结瘤信号途径中相关调控蛋白的研究进展
2015-08-09柯丹霞李祥永
柯丹霞 ,李祥永
(信阳师范学院 生命科学学院,河南 信阳464000)
0 引言
宿主豆科植物与根瘤菌之间的共生固氮是自然界中最主要的一种生物固氮形式.根瘤器官的形成是豆科植物与根瘤菌之间相互识别,信号分子之间相互作用的结果.根瘤菌首先感知豆科植物根际分泌的类黄酮信号,随后激活结瘤基因的表达,合成和分泌结瘤因子,宿主植物体内启动一系列早期结瘤反应:根毛顶端卷曲,形成牧羊拐结构;根瘤菌在卷曲根毛顶端增殖,侵入线形成;根皮层细胞迅速分裂,形成根瘤原基;侵入线内的根瘤菌释放,进入根瘤原基.根瘤菌侵染和根瘤器官发生这两个过程协同发展最终形成有固氮功能的根瘤[1].
豆科植物与根瘤菌共生关系的建立极为复杂,但有序.通过对豆科植物不结瘤突变株的研究,结瘤早期信号转导途径开始逐步明晰.早在1985年,Carrol和Gresshoff等人就通过EMS化学诱变的方法从大豆中分离到超量结瘤和不结瘤突变株[2],接着在其他豆科植物如豌豆、三叶草,以及模式豆科植物百脉根和苜蓿中也获得超量结瘤和不结瘤突变株.自2002年以来,随着两大模式豆科植物百脉根(Lotusjaponicus)和蒺藜苜蓿(Medicagotruncatula)中关键共生调控基因的陆续发现,结瘤早期信号转导模型初步建立起来,见图1[3].目前单一的对某个蛋白在共生互作中功能的研究已经达到饱和,以此结瘤信号转导模型为基础,检测信号途径中已知蛋白间的相互作用,确定蛋白质相互作用的活性位点,寻找与已知蛋白相互作用的新蛋白,从生化和分子水平上进一步研究蛋白调控网络已成为当今共生固氮领域新的研究热点.近年来,许多与已知关键调控蛋白互作的新蛋白已经从模式豆科植物中被筛选出来,并且这些蛋白也被证实在根瘤共生过程中发挥重要作用,结瘤早期信号转导途径因此得到进一步的补充和完善.本文以根瘤共生为视角,主要介绍参与早期共生信号转导的关键调控蛋白特别是与已知蛋白互作的新蛋白的最新研究进展,以期为豆科植物与根瘤菌共生关系的研究提供参考.
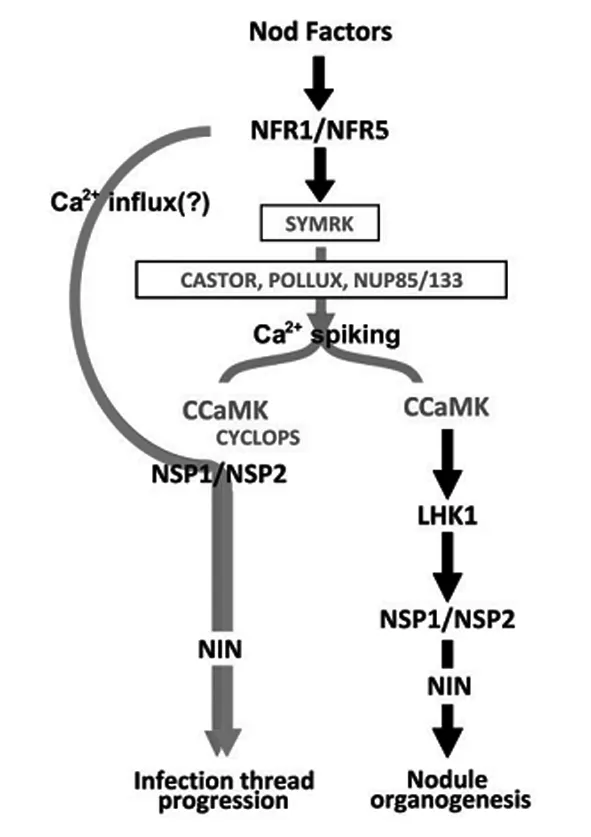
图1 结瘤信号传导模式图Fig. 1 Nodulation signaling transduction pattern.
