Effects of cultivation of OsrHSA transgenic rice on functional diversity of microbial communities in the soil rhizosphere
2015-11-12XiobingZhngXujingWngQiolingTngNingLiPeileiLiuYufengDongWeiminPngJingtoYngZhixingWng
Xiobing Zhng,Xujing Wng*,Qioling TngNing Li,Peilei Liu,Yufeng DongWeimin PngJingto YngZhixing Wng*
aBiotechnology Research Institute,Chinese Academy of Agricultural Sciences,Beijing 100081,China
bBiology Institute,Hebei Academy of Sciences,Shijiazhuang 050051,China
cDevelopment Center of Science and Technology,Ministry of Agriculture,Beijing 100122,China
Effects of cultivation of OsrHSA transgenic rice on functional diversity of microbial communities in the soil rhizosphere
Xiaobing Zhanga,b,Xujing Wanga,*,Qiaoling Tanga,Ning Lic,Peilei Liuc,Yufeng Donga,Weimin Panga,Jiangtao Yanga,Zhixing Wanga,*
aBiotechnology Research Institute,Chinese Academy of Agricultural Sciences,Beijing 100081,China
bBiology Institute,Hebei Academy of Sciences,Shijiazhuang 050051,China
cDevelopment Center of Science and Technology,Ministry of Agriculture,Beijing 100122,China
A R T I C L E I N F O
Article history:
13 November 2014
Accepted 4 January 2015
Available online 13 January 2015
Biolog method
ELISA
Soil microbes
Functional diversity
With the widespread cultivation of transgenic crops,there is increasing concern about unintended effects of these crops on soil environmental quality.In this study,we used the Biolog method and ELISA to evaluate the possible effects of OsrHSA transgenic rice on soil microbial utilization of carbon substrates under field conditions.There were no significant differences in average well-color development(AWCD)values,Shannon-Wiener diversity index(H),Simpson dominance indices(D)and Shannon-Wiener evenness indices(E)of microbial communities inrhizosphere soils at eight samplings between OsrHSA transgenic rice and its non-transgenic counterpart.The main carbon sources utilized by soil microbes were carbohydrates,carboxylic acids,amino acids and polymers.The types,capacities and patterns of carbon source utilization by microbial communities in rhizosphere soils were similar throughout the detection period.We detected no OsrHSA protein in the roots of OsrHSA transgenic rice.We concluded that OsrHSA transgenic rice and the rHSA protein it produced did not alter the functional diversity of microbial communities in the rhizosphere.
©2015 Production and hosting by Elsevier B.V.on behalf of Crop Science Society of China and Institute of Crop Science,CAAS.This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
1.Introduction
The development of transgenic crops has brought huge economic benefits along with concerns of ecological safety,such as possible effects on soil organisms.Numerous studies have been carried out to investigate the effects of transgenic crops on soil microbial communities[1-3].Most results indicate that transgenic Bt crops have not caused significant effects on soil microbes compared to conventional crops[4-10].OsrHSA transgenic rice is transformed with the human serum albumin gene that produces a soluble medicinal globular protein[11]that has attracted attention regarding its environmental safety.However,few studies have examined the effects of OsrHSA transgenic rice on soil microorganisms.The aim of this study was to evaluate the effect of OsrHSA transgenic rice on soil microbial functions under field conditions and to provide a scientific basis for evaluating the safety of OsrHSA transgenic rice in China.

