珙桐未成熟胚的萌发及植株再生的研究*
2015-08-16方志荣蔡光泽
方志荣,蔡光泽
(西昌学院,四川 西昌615013)
珙桐未成熟胚的萌发及植株再生的研究*
方志荣,蔡光泽
(西昌学院,四川 西昌615013)
为了建立珙桐的再生体系,试验以珙桐未成熟胚为外植体,获得了再生植株。采用正交设计和极差分析法对影响未成熟胚再生植株的四种主要因素α-萘乙酸(α-Naphthaleneacetic acid,NAA)、噻苯隆(Thidiazuron,TDZ)、赤霉素(gibberellic acid,GA)和活性炭(active carbon,AC)进行了研究。结果表明:胚萌发的最佳培养基组合为WPM+0.09 mg/L NAA +0.6 mg/L TDZ+0.9 mg/L GA+0.8 g/L AC,芽诱导率为71.43%;萌发胚芽分化及生根的最佳培养基组合为WPM培养基+ 0.1 mg/L NAA+3 mg/L TDZ+6 mg/L GA+1.2 g/L AC,再生率为50%。AC能促进下胚轴变绿和子叶变绿,NAA、GA对解除胚的形态后熟有重要作用。GA和TDZ在芽分化及生根过程中起主导作用。
珙桐;未成熟胚;再生植株
珙桐(Davadia involucrataBaill)又名鸽子树,为我国特有的珙桐科(Davadiaceae)单型属植物。系第三纪古热带植物区系的孑遗种,有“活化石”之称,是世界著名的观赏植物,被列为国家一级珍稀濒危保护植物[1]。珙桐天然分布在我国的贵州、湖南、湖北、陕西、四川、云南以及甘肃7省,7省内分布范围较窄,资源量较少,仅限于一些边远山区人迹罕到之处[2]。
