直减小,在2013年1~4月的时间段内降幅最大,达到39.27%.相关研究表明[42-44],底泥中C/N比的大小对土壤中有机氮的矿化和矿质氮微生物的固持2个过程有较为明显的影响.当C/N> 30时,矿质氮的生物固持作用大于有机氮的矿化作用,表现为矿质氮的净生物固持;当C/N <20时,有机氮的矿化作用大于矿质氮的生物固持作用,表现为有机氮的净矿化;当C/N比处于20~30之间时,两个过程强度均等,既不表现矿质氮的净生物固持也不表现有机氮的净矿化.同时,当C/N<14时,植物才能从外界环境大量吸收氮.实验周期内,空白组和苦草组底泥中的C/N比一直小于20,最大值仅为10.22.这时的C/N比使底泥中的氮主要表现为有机氮的矿化.这一点从图7b底泥中氨化菌n(MPN)的变化上得到了验证,氨化菌n(MPN)在实验周期内均处于高水平.而由于底泥中的溶解氧较低,反硝化作用明显(图7d),导致空白组及苦草组底泥TOC及TN含量持续减少.其中,苦草组微生物活性较高,促使其系统TOC及TN的减少量较大;另外,底泥中的C/N比,十分有利于苦草从底泥中吸收营养盐,这也加剧了底泥中TOC及TN含量的减少.

图9 底泥中TOC/TN比的变化Fig.9 Changes of TOC/TN in sediment
3 结论
3.1 苦草的存在明显加强了“水-底泥-沉水植物”系统氮素的迁移转化,至实验结束,系统TN、含量共减少了41.68%、81.96%、 93.34%,分别比空白组提高了11.39%、31.90、 0.28%;苦草组底泥含量分别减少了43.45%、87.41%、96.5%,分别比空白组提高了13.78%、37.26%、1.68%.
3.2 苦草的存在明显改变了底泥电子受体活性及氮循环菌的数量,2012年7月~10月,电子受体活性及各氮循环菌数量均比空白组升高,促进了氮的迁移转化的力度与速率,明显改变形态氮的迁移及转化方向.
3.3 苦草整个生命周期内,系统TN含量持续降低,并没有因为苦草存在而打破这一格局,但在不同阶段苦草的存在能改变形态氮的迁移方向和迁移速率.2012年4月~7月,苦草快速生长,系统内部分氮素被苦草同化而进入苦草体内,与空白组相比,移出系统的氮减少;2012年7月~10月,温度较高,微生物活性及数量均达到最大值,苦草组系统氮减少量最大,2012年10月,水体中总氮含量最低;2012年10月~2013年4月,苦草经历衰亡和腐解期,苦草中的氮素进入水体,部分迁移至底泥中.
[1]Walter K D, Wes W B, Jeffrey L E, et al. Eutrophication of U. S. Freshwaters: Analysis of Potential Economic Damages [J]. Environmental Science and Technology, 2009,43(1):12-19.
[2]许 海,朱广伟,秦伯强,等.氮磷比对水华蓝藻优势形成的影响[J]. 中国环境科学, 2011,31(10):1676-1683.
[3]秦伯强,杨柳燕,陈非洲,等.湖泊富营养化发生机制与控制技术及其应用 [J]. 科学通报, 2006,51(16):1858-1866.
[4]张来甲,叶 春,李春华,等.不同生物量苦草在生命周期的不同阶段对水体水质的影响 [J]. 中国环境科学, 2013,33(11):2053-2061.
[5]Sabine H, Elisabeth M G, Michael H, et al. Restoration of submerged vegetation in shallow eutrophic lakes - A guideline and state of the art in Germany [J]. Limnologica-Ecology and Management of Inland waters, 2006,36(3):155-171.
[6]叶 春,王 博.沉水植物黑藻早期分解过程及影响因素研究[J]. 中国农学通报, 2009,25(17):5-10.
[7]王 博,叶 春,李春华,等.初春苦草腐解过程中营养盐释放过程及规律 [J]. 生态与农村环境学报, 2012,28(2):171-175.
[8]张来甲,叶 春,李春华,等.沉水植物腐解对水体水质的影响[J]. 环境科学研究, 2013,26(2):146-151.
[9]王立志,王国祥,俞振飞,等.苦草(Vallisneria natans)生长期对沉积物磷形态及迁移的影响 [J]. 湖泊科学, 2011,23(5):753-760.
[10]宋 睿,姜锦林,耿金菊,等.不同浓度铵态氮对苦草的生理影响[J]. 中国环科科学, 2011,31(3):448-453.
