寒地粳稻低温发芽力和芽期耐冷性与SSR标记的关联分析
2015-11-11
寒地粳稻低温发芽力和芽期耐冷性与SSR标记的关联分析
邹德堂,孙桂玉,王敬国,刘化龙,孙健
(东北农业大学农学院,哈尔滨150030)
摘要:以220份寒地粳稻品种资源进行低温发芽力和芽期耐冷性鉴定,利用118对SSR标记对低温发芽力和芽期耐冷性进行SSR标记关联分析。结果表明,品种间低温发芽力和芽期耐冷性存在明显差异。与相对发芽势、相对发芽率、相对发芽指数和成苗率相关联的SSR位点分别有4、2、3和4个。其中RM1220同时与低温发芽力的3个性状相关联,RM109和RM453同时与两个性状相关联,通过优异等位基因挖掘,筛选出一系列优异等位变异及相应载体材料,这些分子标记、优异等位变异和载体材料可为寒地粳稻耐冷育种提供有价值的信息和材料。
关键词:寒地粳稻;芽期;耐冷性;SSR标记;关联分析
网络出版时间2015-3-13 15:24:00
[URL]http://www.cnki.net/kcms/detail/23.1391.S.20150313.1524.015.html
邹德堂,孙桂玉,王敬国,等.寒地粳稻低温发芽力和芽期耐冷性与SSR标记的关联分析[J].东北农业大学学报, 2015, 46(3): 1-8.
水稻是喜温作物,低温可对水稻生长发育和产量造成影响[1]。黑龙江省低温冷害平均每3~4年发生1次,黑龙江省近40年出现严重影响全省水稻产量低温冷害9次[2]。2002年受害面积达100万hm2,受害区域水稻平均减产30%,减收18亿kg[3]。水稻从种子发芽到成熟整个生长发育期都可能遭受低温冷害。水稻芽期冷害一般发生在直播田和秧田,直接造成成苗率下降、烂秧、弱苗、抽穗分散和成熟期不一致,导致水稻产量降低[4]。
实践证明,利用耐冷基因,选育耐冷水稻品种是解决芽期低温冷害的有效方法[5-6]。水稻芽期耐冷性多受数量性状基因(QTL)控制,纪素兰等利用籼粳交的自交群体,15℃条件下测定种子平均发芽率并进行QTL分析,在同一年中共检测到11个控制种子低温发芽力的QTL[7];陈亮等利用籼粳交的DH群体,15℃条件下处理6 d,检测到2个与低温发芽力有关的QTL,分别位于第3和第10染色体上[8];姜旋等利用RIL群体检测到7个与水稻低温发芽力有关的QTL,分布在第1、3、5、6、8染色体上,其中贡献率最大为16.4%[9];周勇等利用水稻单片段代换系在4℃低温条件下处理10 d,以成苗率作为芽期耐冷性鉴定指标,共检测出18个与芽期耐冷性相关的QTL[10];陈玮等利用纸卷法测定水稻RIL群体在10℃下低温耐冷性,检测到4个QTL,分别位于第1、3、7和11染色体上[11];Lin等以Nipponbare为供体亲本,9311为受体亲本,1 500个BC4F1群体,检测到4个芽期耐冷性QTL[12]。