Fig.1-The standard curve of protein determination using the BCA method.
2.Materials and methods
2.1.Sampling and pretreatment
The study carried out using OsrHSA transgenic rice and its non-transgenic parent cultivar TP309 as a control,was conducted at an experimental farm located in Xiantao,Hubei,China(30°N,113°E).The soil type was paddy(pH 7.8,total N 2.24 g kg-1,total P 0.61 g kg-1,total K 18.2 g kg-1;organic matter 28.4 g kg-1).All treatments were set up in a completely randomized design with three replicates.
Soil samples collected from five points in an S-shaped curve across each field were taken from depths of approximately 10 cm,along with as much associated root material as possible.Sampling occurred at the tillering,jointing,milk and mature stages,and at 83,204,261,and 321 days after the rice was harvested.The soils were placed into plastic bags,which were placed in ice cube boxes and immediately taken to the laboratory,where they were stored in a refrigerator at 4°C and subsequentlyusedfordeterminationofsoilmicrobial indicators.
2.2.Biolog cultivation and statistical analysis
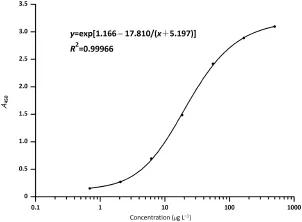
Fig.2-The standard curve of protein determination by ELISA.
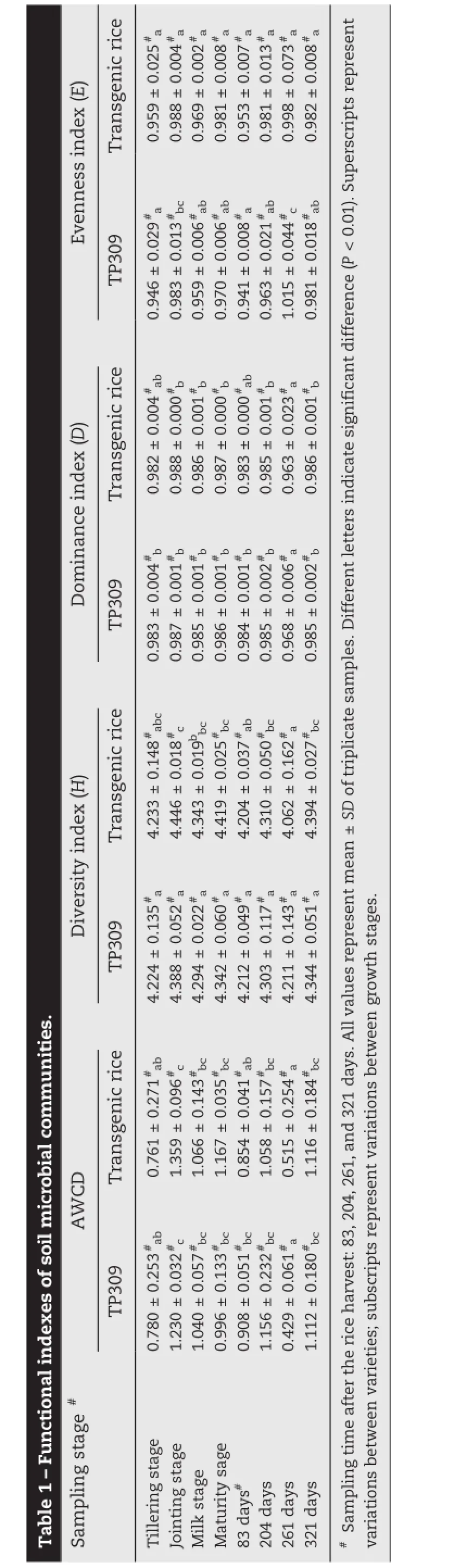
Table 2-Loading factors of principle components of 31 types of carbon sources.
The rice rhizosphere(root system)was shaken vigorously to remove large soil particles and the rhizosphere soil wascollected from the root surfaces.Fresh 2 g soil samples were placed in 20 mL of sterile physiological saline solution,sealed with sealing film and shaken.The soil suspensions were stepwise diluted to 1000 timesand added toBiolog Eco microplates(Biolog Co.,CITY,USA).The inoculated microplates were incubated at 28°C.Color developmentinthewellswasmeasured astheabsorbancevalue withamicroplatereader(Bio-TekInstrumentsInc.,USA)after24,48,72,96,120,144,and 168 h.
Following inspection of the data the absorbance values at 72 h were subjected to statistical analysis.An overall microbial activity indicator was estimated by average well-color development(AWCD)value,which reflected the capacity of carbon source utilization and metabolic activity of soil microbes.It was calculated as follows:

where C indicates the absorbance values of media of each well;R indicates the absorbance values of the control well;n indicates the number of carbon source types in the Biolog Eco microplate.
The Shannon-Wiener diversity index(H),Simpson dominance index(D)and Shannon-Wiener evenness index(E)were used to assess the richness,dominating population and evenness of soil microorganisms,respectively.These indices were calculated using the formulae:

where Piindicates the ratio of the relative absorbance value in the ith well to the total relative absorbance values in the entire microplate.

where H indicates the Shannon-Wiener diversity index and S indicates the number of wells with color variation.