[11]Xie Yonghong, An Shuqing, Yao Xin, et al. Short-time response in root morphology of Vallisneria natans to sediment type and water-column nutrient [J]. Aquatic Botany, 2005,81(1):85-96.
[12]Wang Chao, Lu Jie, Zhang Songhe, et al. Effects of Pb stress on nutrient uptake and secondary metabolism in submerged macrophyte Vallisneria natans [J]. Ecotoxicology and Environmental Safety, 2011,74(5):1297-1303.
[13]国家环境保护总局《水和废水监测分析方法》编委会.水和废水监测分析方法 [M]. 北京:中国环境科学出版社, 2002:243-284.
[14]林先贵.土壤微生物研究原理与方法 [M]. 北京:高等教育出版社, 2009:39-60.
[15]鲁如坤.土壤农业化学分析方法 [M]. 北京:中国农业科技出版社, 2000:146-163.
[16]Vera I, Márk H, Ádám K, et al. Distribution of submerged macrophytes along environmental gradients in large, shallow Lake Balaton [J]. Aquatic Botany, 2008,88(4):317-330.
[17]Ye Chun, Yu Haichan, Kong Hainan, et al. Community collocation of four submerged macrophytes on two kinds of sediments in Lake Taihu, China [J]. Ecological Engineering, 2009,35(11):1656-1663.
[18]潘慧云,徐小花,高士祥.沉水植物衰亡过程中营养盐的释放过程及规律 [J]. 环境科学研究, 2008,21(1):64-68.
[19]吴振斌,邱东茹,贺 锋,等.沉水植物重建对富营养水体氮磷营养水平的影响 [J]. 应用生态学报, 2003,14(8):1351-1353.
[20]Cisternas R E, Yates L R. Nitrogen in litterfall and precipitation and its release during litter decomposition in the Chilean piedmont matorral [J]. Developments in Plant and Soil Sciences,1982,67(1-3):359-365.
[21]唐国勇,黄道友,童成立,等.土壤氮素循环模型及其模拟研究进[J]. 应用生态学报, 2005,16(11):2208-2210.
[22]Burns D A, Murdoch P. Effects of a clear cut on the net rates of nitrification and N mineralization in a northern hardwood forest,Catskill Mountains, New York, USA [J]. Biogeochemistry, 2005,72:123-146.
[23]Li Chunhua, Ye Chun, Wong Yukshan, et al. Effect of Mn (IV)on the biodegradation of polycyclic aromatic hydrocarbons under low-oxygen condition in mangrove sediment slurry [J]. Journal of Hazardous Materials, 2011,190(1-3):786-793.
[24]Li Feng, Zhu Lizhong. Effect of surfactant-induced cell surface modifications on electron transport system and catechol 1,2-dioxygenase activities and phenanthrene biodegradation by Citrobacter sp. SA01 [J]. Bioresource Technology, 2012,123:42-48.
[25]周群英,高廷耀.环境工程微生物学(第二版) [M]. 北京:高等教育出版社, 1999,207-213.
[26]刘东山,罗启芳.东湖氮循环细菌分布及其作用 [J]. 环境科学,2002,23(3):29-35.
[27]王国祥,濮培民,黄 宜,等.太湖反硝化、硝化、亚硝化及氨化细菌分布及其作用 [J]. 应用与环境生物学报, 1998,5(2):190-194.
[28]安 慧,上官周平.植物氮素循环过程及其根域调控机制 [J].水土保持研究, 2006,13(1):84-88.
[29]金相灿,江 霞,徐玉慧,等.太湖东北部沉积物可溶性氮、磷的季节性变化 [J]. 中国环境科学, 2006,26(4):409-413.
[30]McCarthy M J, Gardner W S, Lavrentyev P J et al. Effects of Hydrological Flow Regime on Sediment-water Interface and Water Column Nitrogen Dynamics in a Great Lakes Coastal Wetland (Old Woman Creek, Lake Erie) [J]. Journal of Great Lakes Research, 2007,33(1):210-231.
[31]Alvarez N O, Caetano M, Vale C, et al. Exchange of nutrients across the sediment-water interface in intertidal ria systems (SW Europe) [J]. Journal of Sea Research, 2014,85:349-358.
[32]Xia Xinghui, Liu Ting, Yang Zhifeng, et al. Dissolved organic nitrogen transformation in river water: Effects of suspended sediment and organic nitrogen concentration [J]. Journal of Hydrology, 2013,484:96-104.