以上研究均为基于连锁分析的QTL检测,而连锁分析只能鉴别出两个亲本之间等位基因优劣,不能发现该物种中最优等位变异,而关联分析方法不但可检测到与性状有关的分子标记位点,还可挖掘自然群体中的优异等位基因[13]。在水稻耐冷性方面,张艳梅等用84个SSR标记对96份东北粳稻品种进行SSR标记关联分析,结果检测到与分蘖期11个性状相关联的18个位点,累计25次显著关联[14]。而有关水稻芽期耐冷性关联分析的研究则未见报道。本研究利用寒地粳稻220份品种资源,对低温发芽力和芽期耐冷性进行鉴定,并对SSR标记和低温发芽力及芽期耐冷性进行关联分析,旨在检测与芽期耐冷性有关的SSR标记位点,挖掘优异等位变异及载体材料,为寒地粳稻芽期耐冷性分子育种提供参考。
1 材料与方法
1.1材料
供试材料为220份寒地粳稻品种,由东北农业大学农学院水稻研究所提供。
1.2方法
1.2.1低温条件下种子发芽力鉴定
低温发芽力鉴定参照李太贵方法[15]。利用盐水法挑选出饱满的水稻种子,用次氯酸钠消毒20 min后清洗,置于恒温箱内,在45℃下处理48 h以打破休眠。每个品种取50粒种子均匀播在垫有两张滤纸直径为9 cm的培养皿内,加入10 mL蒸馏水,置于10℃恒温培养箱中,处理14 d,逐日记录出芽情况。另设25℃处理为对照,3次重复。以相对发芽势(Relative germination, RG),相对发芽率(Relative germination rate, RGR)和相对发芽指数(Relative germination index, RGI)评价品种低温发芽力。
处理试验以第11天的发芽率为发芽势,对照试验以第4天的发芽率为发芽势。
相对发芽势(%)=处理发芽势/对照发芽势× 100%;
发芽率(%)=发芽的种子/总粒数×100%;
相对发芽率(%)=处理发芽率/对照发芽率× 100%;
发芽指数GI=∑(Gt/Dt),Gt为在t日的发芽数,Dt为相应的发芽日数;GI越大,表明发芽速度越快。相对发芽指数(%)=处理发芽指数/对照发芽指数×100%。
1.2.2芽期耐冷性鉴定
芽期耐冷性鉴定参照韩龙植等方法[16]。利用盐水法挑选出饱满的水稻种子,用次氯酸钠消毒20 min后清洗,置于恒温箱内,在45℃下处理48 h以打破休眠。每个品种取出50粒种子置于垫有两层滤纸直径为9 cm培养皿内,加适量水,使种子一半体积淹没在水里。在32℃恒温培养箱中催芽。当芽长长至5 mm时,置于培养箱中5℃低温处理10 d,随后将培养皿置于20~30℃有阳光的室内环境下正常生长10 d,调查活苗数,并计算成苗率。以成苗率作为水稻芽期耐冷性鉴定标准(见表1),3次重复,以平均值作为统计单位。
成苗率(%)=成活苗数/出芽总粒数×100%
1.3 DNA提取及SSR分析
根据Doyle等改进的CTAB法提取水稻基因组DNA[17]。随机抽取10份材料用于SSR引物多态性筛选,从1 000对SSR引物中筛选出118对均匀分布于水稻12条染色体且多态性高的引物,引物序列信息从www.gramene.org网站获得。SSR-PCR反应体系及扩增程序参照Chen等方法进行[18],PCR产物经6%聚丙烯酰胺凝胶电泳进行分离,银染法检测,并拍照记录。

表1 水稻芽期耐冷性分级标准Table 1 Grading standard of cold tolerance at bud stage in rice
1.4统计分析
1.4.1表型数据统计分析
采用SPSS 17.0数据分析软件进行相关性分析,利用Excel 2010进行描述性统计。