1 结瘤因子受体激酶及其互作蛋白
不同种类豆科植物的结瘤因子受体蛋白激酶陆续被筛选出来:包括百脉根中的LjNFR1和LjNFR5[4-5];苜蓿中的MtLYK3/MtLYK4和MtNFP[6-8];豌豆中的PsSYM37和PsSYM10[9]以及大豆中的GmNFR1α/β和GmNFR5α/β[10].不同豆科植物中鉴定到的结瘤因子受体被统一归为LysM类受体激酶(LysM-RLKs),由膜内的丝/苏氨酸蛋白激酶结构域、中部跨膜结构域和膜外2~3个LysM结构域共同组成.
在大多数真核生物的蛋白激酶中,调节激酶活性的激活环位于蛋白的激酶结构域,而磷酸化位点一般位于激活环(activation loop).模式豆科植物百脉根的结瘤因子受体蛋白激酶LjNFR1和苜蓿的结瘤因子受体蛋白激酶MtLYK3均含有激活环,因此具有蛋白激酶活性.另外的两个结瘤因子受体蛋白激酶LjNFR5和MtNFP因为缺少激活环而不具备磷酸化活性.体外实验也证实LjNFR1是典型的蛋白激酶,具有自磷酸化和底物水平磷酸化活性.LjNFR1通过磷酸化LjNFR5,将结瘤因子信号传递到下游.与此同时,LjNFR1和LjNFR5可以在烟草和韭葱细胞内形成异源二聚体[11],并且LjNFR1和LjNFR5能够高亲和性地直接结合结瘤因子(NF)[12].最新研究发现LjNFR5的LysM2结构域结合NF,由此引起LysM2结构域构象发生变化[13].这些生化上的直接证据进一步证实结瘤因子受体蛋白激酶可能以异源二聚体的形式与NF相结合,共同参与接收NF信号,然后通过两个激酶结构域之间的转磷酸作用传递NF信号,见图2[1].
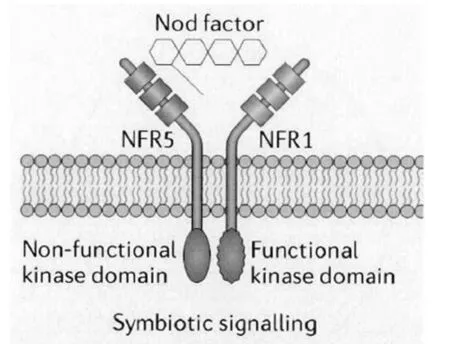
图2 NFR1和NFR5共同识别结瘤因子Fig. 2 Perception of nod factors by NFR1 and NFR5.
NFR5如何将NF信号传递到下游,这一过程是否需要新的蛋白参与.笔者前期利用酵母双杂交技术在百脉根中筛选到与NFR5相互作用的小G蛋白ROP6.证实NFR5与ROP6在植物细胞的细胞膜相互作用,ROP6基因的表达依赖于根瘤菌的侵染,其RNA干扰实验导致侵染线形成和根瘤数目下降,表明ROP6作为正调节子参与根瘤菌对豆科植物的侵染和根瘤的发育[14].ROP作为分子开关,有着极其复杂和保守的调控机制,ROP与RopGAP、RopGEF和RopGDI作为一个调控体系共同发挥功能.因此,为了阐明ROP6对其互作蛋白NFR5的调控机制,下一步必须寻找ROP6相对应的调节子RopGAP、RopGEF和RopGDI,即将ROP类小G蛋白介导的信号通路引入共生信号转导途径中.