[33]Willian J J. The role of water plant in water treatment [J].Agricultural Engeneering, 1986,57(6):9-10.
[34]Bailey P, Boon P, Morris K. Managing nutrients in flood plain wetlands and shallow lakes: 2river and riparian land management technical guideline update [M]. Canberra: Land and Water Australia, 2002.
[35]徐红灯,席北斗,王京刚,等.水生植物对农田排水沟渠中氮、磷的截留效应 [J]. 环境科学研究, 2007,20(2):84-88.
[36]Qiu D R, Wu Z B, Liu B Y. et al. The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lakein Hubei Province, China [J]. Ecological Engineering,2001,18:147-156.
[37]Bastviken S K, Eriksson P G. Potential Denitrification in Wetland Sediments With Different PlantSpecies Detritus [J]. Ecological Engineering, 2005,25(2):183-190.
[38]黎慧娟,倪乐意,曹 特,等.弱光照和富营养对苦草生长的影响[J]. 水生生物学报, 2008,32(2):225-230.
[39]王 华,逄 勇,刘申宝,等.沉水植物生长影响因子研究进展[J]. 生态学报, 2008,28(8):3958-3968.
[40]Pérez G L, Torremorell A, Bustingorry J, et al. Optical characteristics of shallow lakes from the Pampa and Patagonia regions of Argentina [J]. Limnologica Ecology and Management of Inland Waters, 2010,40(1):30-39.
[41]Brenchley J L, Probert R J. Seed germination responses to some environmental factors in the seagrass Zostera capricorni from eastern Australia [J]. Aquatic Botany, 1998,62,177-188.
[42]鲁彩艳,陈 欣.有机碳源添加对不同C/N比有机物料氮矿化进程的影响 [J]. 中国科学院研究生院学报, 2004,21(1):109-112.
[43]鲁彩艳,陈 欣.不同施肥处理土壤及不同C/N比有机物料中有机N的矿化进程 [J]. 土壤通报, 2003,34(4):268-270.
[44]Larsen K L, McCartney D M. Effect of C: N ratio on microbial activity and N retention: bench-scale study using pulp and paper biosolids [J]. Biosolids1Compost Science and Utilization, 2000,8(2):147-159.
Effect on nitrogen transfer and migration by Vallisneria natans (Lour.) Hara in water-sediment-submerged macrophytes system.
KONG Xiang-long1,2, YE Chun2, LI Chun-hua2, ZHANG Lai-jia2,3, LI Yong-feng1*
(1.School of Forestry, Northeast Forestry University, Harbin 150040, China;2.State Key Laboratory of Environmental Criteria and Risk Assessment, Centre of Lake Engineering and Technology, Chinese Research Academy of Environmental Sciences,Beijing 100012, China;3.School of Environment and Safety Engineering, Changzhou University, Changzhou 213100,China).
China Environmental Science, 2015,35(2):539~549
The changes of nitrogen in water, sediment and submerged macrophytes (Vallisneria natans (Lour.) Hara) as well as its migration and transformation in the life cycle of V. natans were studied. The results indicated that the total nitrogen content was reducing continuously in V. natans and control treatments during the research stage, and the removing rate of total nitrogen in V. natans treatments was obviously higher than that in control treatments. The content of TN、-N、-N in the water-sediment-submerged macrophytes system decreased by 41.68%, 81.96% , 93.34%, and the removal rates increased by 11.39%, 31.90, 0.28% than that in control treatment, respectively. In this system, the content of TN-N,-N in sediment decreased by 43.45%, 87.41%, 96.50%, and the removal rates are increased 13.78%, 37.26%, 1.68% than that in control treatment, respectively. V. natans contributed to nitrogen nutrients release in the sediment. The sediment microorganisms activity and the amount of nitrogen cycling bacteria were improved significantly. Therefore, the nitrogen cycle speed would be accelerated. The direction of transfer and transformation of nitrogen would be changed in the different stage of life cycle of V. natans. Compared with other stages, the nitrogen content in experiment group with V. natans decreased most during July to October of 2012. To October, the nitrogen content in water reached a minimum value.
Vallisneria natans;lifecycle;nitrogen;transfer and transformation;aquatic plant
X171
A
1000-6923(2015)02-0539-11
孔祥龙(1987-),男,山东日照人,东北林业大学林学院硕士研究生,主要从事湖泊富营养化治理和生态恢复理论与技术研究.
2014-6-10
国家自然科学基金资助项目(40971277);国家水体污染控制与治理科技重大专项(2012ZX07101-009)
* 责任作者, 教授, dr_lyf@163.com