1.4.2连锁不平衡与群体结构分析
利用Tassel 2.1分析群体连锁不平衡程度[19],并参照Temnykh等[20]和McCouch等[21]遗传图谱估计遗传距离,使用Structure 2.2软件估测样本群体结构[22],利用118个位点中>30 cM的50对SSR位点进行群体结构分析,将MCMC(Markov chain monte carlo)开始时的不作数迭代(Length of burn-in period)设为10 000次,再将不作数迭代后的MCMC设为100 000次,K值设为1~10,重复运算10次,以极大似然值最大为原则,选择合适的K值。
1.4.3关联分析
使用Tassel 2.1软件的一般线性模型(General linear model,GLM)和混合线性模型(Multiple lin⁃ear model,MLM)分别进行性状与标记的关联分析。其中GLM以每个个体Q值作为协变量进行回归分析,MLM以Q+K方法分析,选择计算每个标记的遗传力的计算方式,并选择EM方法进行分析,在显著水平(P<0.01)下,检测与表型性状相关的标记位点。
1.4.4优异等位基因的挖掘
根据Breseghello等提出的无效等位变异方法[23]:

式中,ai表示第i个等位变异的表型效应值,其中xij为携带第i个等位变异的第j个材料性状表型测定值,ni为具有第i个等位变异的材料数。Nk为携带无效等位变异的第k个材料的表型测定值,nk为具有无效等位变异的材料数。若ai>0,则认为该等位变异为增效等位变异,反之,ai<0为减效等位变异。
2 结果与分析
2.1寒地粳稻低温发芽力与芽期耐冷性鉴定
2.1.1低温发芽力鉴定
对220份寒地粳稻进行低温条件下种子发芽力鉴定(见图1)。

图1 低温条件下相对发芽势、相对发芽率和相对发芽指数在供试群体中的分布Fig. 1 Distribution of relative germination,relative germination rate and relative germination index inthe test population at low temperature
图1结果表明,相对发芽势在0~20%内有86份,占39.10%;20%~40%有67份,占30.45%;40%以上有67份,占30.45%,其中相对发芽势较高的代表材料有龙粳21号、牡丹江4号、松粳3号、松粳8号和松粳12号。相对发芽率分布较均匀,在0~20%内有22份,占10%;20%~40%有67份,占30.45%;40%~60%有56份,占25.45%;60%~80%有42份,占19.10%;80%~100%有33份,占15%,其中相对发芽率较高的代表材料有龙粳21号、牡丹江4号、龙盾103、松粳3号和松粳12号。相对发芽指数在0~10%内有86份,占39.10%;10%~20%有70份,占31.82%;20%~30%有37份,占16.82%,其中相对发芽指数较高的代表材料有龙粳21号、牡丹江4号、龙盾103、松粳3号和松粳12号。
2.1.2芽期耐冷性鉴定