Table 3-Score of principal variables affecting PC1 and PC2.
Variance analysis was performed using SPSS(SPSS for Windows,Version 16.0,USA).
2.3.Extraction and quantification of OsrHSA protein
Three grams of roots,stems,leaves and spikes(fresh weight equivalent)were ground.Aliquots of approximately 400 mg of crushed tissue samples were separately mixed with 1.0 mL proteinextractionreagent(PlantProteinExtractionKit,Beijing Cowin Bioscience Co.,Ltd.,China).The suspensions were centrifuged,and the clarified supernatants were transferred to new tubes.
Total soluble protein concentration(Fig.1)was measured with a BCA Protein Assay Kit(Beijing Cowin Bioscience Co.,Ltd.,China)and calculated as follows:

OsrHSA protein(Fig.2)was quantified using a Human Albumin ELISA Quantitation Set(Bethyl Laboratories,Inc.,USA)and calculated as:

3.Results and discussion
The Biolog system is an effective approach that is commonly employed to assess spatial and temporal changes in microbial communities.Higher AWCD values indicate higher metabolic activity by microbial communities.Higher H,D,andE suggested higher functional diversity of soil microbes and more uniform distribution of individuals among species.As shown in Table 1 differences in the AWCD,H,D,and E of microbial communities between OsrHSA transgenic rice and its non-transgenic rice TP309 counterpart were not significant at any observation point.
The 31 carbon source substrates on the microplate were divided into six categories:carbohydrates,amino acids,carboxylicacids,polymers,aminesandmetabolismproducts.Theinitial loading factor reflects the correlation coefficient between principal components and carbon utilization.As shown in Table 2,25 carbon sources used by soil microbes on the transgenic rice had relatively high correlation with PC1,20 substrates used by soil microbes on TP309 had relatively high correlation with PC1. According to the scores of variables affecting PC1 and PC2(Table 3),the main carbon sources utilized by soil microbes on OsrHSA transgenic and non-transgenic rice are carbohydrates,carboxylic acids,amino acids and polymers.
The principle behind principal component extraction is to extract the first m principal components with corresponding eigenvalues greater than one.Based on this concept,a total of seven principal components were extracted,with a cumulative contribution rate of 86.82%.Specifically,the contribution rate of PC1 was 65.07%,accounting for the greatest proportion,and the contribution rate of PC2 was 7.00%.Consequently,PC1 as the horizontal axis and PC2 as the vertical axis were employed to construct a principal component analysis diagram of carbon utilization,and the scores of the two principal components were used as the coordinates(Fig.3).As shown in Fig.3,On the PC1 axis,both OsrHSA transgenic rice and its non-transgenic counterpart were distributed in the positive range for all eight stages.On the PC2 axis,they were distributed in the positive direction as well.These results demonstrate similar capacities and patterns of carbon source utilization by microbial communities on both genotypes.
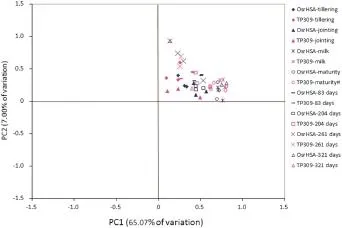
Fig.3-Principal component analysis of carbon utilization by soil microbial communities in different treatment groups.
There are numerous factors that affect evaluations of the safety of transgenic plants,such as temperature,soil type,exogenous chemicals and plant species.Liu et al.[12]and Lee et al.[13]reported differences associated with the plant growth stage.We determined that significant variations were associated with different sampling times;these might be due to seasonal or temperature fluctuations.
We quantified OsrHSA protein using ELISA,which allowed a threshold detection of 6.25 ng mL-1.OsrHSA protein was detected only in the spikes of transgenic rice at the milk and maturity stages(Fig.4).Although a strong endosperm-specific promoter and its signal peptide wereusedinthe transgenic line OsrHSA protein was expressed only in the endosperm[11].In this study,OsrHSA protein was not detected in transgenic rice roots.Moreover,protein released into soil cannot accumulate[9].Therefore,OsrHSA protein had no effect on the functional diversity of soil microorganisms.