在苜蓿中利用酵母双杂交技术也筛选到MtLYK3(LjNFR1的同系物)的相互作用蛋白PUB1.生物信息学分析表明PUB1含有植物保守的U-box结构域,体外蛋白实验证实PUB1具有E3泛素连接酶活性.MtLYK3的激酶结构域能够磷酸化PUB1,二者共定位于植物的细胞膜.PUB1的超表达和RNAi实验显示PUB1负调控根瘤菌的侵染过程,是共生信号转导途径的一个新的负调节子[15].此外,苜蓿中的MtSYMREM1能够分别与MtLYK3与MtNFP互作,MtSYMREM1属于Remorin蛋白,是一个新发现的与质膜/脂筏相连的植物专一性蛋白家族.对MtSYMREM1的表达谱和功能缺失突变体研究发现MtSYMREM1参与调控侵染线的形成.MtSYMREM1在质膜通过调控受体蛋白参与侵染早期过程,影响侵染线的极性生长以及根瘤菌的释放[16].随后,在百脉根中也克隆到MtSYMREM1的同系物LjSYMREM1.LjSYMREM1能够与受体蛋白在体内瞬时互作,在体外LjSYMREM1的N-端结构域能够被受体蛋白磷酸化.进一步的生物学功能研究证实其在根瘤菌侵染中发挥作用[17].
综上,一些与结瘤因子受体蛋白互作的新蛋白被发现,并且越来越多的蛋白也被证实调控根瘤菌早期侵染过程,但是这些蛋白与结瘤因子受体蛋白是如何协作共同调控早期侵染过程的,以及这些蛋白之间的网络调控关系是怎样的目前仍不清楚,还需要将来更深入和细致的研究来揭示和阐明.
2 共生受体激酶及其互作蛋白
最初在百脉根中克隆到的共生受体激酶基因是SymRK(symbiosis receptor kinase)[18],其编码富含亮氨酸重复序列的类受体激酶(LRR-RLK).SymRK定位于质膜和侵入线膜上.在蒺藜苜蓿中的同源基因为DMI2 (do not make infections),紫花苜蓿中为NORK(nodulation receptor kinase)[19].
通过酵母双杂交的方法,以MtDMI2的蛋白激酶结构域为诱饵,在苜蓿中筛选到甲羟戊酸合酶HMGR1.甲羟戊酸是植物类固醇类激素、细胞分裂素等特定类异戊二烯类物质的重要前体,而研究人员筛选到的HMGR1蛋白能够催化甲羟戊酸这一重要物质的合成.基因沉默实验表明HMGR1参与根瘤菌的侵染及根瘤器官发生两个过程[20].在百脉根中通过酵母双杂交的方法,以LjSymRK的蛋白激酶结构域为诱饵,相继筛选到ARID型DNA绑定蛋白LjSIP1,E3连接酶LjSINA4和LjSIE3,以及MAP激酶激酶LjSIP2.LjSIP1为一个具有DNA结合功能的转录因子,主要和富含AT的序列结合(ARID),并且能特异结合起始结瘤基因NIN的启动子序列.LjSIP1蛋白在细胞中定位于细胞核,它并不是SymRK蛋白的磷酸化底物[21].后续的研究发现了SIP1的变异体SIP1L,SIP1L 在C端含有额外的17个氨基酸.与SIP1不同的是,SIP1L不能够与SymRK互作.SIP1L和SIP1能够形成异源二聚体.SIP1-RNAi转基因植株抑制结瘤,超表达SIP1L和 SIP1都能够增加根瘤数目,说明SIP1正调控结瘤过程[22].Wang (2013)等推测在进化过程中SIP1L在百脉根中的功能丧失,由较短的剪接变体SIP1与SymRK互作将共生信号传递到下游.此外,研究发现来自两个不同家族的E3泛素连接酶LjSINA4和LjSIE3均能与SymRK蛋白互作.LjSINA4负调控根瘤菌的侵染过程,而与之相反的是LjSIE3正调控根瘤菌的侵染过程.异位表达LjSINA4导致侵染线不能正常发育,SymRK的蛋白表达也受到抑制[23].LjSIE3在植物体内具有介导SymRK泛素化修饰的功能,而不能介导SymRK蛋白泛素化降解.基因超表达和RNA干扰研究表明,LjSIE3正调控根瘤菌侵染过程[24].两种不同的E3泛素连接酶虽然都能与SymRK蛋白互作,但是由于作用机制不同,导致两种蛋白在调控根瘤菌侵染过程中功能相反.MAP激酶激酶参与植物体内多种生理活动,在百脉根早期结瘤信号途径中,也筛选到一个MAP激酶激酶LjSIP2能够与SymRK相互作用.体外磷酸化实验表明SIP2和SymRK不能互为磷酸化底物,但是SymRK能够负调控SIP2的激酶活性.RNAi分析表明,SIP2表达量的减少导致侵染线和根瘤原基数目明显减少[25].Remorin家族蛋白LjSYMREM1和MtSYMREM1也能够分别与共生受体激酶SymRK和DMI2互作,推测其在共生过程中可能作为支架蛋白从空间上参与调控信号传导复合物的装配[16-17].