对供试群体进行芽期耐冷性鉴定,结果表明(见图2),220份材料在低温下成苗率变化为0~ 100%,平均值为76.51%。按成苗率分级标准,1级极强耐冷品种有26份,占总材料的11.82%;3级强耐冷品种最多,有127份,占总材料的57.73%;5级中等耐冷品种有28份,占总材料的12.73%;7级较弱耐冷品种有25份,占总材料的11.36%;9级极弱耐冷品种有14份,占总材料的6.40%,芽期鉴定出耐冷性强的代表材料有龙粳21号、龙粳香1号、合江3号、龙盾103和松粳10号。耐冷性分级结果表明,寒地粳稻品种具有较丰富的芽期耐冷性表型多样性,各级耐性材料均有分布。

图2 水稻芽期耐冷性分级Fig. 2 Classification of rice cold tolerance at bud stage
通过对水稻低温发芽力和芽期耐冷性4个性状进行统计分析(见表1)可以发现,相对发芽势变异范围为2.04%~97.96%、平均值为31%,变异系数为76.31%;相对发芽率变异范围为4.55%~100%,平均值为48.82%,变异系数为48.82%;相对发芽指数变异范围为1.23%~62.46%,平均值为15.08,变异系数为73.35%;成苗率变异范围为0~100%,平均值为76.51%,变异系数为40.12%。4个性状在220份材料中变化规律相近,大部分品种每个性状数值高低趋势一致,说明4个性状耐冷性鉴定结果在各材料之间有较好的吻合性,可反映水稻芽期耐冷性。

表1 寒地粳稻低温发芽力和芽期耐冷性表现Table 1 Performance of low temperature germinability and cold tolerance at bud stage of japonica rice in cold region
2.2低温发芽力与芽期耐冷性的相关性分析
对供试材料相对发芽势、相对发芽率和相对发芽指数等4个性状进行相关分析,由表2可知,3个性状间均呈极显著正相关,其中相对发芽势与相对发芽指数相关系数为0.975,与相对发芽率相关系数为0.886;相对发芽指数与相对发芽率相关系数为0.900;成苗率与相对发芽势和相对发芽率的相关系数分别是0.559和0.448。
2.3连锁不平衡及群体结构分析
关联分析以不同位点等位基因之间的连锁不平衡为基础,图3结果显示118个SSR标记在各连锁群上的连锁不平衡状况。在检测到的6 903种SSR位点成对组合中,无论共线性组合(同一连锁群)或者非共线组合(不同连锁群)都存在一定程度LD存在,r2统计概率(P<0.01)支持的LD成对位点4 505个,占全部位点组合的65.26%,r2平均值为0.011。
利用STRUCTURE 2.2软件分析供试群体的群体结构(见图4),结果表明,LnP(D)的值持续增长,没有拐点,因此用ΔK值确定K值,且K=2时极大似然值最大,并有较稳定的值,因此认为该群体有两个亚群。
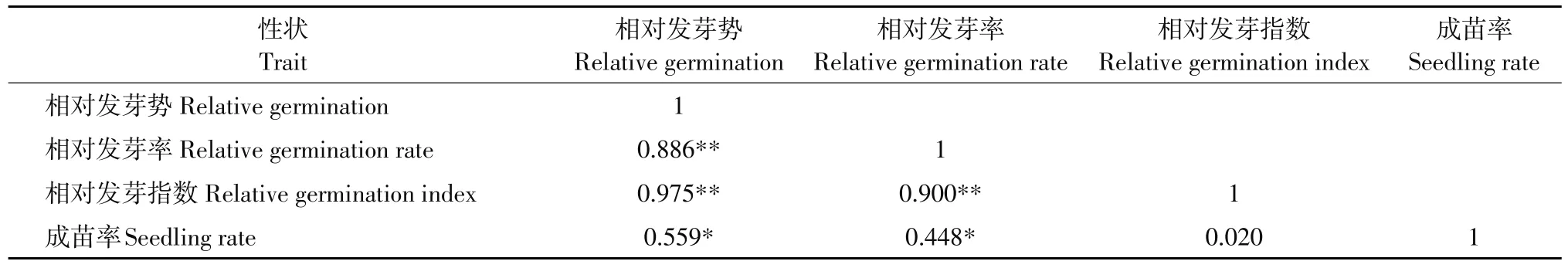
表2 低温发芽力之间的相关性分析Table 2 Correlation analysis of low temperature germinability