Fig.4-OsrHSA and total protein in rice.All values refer to fresh tissue weight and represent means±SD of triplicate samples.*Indicates“below detection limit”.
Acknowledgments
This work was supported by the Major Project of China on New Varieties of GMO Cultivation(2012ZX08011-003 and 2014ZX08011-004B).We thank Prof.Daichang Yang(College of Life Sciences,Wuhan University,China)for providing test specimens.
R E F E R E N C E S
[1]P.Garbeva,J.D.V.Elsas,J.A.V.Veen,Rhizosphere microbial community and its response to plant species and soil history,Plant Soil 302(2008)19-32.
[2]J.A.Lucas,A.Garcia-Villaraco,B.Ramos,J.Garcia-Cristobal,E.Algar,J.Gutierrez-Manero,Structural and functional study in the rhizosphere of Oryza sativa L.plants growing under biotic and abiotic stress,J.Appl.Microbiol.115(2013)218-235.
[3]E.R.Hunting,C.M.White,M.van Gemert,D.Mes,E.Stam,H.G. van,G.der,M.H.Kraak,W.Admiraal,UV radiation and organic matter composition shape bacterial functional diversity in sediments,Front.Microbiol.4(2013)317.
[4]M.Fang,P.P.Motavalli,R.J.Kremer,K.A.Nelson,Assessing changes in soil microbial communities and carbon mineralization in Bt and non-Bt corn residue-amended soils,Appl.Soil Ecol.37(2007)150-160.
[5]H.Fang,B.Dong,H.Yan,F.Tang,B.Wang,Y.Yu,Effect of vegetation of transgenic Bt rice lines and their straw amendment on soil enzymes,respiration,functional diversity and community structure of soil microorganisms under field conditions,J.Environ.Sci.24(2012)1259-1269.
[6]H.Hu,M.Xie,Y.Yu,Q.Zhang,Transgenic Bt cotton tissues have no apparent impact on soil microorganisms,Plant Soil Environ.59(2013)366-371.
[7]C.B.Blackwood,J.S.Buyer,Soil microbial communities associated with Bt and non-Bt corn in three soils,J.Environ. Qual.33(2004)832-836.
[8]Y.J.Chun,H.J.Kim,K.W.Park,S.C.Jeong,B.Lee,K.Back,H.M. Kim,C.G.Kim,Two-year field study shows little evidence that PPO-transgenic rice affects the structure of soil microbial communities,Biol.Fertil.Soils 48(2012)453-461.
[9]R.Miethling-Graff,S.Dockhorn,C.C.Tebbe,Release of the recombinant Cry3Bb1 protein of Bt maize MON88017 into field soil and detection of effects on the diversity of rhizosphere bacteria,Eur.J.Soil Biol.46(2010)41-48.
[10]R.J.Singh,I.Ahlawat,S.Singh,EffectsoftransgenicBtcottonon soil fertility and biology under field conditions in subtropical inceptisol,Environ.Monit.Assess.185(2013)485-495.
[11]Y.He,T.Ning,T.Xie,Q.Qiu,L.Zhang,Y.Sun,D.Jiang,K.Fu,F.Yin,W.Zhang,L.Shen,H.Wang,J.Li,Q.Lin,Y.Sun,H.Li,Y. Zhu,D.Yang,Large-scale production of functional human serum albumin from transgenic rice seeds,Proc.Natl.Acad. Sci.U.S.A.108(2011)19078-19083.
[12]W.Liu,H.Hao Lu,W.Wu,Q.Kun Wei,Y.Xu Chen,J.E.Thies,Transgenic Bt rice does not affect enzyme activities and microbial composition in the rhizosphere during crop development,Soil Biol.Biochem.40(2008)475-486.
[13]S.H.Lee,C.G.Kim,H.Kang,Temporal dynamics of bacterial and fungal communities in a genetically modified(GM)rice ecosystem,Microb.Ecol.61(2011)646-659.
Open Access full texts are available online in ScienceDirect (http://www.sciencedirect.com/science/journal/22145141)
The homepage ofTheCropJournal (http://www.journals.elsevier.com/the-crop-journal)
11 August 2014
in revised form
s.
E-mail addresses:xujingwang0514@126.com(X.Wang),wangcotton@126.com(Z.Wang).
Peer review under responsibility of Crop Science Society of China and Institute of Crop Science,CAAS.
http://dx.doi.org/10.1016/j.cj.2014.11.001
2214-5141/©2015 Production and hosting by Elsevier B.V.on behalf of Crop Science Society of China and Institute of Crop Science,CAAS. This is an open access article under the CC BY-NC-ND license(http://creativecommons.org/licenses/by-nc-nd/4.0/).
杂志排行

The Crop Journal的其它文章
- An AFLP marker linked to the leaf rust resistance gene LrBi16 and test of allelism with Lr14a on chromosome arm 7BL
- Genetic variation for phytic acid content in mungbean(Vigna radiata L.Wilczek)
- Quantifying cardinal temperatures and thermal time required for germination of Silybum marianum seed
- Characterization and mapping of QTLs on chromosome 2D for grain size and yield traits using a mutant line induced by EMS in wheat
- Reduced grain chalkiness and its possible physiological mechanism in transgenic rice overexpressingL-GalLDH
- Bed planting of wheat(Triticum aestivum L.)improves nitrogen use efficiency and grain yield compared to flat planting