在自然状态下,珙桐种子休眠期长,萌发率低,存在二次休眠,其中种胚的形态后熟和生理后熟是种子休眠的主要原因[3,4]。新采收的珙桐种子必须在湿砂中层积近2年才能逐渐分化成具有纤毛和叶脉的胚芽,完成形态后熟。此后大部分种子进入二次休眠,须继续层积2年,才能迅速萌发[3]。利用成熟期果实中的未成熟胚为外植体,采用加入外源激素的办法可以缩短未成熟胚完成形态后熟的时间,实现未成熟胚萌发[5,6]。然而,迄今为止尚无利用珙桐成熟前期果实中的未成熟胚直接再生植株并无报道。
本研究以珙桐成熟前期果实中的未成熟胚为材料,研究了以珙桐未成熟胚为外植体直接再生植株的培养基配方,以期为研究珙桐未成熟胚的后熟提供材料,为珙桐未成熟胚再生体系激素配方的研究提供理论基础。
1 材料与方法
1.1 试验材料
2012年9月1日于四川省雅安市荥经县龙苍沟(海拔:1530 m,N 29.33°,E 102.44°)采集珙桐当年生果实,带回实验室后4℃保存。
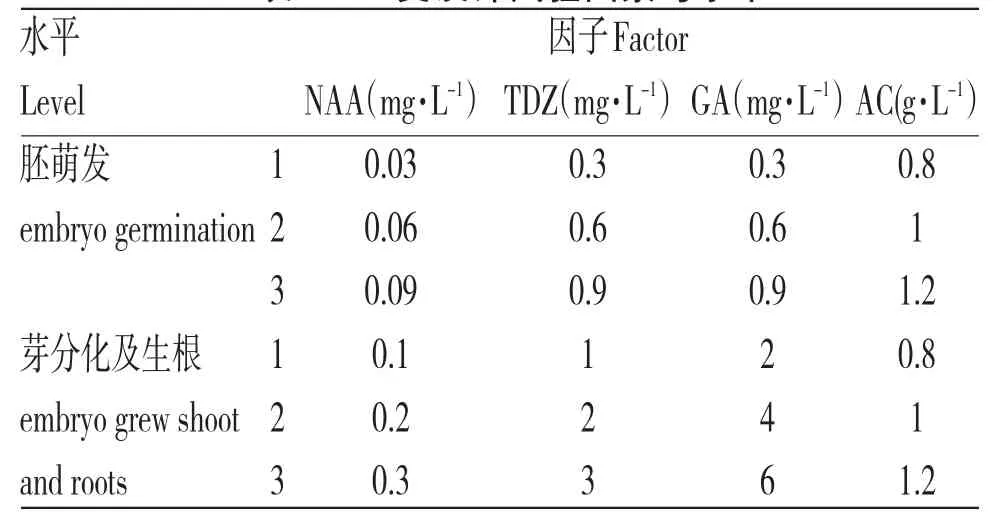
表1 正交设计试验因素与水平
1.2 研究方法
1.2.1 培养基的筛选
本实验共分为两个阶段,第一阶段先诱导未成熟胚萌发;第二阶段诱导芽分化及生根。以WPM培养基为基础培养基,采用正交设计法对影响珙桐胚培养的四种主要因素α-萘乙酸( α-Naphthaleneaceticacid,NAA)、噻 苯 隆(Thidiazuron,TDZ)、赤霉素(gibberellic acid,GA)和活性炭(active carbon,AC)进行研究,试验因素及水平设置见表1。试验方案及结果分析见表2,表3。
1.2.2 接种及培养试验
剥去珙桐果实的外果皮和中果皮后,用小铁锤小心敲开内果皮,从果核深沟里再小心挑出种子,剥去胚乳,将未成熟胚置于纯水中浸泡30分钟。在超净工作台上,先用75%乙醇灭菌20秒,无菌水冲洗3~4次,再用0.1%的升汞灭菌40秒,无菌水冲洗5次以上。将上述灭菌后的未成熟胚接种于含不同浓度激素的胚萌发培养基上。每瓶接2个胚,每组8瓶。将培养物放入人工气候箱中光照16 h,黑暗8 h,温度25℃培养。以上胚轴开始出现肉眼可见的芽为萌发的标准,30天后统计胚的萌发率。萌发率=(未污染萌发胚数/未污染总胚数)x100%。将未污染的萌发的胚转接到芽分化及生根培养基上。每瓶接1个胚,每组12瓶。