图4 STRUCTURE软件分析预测的LnP(D)、ΔK折线Fig. 4 Estimated LnP(D) and ΔK by STRUCTURE analysis
2.4低温发芽力和芽期耐冷相关性状的关联分析
通过Tassel 2.1软件的GLM和MLM两种方法,对低温发芽力和芽期耐冷相关性状进行关联分析(见表3)。运用GLM(P<0.01)方法,检测到有24个SSR位点与相对发芽势、相对发芽率、相对发芽指数及成苗率等4个性状相关,运用MLM(P<0.01)方法检测到11个SSR位点与4个性状相关。两种方法共同检测到9个SSR位点,累计13次显著相关,分布在1、2、3、5、6和12条染色体上。从表3中可以看出,与相对发芽势关联的位点有4个,与相对发芽率关联的位点有2个,与相对发芽指数关联的位点有3个,与成苗率关联的位点有4个。其中RM1220同时与低温发芽力的3个性状相关联,RM109和RM453同时与两个性状相关联。

表3 与低温发芽力和成苗率的相关SSR标记位点表型变异的解释率Table 3 Explained phenotypic variation at loci highly associated with low temperature germinability and percentage of seedling emergency
2.5优异等位基因的挖掘
由表4可知,与相对发芽势关联的4个标记中,共有15个等位变异,包括6个增效等位变异和9个减效等位变异,增效等位变异对品种具有增效作用,减效等位变异反之。其中增效等位变异平均效应最大的是RM1251,代表材料有垦粳3号等;与相对发芽率关联的2个标记中共有8个等位变异,包括5个增效等位变异和3个减效等位变异。其中RM1220增效等位变异最多,有4个,RM1220- 167 bp表型效应最大,为32.68,代表材料有垦稻22号等;与相对发芽指数关联的3个标记中有12个等位变异,包括9个增效等位变异和3个减效等位变异。其中RM1220-167 bp表型效应最大,为9.78,代表材料有老头稻等;与成苗率关联的4个标记中有13个等位变异,包括6个增效等位变异和7个减效等位变异。其中RM480-216 bp表型效应最大,为15.79,代表材料有龙粳20号等。
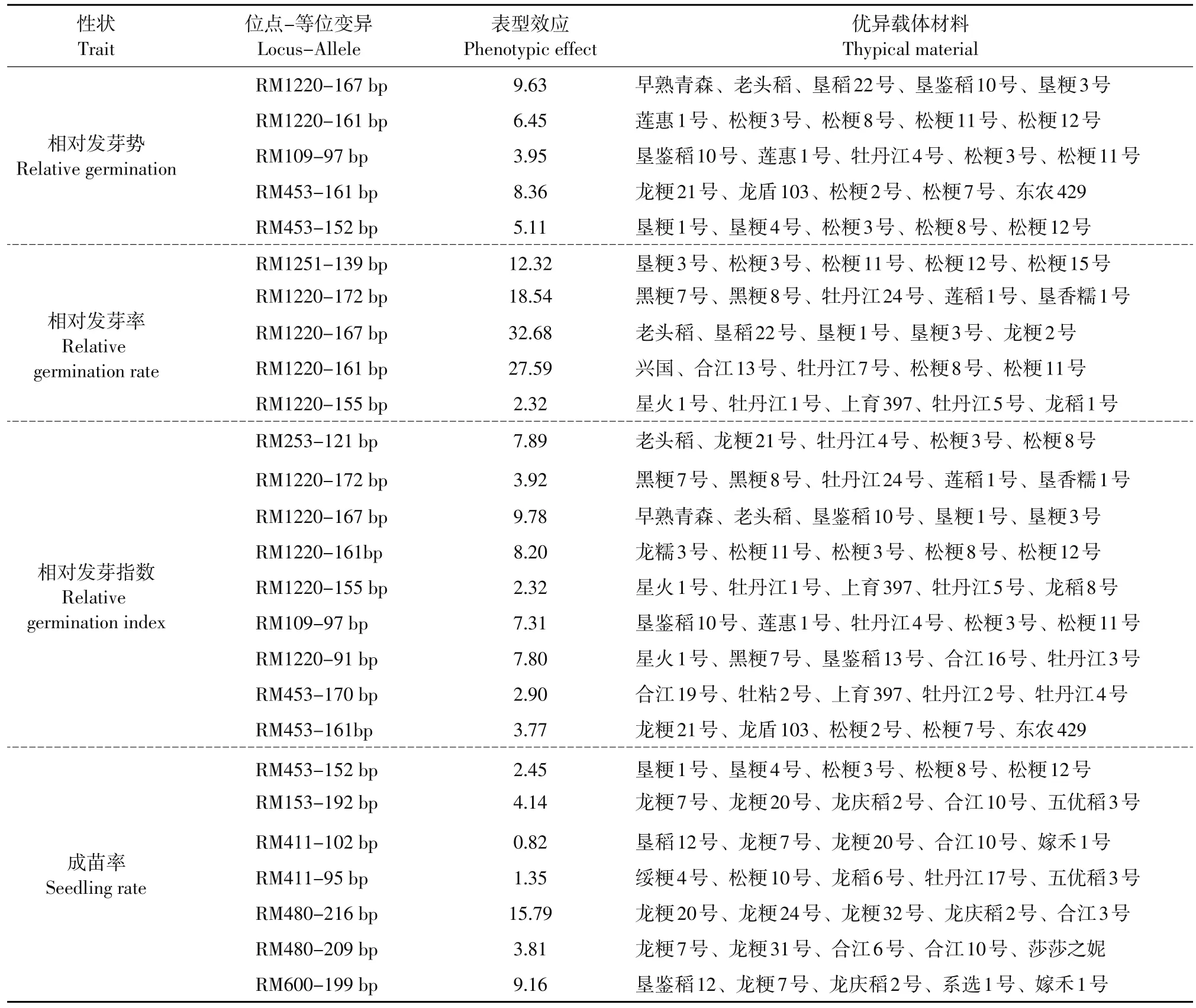
表4 与芽期耐冷相关的优异等位基因及载体材料Table 4 Favorable alleles and their carries related to cold tolerance at bud stage
3 讨论与结论
水稻低温发芽力和芽期耐冷性是反应水稻耐冷性的重要指标。金铭路等研究表明低温发芽力和芽期耐冷性较强的水稻种质一般表现为幼苗耐冷性和孕穗期耐冷性较强,并且低温下解释率的变异比较小,低温发芽力强的水稻种质一般表现为较强的芽期耐冷性[24]。李太贵利用50个水稻品种进行多个时期耐冷性测定,结果表明,芽期耐冷性与苗期和开花期耐冷性呈显著正相关[25]。韩龙植等研究表明,芽期耐冷性与苗期生长、抽穗天数、穗抽出度、孕穗期耐冷性和成熟期耐冷性有密切相关的关系,通过对芽期耐冷性的筛选一般可得到低温下幼苗生长能力强、生育期较早、穗抽出度良好、孕穗期和成熟期耐冷性综合较强的品种[26]。