上述的研究结果说明,共生受体激酶可能与这些蛋白之间形成复杂的蛋白网络共同调控豆科植物与根瘤菌的共生过程.Chen等(2012)首次在根瘤共生信号途径中发现MAP激酶的踪迹,该研究补充和完善了MAPK信号传递网络,也为共生固氮领域的研究提供了新的视角.水稻OsRac1(LjROP6的同系物)通过OsMAPK3/6 级联调控下游转录因子RAI1,从而将天然免疫信号传递到下游[26].由此推测,笔者前期鉴定的LjROP6是否也与MAPK级联成员LjSIP2存在一定的联系,从而通过NFR5→ROP6→SIP2信息流将共生信号传递到下游,这个推测还需要大量的实验数据来证实.此外,植物体内为什么存在三个受体激酶,结瘤因子受体激酶与共生受体激酶之间的关系又是怎么样的也是研究人员一直困惑并试图解决的难题.
3 钙离子与钙调素依赖的蛋白激酶及其互作蛋白
CCaMK编码钙离子和钙调素依赖的蛋白激酶[27].生化分析表明CCaMK受Ca2+和CaM双调控,使得CCaMK能够识别复杂的钙离子信号,是解码共生信号传导中Ca2+振荡信号的主要候选蛋白.CCaMK参与根瘤菌侵染过程,在根瘤器官发生过程中也发挥重要调控作用[28].2008年,研究人员利用酵母双杂交技术,在百脉根中筛选出一个新的蛋白CYCLOPS与CCaMK相互作用,在苜蓿中也鉴定出一个新蛋白IPD3[29-30].CYCLOPS/IPD3是CCaMK的磷酸化底物,CCaMK通过磷酸化CYCLOPS/IPD从而参与调控根瘤菌侵染过程,但是并不影响根瘤的器官发生[30].2011年,研究人员在百脉根中鉴定到一个CCaMK相互作用的新蛋白CIP73.体外蛋白实验证实在Ca2+和CaM存在时,CIP73的N端1~413位氨基酸区域能够被CCaMK磷酸化.RNAi分析表明CIP73参与根瘤的器官发生过程,而不影响根瘤菌的侵染[31].上述研究表明,CCaMK能够通过与不同的蛋白相互作用分别激活根瘤菌侵染和根瘤器官发生两个过程.
4 NSP转录因子及其互作蛋白
在百脉根和苜蓿中,NSP1和NSP2都属于GRAS(GAI、RGA、SCR)家族蛋白[32-33].这两个转录因子参与调控结瘤因子诱导的早期结瘤基因表达、根瘤形成等过程.生化研究表明,LjNSP2 能够与自身结合形成同源二聚体[34].苜蓿中NSP1和NSP2在细胞核内形成异源蛋白复合体,并且能够结合ENOD11、 NIN 和ERN等早期结瘤素基因的启动子[35].NSP1与NSP2结合可以作为强调节子正调控ERN1和ENOD11的翻译.在根瘤菌侵染前期,ERN1专一性地激活由NF诱导的ENOD11的表达,在随后的根瘤菌侵染过程中,NSP1/NSP2介导ENOD11表达[36].