2 结果与分析
2.1 未成熟胚芽诱导、芽分化及生根

图1 珙桐未成熟胚的萌发及植株再生
有关珙桐休眠的原因主要有3种:(1)内果皮(图1①)坚硬,透水性和透气性差,及机械阻力造成珙桐种子休眠[4]。(2)未成熟胚的形态后熟和生理后熟是造成种子休眠的另一重要原因[3]。余阿梅等采用在湿沙中沉积1年的未成熟胚为材料,以1/2 MS+ 0.5 mg/L IBA+1 mg/L BA+2.85 mg/L GA+0.25 mg/L AC为培养基诱导未成熟胚萌发,实验中以幼根超过该胚的长度为萌发的标准,30天优化后的培养基萌发率为87.72%[5]。董社琴等以新采收珙桐果实的种胚为材料,未成熟胚4℃处理21天后进行培养,在培养后的56天发现MS+4.0 mg/L BA+12 mg/L GA+3 mg/L IAA培养基上的未成熟胚萌发[6]。本研究中,接种在芽诱导培养基上生长良好的白色胚(图1②)10天以后下胚轴及子叶变成绿色(图1③),30天后在上胚轴开始出现肉眼可见的芽(3 mm以上)(图1④),未成熟胚萌发的时间与前人报道相比有所缩短,此时的胚已经完成形态后熟。(2)珙桐果实中含有大量的抑制种子萌发的物质是造成珙桐种子休眠的又一重要的原因。除果肉,内果皮和胚乳外,胚自身也含有抑制萌发的物质[7]。对珙桐生殖物候的研究表明,珙桐5月下旬形成幼果,10月中下旬果实成熟,并开始脱落[8]。本研究取材的时间处于果实的生长期,此时未成熟胚中含有的萌发抑制物质低于成熟期的萌发抑制物质,可能是未成熟胚发芽时间缩短的原因。完成形态后熟的胚在芽分化及生根培养基上继续培养,则50天可以分化出芽(图1⑤)及70天后产生再生植株(图1⑥)。