蒋向辉等研究表明,水稻低温发芽力与幼苗耐冷性呈显著正相关,同时各品种间常温发芽率与14℃发芽率呈极显著正相关[27]。通过对水稻低温发芽力和芽期耐冷性的鉴定不仅能反应芽期耐冷性,还可以为苗期及生育后期耐冷性鉴定提供参考。本研究以黑龙江寒地粳稻品种资源为材料,通过芽期耐冷性鉴定,筛选出强耐冷性品种,包括龙粳21号、合江3号、龙盾103、牡丹江4号、松粳3号和松粳12号等,可以为寒地粳稻苗期及生育后期耐冷性筛选提供参考。
本研究利用Tassel 2.1软件中的GLM检测到24个与低温发芽力和芽期耐冷性关联的SSR标记位点,利用MLM检测到11个SSR标记位点,综合两种方法共同检测到9个相同的SSR位点。本研究中检测到与相对发芽势关联的位点有4个,与相对发芽率关联的位点有2个,与相对发芽指数关联的位点有3个,与成苗率关联的位点有4个。邓久英检测到1个苗期耐冷性QTL,位于RM580~RG811[28],纪素兰等检测到一个与低温发芽力相关的QTL,位于C560~C1470[7],陈玮检等测到1个芽期耐冷性QTL,位于RM156~RM16[11]。本研究检测到的RM600、RM1251和RM411均在其内;张艳梅等检测到与成熟期耐冷性关联的RM480标记位点在本研究中也检测到[1];Andaya等研究中检测到与水稻
孕穗期耐冷性相关的RM253在本研究中也检测
到[29],另外,RM453和RM153也与前人检测到的水稻耐冷性位点相近[14]。以上与本研究检测到的相同或相近区间QTL位点可能是与耐冷性有关的重要位点,也进一步验证关联分析和连锁分析互补性。但本研究检测到的RM1220和RM109位点与前人研究不一致,可能是与水稻低温发芽力相关的新位点。
本研究筛选的优异等位基因载体材料具有多个等位变异,松粳3号有8个等位变异,松粳6号、松粳11号和松粳12号具有6个等位变异,老头稻、垦鉴稻10号、垦粳1号、龙粳7号和牡丹江4号等具有4个等位变异,这些优异等位变异及载体材料可为寒地粳稻耐冷育种提供更多的基因位点和亲本材料。
[参考文献]
[1]张艳梅,王敬国,刘化龙,等.东北粳稻不同生育期耐冷性分析[J].东北农业大学学报, 2012, 43(4): 43-44.
[2]王晓群,张宇.黑龙江省水稻低温冷害风险评估[J].安徽农业科学, 2012, 40(1): 263-266.
[3]徐希德.低温冷害对黑龙江省水稻的影响极其防御对策[J].中国农学通报, 2003, 19(5): 135-136.
[4]乔永利,韩龙植,安永平,等.水稻芽期耐冷性QTL的分子定位[J].中国农业科学, 2005, 38(2): 217-221.
[5]崔迪,杨春刚,汤翠凤,等.低温胁迫下粳稻选育品种耐冷性状的鉴定评价[J].植物遗传资源学报, 2012, 13(5): 739-747.
[6]邹德堂,李娇,郑洪亮,等.冷水胁迫下水稻分蘖期耐冷性状QTL定位分析[J].东北农业大学学报, 2012, 43(10): 96-102.
[7]纪素兰,江令,王益华,等.水稻种子耐低温发芽力的QTL定位及上位性分析[J].作物学报, 2008, 34(4): 551-556.