研究人员试图通过筛选酵母双杂交文库鉴定与NSP转录因子相互作用的新蛋白.最新的研究显示,一个MYB类转录因子能够特异的与NSP2相互作用,命名为IPN2.IPN2具有转录自激活活性,并且能够结合NIN基因的启动子.在酵母中进一步确定两种蛋白相互作用的活性位点发现NSP2的GRAS结构域和IPN2的卷曲-卷曲结构域是二者相互作用所必需的关键作用区段.利用百脉根毛根转化技术发现NSP2和IPN2共定位于毛根细胞的细胞核中.RNA干扰实验表明IPN2在根瘤器官发生过程中行使重要功能[37].IPN2的发现将MYB类转录因子引入共生信号转导途径中,进一步补充和完善了豆科植物与根瘤菌的早期信号转导途径.
5 结瘤起始蛋白NIN及其相关蛋白
结瘤起始蛋白NIN是一个转录因子,含有一个DNA结合结构域和一个跨膜结构域.研究表明,NIN不影响结瘤因子的感知,根毛的变形等早期反应,但NIN参与侵染线的形成,控制结瘤数量,推测其可能通过调控早期结瘤素基因ENOD11的空间表达从而发挥功能[38].两个核因子LjNF-YA1和LjNF-YB1作为NIN的转录靶物,二者在根瘤原基中表达,在植物细胞中形成NF-Y蛋白复合物.LjNF-YA1敲除实验抑制根瘤器官发生.超表达NIN,在不接种根瘤菌的情况下,皮层细胞也能够分裂形成类似根瘤原基的结构.豆科植物根瘤的发育与NIN 功能的变化紧密相连,NIN的一个重要功能就是调控根瘤器官发生过程中的细胞分裂[39].NIN是结瘤起始关键蛋白,在豆科植物中特异存在,而NF-Y在真核生物中普遍存在.
最新的研究又发现了两个在根瘤器官发生过程中行使功能的调控蛋白LjIPT3正调控结瘤过程,RNAi植株侵染线和结瘤数目减少.LjIPT3介导的CRE1依赖的细胞分裂素途径参与调控结瘤过程,LjIPT3调控结瘤起始和发育,不参与早期侵染事件[40].LjVAG1介导的皮层细胞核内复制是引导共生根瘤菌进入宿主植物分生组织细胞所必需的,因此LjVAG1也在根瘤器官发生过程中发挥作用[41].
6 结语
近10年来,豆科植物与根瘤菌共生固氮的分子机理研究取得了重大进展.利用模式豆科植物遗传资源,研究人员鉴定出大量与结瘤相关的关键基因.但是基因产物的确切生物学功能以及调控机制还不甚明晰;通过正向遗传学方法从传统诱导得到的共生突变体中分离基因似乎也已经达到饱和;当前的研究依然停留在鉴定涉及共生信号途径某一阶段的功能上.因此,如何将这些已知调控蛋白从生化上彼此联系起来,明确蛋白与蛋白之间的时空调控,上位/下位关系,筛选新的相互作用蛋白,进而将这些蛋白与不同的共生过程包括共生信号识别、根瘤菌侵染、根瘤器官发生、根瘤形成以及有功能(固氮)共生体建立等相对应,从而建立一个完整的植物与微生物共生调控网络将是下一步亟待解决的问题.
豆科植物在环境及农业上的应用长期以来一直受到人们的广泛关注.在过去十年里,积累了大量的共生突变体和基因,随着转录组、蛋白质组、代谢组等分析方法的完善和大量创新工具的快速建立,不仅在模式豆科植物上,甚至在重要的豆科农作物大豆上将会有更多全新的有意义的研究成果诞生.更深远的是人们期望能够通过遗传工程的方法将豆科植物与根瘤菌之间的这一复杂但精细的共生调控网络转移到非豆科粮食作物上,实现重要谷类作物的共生固氮.随着技术的不断创新和新资源平台的出现, 我们有理由相信实现非豆科植物的根瘤共生已经成为可能.