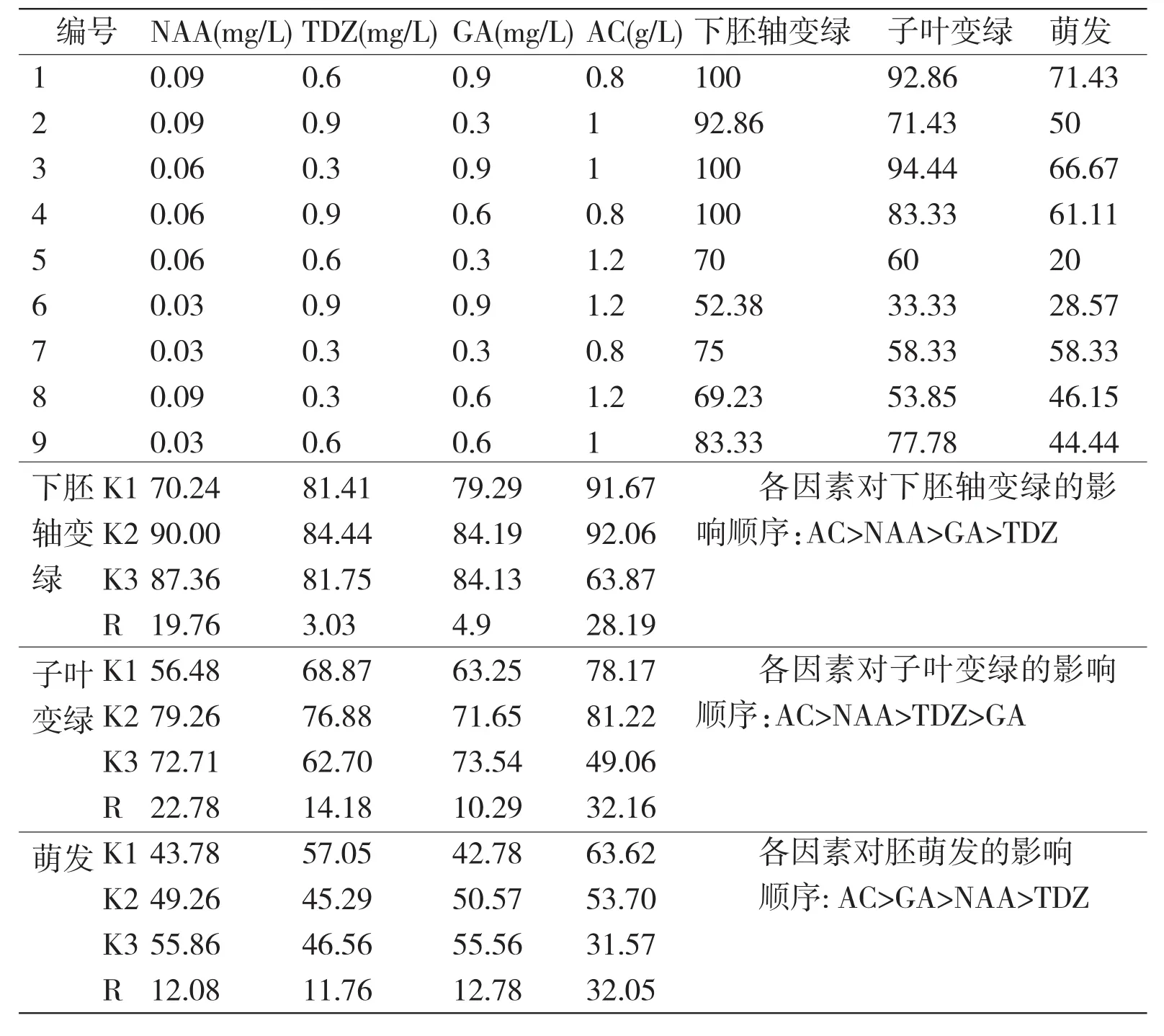
表2 未成熟胚芽诱导培养基正交设计方案、结果及极差分析
2.2 不同因素组合对未成熟胚芽诱导的影响
未成熟胚芽诱导可以分为下胚轴变绿、子叶变绿和萌发三个阶段(表1)。极差分析的结果表明,在三个阶段中,活性炭(AC)发挥了重要作用。这一结果与余阿梅试验结果一致,一定量的活性炭能吸附胚中的萌发抑制物质[5],然而过多的活性炭可能吸附培养基中的生长调节物NAA、TDZ、GA使未成熟胚芽诱导率下降。董社琴等报道了IAA对解除种子形态后熟有作用,GA对解除生理后熟至关重要[6]。从表1的分析可以得出:NAA在芽诱导的前期发挥了重要作用,到后期GA显示出重要的作用。综合表1的结果,可以得到优化后的培养基为WPM+0.09 mg/L NAA+0.3 mg/L TDZ+0.9 mg/L GA+0.8 g/L AC。
2.3 不同因素组合对萌发胚芽分化及生根的影响
陈坤荣等用GA和6-BA浸泡二次休眠的珙桐种子1周,可使珙桐种子在半个月内萌发出苗,说明珙桐种子完成形态后熟后,外源激素GA和6-BA对解除种子休眠至关重要[3]。雷泞菲等测定了珙桐种子休眠萌发过程中,内源激素的变化,发现在珙桐种子萌发的过程中,GA和KT的含量增加,NAA的含量也增加[9]。1976年德国先灵公司宣布TDZ (thidiazuron)合成,其化学名称为N-苯基-N’-(1,2, 3-噻二唑-5基)-脲。国内定名为噻苯隆。噻苯隆是一种新型高效的细胞分裂素用于组培能更好的促进植物的芽分化,其诱导植物细胞分裂的作用是6-BA的100倍[10]。从表2的极差分析结果可以得出,在芽分化及生根的过程中GA和TDZ起到了重要的作用,与陈坤荣、雷泞菲等研究的结果一致[3,9]。然而,本研究所得到的发生芽分化及生根的成熟胚的比例偏小,这可能与加入的外源激素的量有关。增加外源激素的加入量是否能促进芽分化及生根有待进一步的研究证实。