[8]陈亮,娄巧军,孙宗修,等.水稻低温发芽力的QTL定位[J].中国水稻科学, 2006, 20(2): 159-164.
[9]姜旋,李辰昱,毛婷,等.水稻低温发芽性QTL的分子标记定位[J].武汉植物学研究, 2005, 23(3): 216-220.
[10]周勇,朱孝波,袁华,等.水稻单片段代换系芽期和苗期耐冷性分析及耐冷性QTL鉴定[J].中国水稻科学, 2013, 27(4): 381-388.
[11]陈玮,李炜.水稻RIL群体芽期耐冷性基因的分子标记定位[J].武汉植物学研究, 2005, 23(2): 116-120.
[12]Lin J, Zhu W Y, Zhang Y D, et al. Detection of QTL for cold toler⁃ance at bud bursting stage using chromosome. Segment substitu⁃tion lines in rice (Oryza sativa L.)[J]. Rice Sci, 2011, 18: 71-74.
[13]陈氏秋江,党小景,刘强明,等.水稻籽粒性状的SSR关联分析[J].中国水稻科学, 2014, 28(3): 243-257.
[14]张艳梅,邹德堂.东北粳稻分蘖期耐冷性鉴定及SSR标记关联分析[J].中国水稻科学, 2012, 26(4): 423-430.
[15]李太贵.在低温下筛选水稻不同生长期耐寒品种的室内方法[J].国外农业科技, 1984(4): 18-21.
[16]韩龙植,曹桂兰,安永平,等.水稻种植资源芽期耐冷性的鉴定与评价[J].植物遗传资源学报, 2004, 5(4): 346-350.
[17]Doyle J J , Doyle J I. Isolation of plant DNA from fresh tissue[J]. Focus, 1990(12): 149-151.
[18]Chen X, Temnykh S, Xu Y, et al. Development of a microsatellite framework map providing genome-wide coverage in rice (Oryza sativa L.) [J]. Theoretical and Applied Genetics, 1997, 95: 553-567.
[19]Bradbury P J, Zhang Z, Kroon D E, et al. TASSEL: Software for as⁃sociation mapping of complex traits in diverse samples[J]. Bioin⁃formatics, 2007, 23: 2633-2635.
[20]Temnykh S, DeClerk G, Lukashova A, et al. Computational and experimental analysis of microsatellites in rice(Oryza sativa L.): Frequency, length variation, transposon associations, and genetic marker potential[J]. Genom Res, 2001, 11(8): 1441-1452.
[21]McCouch S R, Teytelman L, Xu Y, et al. Development and map⁃ping of rice (Oryza sativa L.) [J]. DNA Res, 2002(9): 199-207.
[22]Pritchard J K, Stephens M, Rosenberg N A, et al. Association mapping in structured populations[J]. Am J Hum Genet, 2000, 67: 170-181.
[23]Breseghello F, Mark E S. Association mapping of kernel size and milling quality in wheat (Triticum aestivum L.) cultivars[J]. Genet⁃ ics, 2006, 172: 1165-1177.
由本试验结果可知,根据氮肥施用量不同采用氮肥施用方法不同,氮肥施用量较低时增加生育前期氮肥施用量有利于提高单株穗数、每穗粒数和单株粒重,而氮肥施用量较高时应增加生育后期氮肥施用量,保证千粒重和单株粒重的提高。增加氮肥施用量时,相应增加钾肥施用量和减少生育前期氮肥施用量将有利于提高茎秆抗倒特性。因此,高产超高产水稻栽培不仅重视氮肥施用量的增加和施用方法,更要重视钾肥施用量的增加和配比方法,以改善茎秆理化特性和提高抗倒伏性,保证水稻高产高效。
[1]杨慧杰,杨仁崔,李义诊,等.水稻茎秆性状与抗倒性的关系[J].福建农业大学学报, 2000, 15(2): 1-7.
[2]李扬汉.禾本科作物的形态与解剖[M].上海:上海科学技术出版社, 1979: 143-148.
[3]杨世民,谢力,郑顺林,等.氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响[J].作物学报, 2009, 35(1): 93-103.