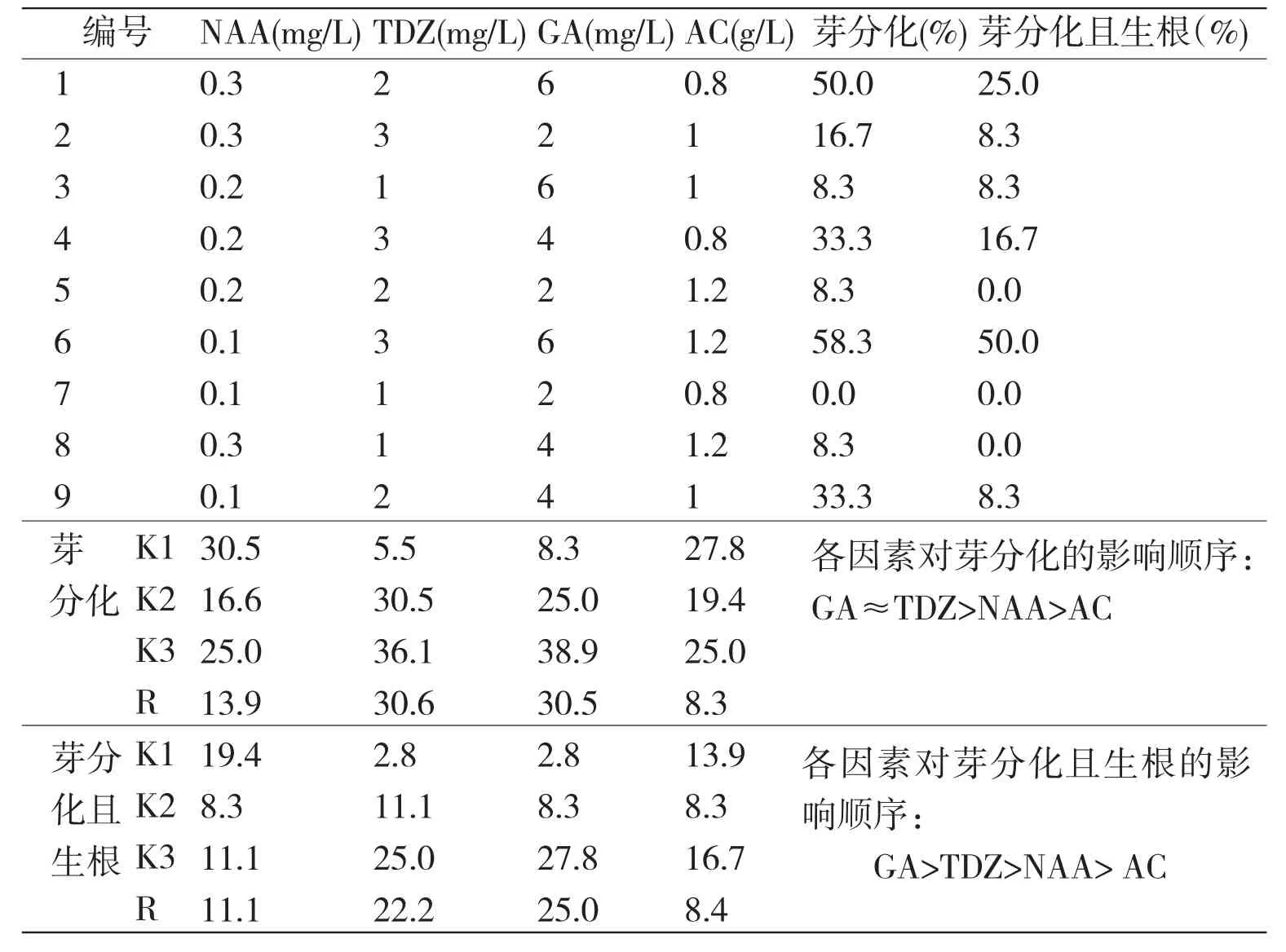
表3 芽分化及生根培养基正交设计方案、结果及极差分析
3 结论与讨论
本研究通过两步法即芽诱导、芽分化及生根,实现了珙桐种子直接再生植株。α-萘乙酸(NAA)、噻苯隆(TDZ)、赤霉素(GA)和活性炭(AC)在再生植株形成的不同阶段发挥的作用有所差异。在下胚轴变绿、子叶变绿阶段,AC和NAA起了主导作用,GA在萌发阶段开始起主导作用,这种作用一直持续到形成完整植株。在芽分化及生根的过程中,TDZ也成为决定芽分化的主要因素,此时活性炭变为一种次要因素。本研究取材的时间是9月初,未成熟胚中萌发抑制物尚少,是本研究较高的未成熟胚萌发率得以实现的前提条件,然而所取的珙桐果实在冰箱中放置时间超过两周后,就会造成种子生活力严重下降,未成熟胚萌发率严重降低。因此采用此法获得珙桐的再生植株,珙桐果实的保存是今后需要解决的问题。
通过本研究得到了芽诱导的最佳培养基0.09mg/L NAA+0.6 mg/L TDZ+0.9 mg/L GA+0.8 g/L AC+WPM培养基,此时未成熟胚萌发率为71.43%;芽分化及生根的最佳培养基为0.1 mg/L NAA+3 mg/ LTDZ+6 mg/L GA+1.2g/L AC+WPM培养基,此时再生植株的比例为50%。通过极差分析,得到了芽诱导阶段的最佳培养基组合,然而由于材料的限制未能得到验证。
邹利娟等采用组织培养的方法成功的繁殖珙桐苗,且幼苗成活率高达60%[11],因此作为快繁技术未成熟胚再生植株因为成本高,取材时间受限,及材料保存困难受到限制。然而本实验的结果却为研究珙桐胚的形态后熟和生理后熟提供了良好的材料。激素及萌发抑制物质的对胚的萌发及芽分化和生根的调控有待进一步的研究。
注释及参考文献:
[1]傅立国.中国植物红皮书(第一册)[M].北京:科学出版社.1991:474-475.
[2]贺金生,林洁,陈伟烈.我国珍稀特有植物珙桐的现状及其保护[J].生物多样性.1995,3(4):213-221.
[3]陈坤荣,李桐森,田广红,等.珙桐繁殖的生物学特性[J].西南林学院学报,1998,18(2):8-13.
[4]万朝琨.珙桐种子休眠的解剖学研究[J].中南林学院学报,1988,8(1):35-39+114-115.
[5]余阿梅,苏智先,王立强,等.珍稀濒危植物珙桐胚的萌发与快速繁殖[J].植物学报,2009,44(4):491-496.
[6]董社琴,李冰雯,王爱荣.植物生长调节剂对珙桐种胚离体培养的影响[J].湖北农学院学报2004,24(4):291-294.
[7]雷泞菲,苏智先,陈劲松,等.珍稀濒危植物珙桐果实中的萌发抑制物质[J].应用与环境生物学报,2003,9(6):607-610.
[8]苏智先,张素兰.珙桐种群生殖物候及其影响因子研究[J].四川师范学院学报(自然科学版),1999,20(4):4-9.
[9]雷泞菲,彭书明,牛蓓,等.珍稀濒危植物珙桐种子休眠萌发过程中内源激素的变化[J].广西植物,2009,29(1):66-69+131.
[10]刘承德,成磊.新型植物生长调节剂噻苯隆机理作用及应用[J].新疆农垦科技,2011,2:63-64.
[11]邹利娟,苏智先,胡进耀,等.濒危植物琪桐的组织培养与植株再生[J].植物研究,2009,29(2):187-192.
In Vitro Embryo Germination and Plant Regeneration from Immature Embryos of Davidia Involucrata Baill
FANG Zhi-rong,CAI Guang-ze
(Xichang College,Xichang,Sichuan 615013)
In order to obtain Davidia involucrata generation system,the immature embryos of D.involucrata were taken as explants,and regenerated plants were obtained.Four factors(α-Naphthaleneacetic acid[NAA], Thidiazyron[TDZ],gibberellic acid[GA]and active carbon[AC]affecting plant regeneration from immature embryos of D.involucrata were studied by use of orthogonal design and analyzed by range analysis.The results showed that: the best combination medium for embryo germination was WPM+0.09 mg/L NAA+0.6 mg/L TDZ+0.9 mg/L GA+1 g/L AC,and the germination rate were 71.43%.While the best combination medium for embryos grew into shoots and rooted was WPM+0.1 mg/L NAA+3 mg/L TDZ+6 mg/L GA+1.2 g/L AC,and the generation rates was 50%. AC could promote turning green of hypocotyls and cotyledon.NAA and GA were important to break after-ripening of embryo.GA and TDZ played a leading role in growing shoots and roots.
Davidia involucrate Baill;immature embryo;plant regeneration
Q943.1
A
1673-1891(2015)01-0007-04
2014-10-15
四川省教育厅项目(项目编号:08zb015);西昌学院院自立项目(项目编号:ZZSSA0808)。
方志荣(1981-),女,讲师,在读博士,研究方向:植物学。