[4]李娟,黄平娜.不同施肥模式对水稻生理特性、产量及其N肥农学利用率的影响[J].核农学报, 2011, 25(1): 169-173.
[5]李国辉,钟旭华,田卡,等.施氮对水稻茎秆抗倒伏能力的影响及其形态和力学机理[J].中国农业科学, 2013, 46(7): 1323-1334.
[6]张明聪,刘元英,罗盛国,等.养分综合管理对寒地水稻抗倒伏性能的影响[J].中国农业科学, 2010, 43(21): 4536-4542.
[7]郭玉华,朱四光,张龙步.不同栽培条件对水稻茎秆生化成分的影响[J].沈阳农业大学学报, 2003, 34(2): 89-91.
[8]鲍士旦.土壤农化分析[M].北京:中国农业出版社, 2000: 30-107.
[9]中国土壤学会农业化学专业委员会.土壤农业化学常规分析方法[M].北京:科学出版社, 1983: 2742-286.
[10]杨守仁.水稻理想株型育种新动态[J].中国水稻科学, 1988, 2 (3): 1-5.
[11]袁隆平.杂交水稻超高产育种[J].杂交水稻, 1997, 12(6): 1-3.
[12]陈温福,徐正进,张龙步.水稻理想株型的研究[J].沈阳农业大学学报, 1989, 20(4): 417-420.
[13]马均,马文波,田彦华,等.重穗型水稻植株抗倒伏能力的研究[J].作物学报, 2004, 30(2): 143-148.
[14]梁康迳,王雪仁,章清杞,等.基因型×环境互作效应对水稻茎秆抗倒性杂种优势的影响[J].福建农业大学学报, 2000, 15(1): 12-17.
[15]杨长明,杨林张,颜廷梅,等.不同养分和水分管理模式对水稻抗倒伏能力的影响[J].应用生态学报, 2004, 15(4): 646-650.
[16]张丰转,金正勋,马国辉,等.水稻抗倒性与茎秆形态性状和化学成分含量间相关分析[J].作物杂志, 2010(4): 15-19.
[17]张喜娟,李红娇,李伟娟,等.北方直立穗型粳稻抗倒性的研究[J].中国农业科学, 2009, 42(7): 2305-2313.
[18]罗茂春,田翠婷.水稻茎秆形态结构特征和化学成分与抗倒伏关系综述[J].西北植物学报, 2007, 27(11): 2346-2353.
Zou detang, Sun guiyu, Wang jingguo, et al. Association analysis of germinability under low temperature and cold tolerance at bud stage with SSR markers of japonica rice in cold region[J]. Journal of Northeast Agricultural University, 2015, 46(3): 1-8. (in Chinese with English abstract)
Association analysis of germinability under low temperature and cold tolerance at bud stage with SSR markers of japonica rice in cold
region
/ZOU Detang, SUN Guiyu, WANG Jingguo, LIU Hualong, SUN Jian (School of Agriculture,Northeast Agricultural University, Harbin 150030, China)
Abstract:In order to investigate the germinability under low temperature and the cold tolerance at the bud stage of japonica rice, 220 varieties of japonica rice were used as materials and 118 pairs of SSR primes were selected. The results showed that there was a significant difference between varieties of the germinability under low temperature and the cold tolerance at the bud stage. The relative germination, relative germination rate, relative germination index and seedling rate associated SSR loci were 4, 2, 3 and 4, respectively. RM1220 was associated with three traits of the germinability under low temperature , RM109 and RM453 were associated with two traits, Through mining the superior allele, we can screen a series of favorable alleles and the corresponding carrier material, which would provide valuable information and materials for breeding cold tolerance of japonica rice in cold region.
Key words:japonica rice in cold region; bud stage; cold tolerance; SSR marker; association analysis
作者简介:邹德堂(1965-),男,教授,博士,博士生导师,研究方向为水稻遗传育种。E-mail: zoudt@163. com
基金项目:“十二五”农村领域国家科技计划课题(2013BAD20B04);国家科技支撑计划(2011BAD16B11,2011BAD35B02-01-01)
收稿日期:2014-12-07
文章编号:1005-9369(2015)03-0001-08
文献标志码:A
中图分类号:S511
