香榧叶片抗寒性1)
2015-08-08刘千玲赵燕刘桂华
刘千玲 赵燕 刘桂华
(安徽农业大学,合肥,230036)
低温伤害是一种全球性的自然灾害。低温会影响植物的生长代谢,使植物受到伤害导致减产,严重时甚至会导致植物死亡。因此,对植物的抗寒性进行研究,并采取一定方法提高植物的抗寒能力,可在一定程度上降低低温伤害,使植物正常生长,提高产量。香榧(Torreya grandis)为常绿乔木,是我国特有的集食用、油用、药用、材用和观赏于一体的珍贵树种。在香榧的栽培历史中,人们积累的生产经验和研究成果主要集中在种质资源、丰产培育技术、生长发育特性和种子的商品加工技术等方面[1-3]。对于香榧丰产培育的生理学基础,特别是对香榧扩大培育区的光热生理生态学基础的研究报道较为少见。文中通过定位采样和室内测定,对一年中不同时期和不同树龄香榧的叶片抗寒特性进行了研究,为香榧优质丰产培育和产区区划提供理论依据。
1 试验地概况
试验地处于皖南盆地中心的徽州区,位于北纬29°30'25″~30°09'10″,东经 118°04'10″~118°53'50″。以山地丘陵为主,地势北高南低。属亚热带季风气候,年均气温16℃,极端最高气温40.8℃,极端最低气温-12.7℃,无霜期230 d,≥10℃年积温4 950℃。全年日照时间1 960 h。年平均降水量1 600 mm,其中4—10月份占全年70%。试验地土壤为黄红壤,土层厚度70 cm以上,有机质平均质量分数 2.3%,pH 值 5.5~6.0。
2 材料与方法
选择生长立地相同、树龄不同(3、6、8、12、18、22、27年生)的健康香榧树为试材,分别于2013年1、2、3、10、11、12 月份在不同树龄被试植株的 4 个方向分别选择生长健康的枝条各3枝,采集各枝条10 cm顶梢段的全部叶片,将四个方向12枝枝条叶片充分混合后分成3等份(重复3次),用潮湿纱布包裹分别装入密封塑料袋,带回实验室,分别用自来水将叶片冲洗干净,再用去离子水漂洗3次放在滤纸上吸干,再分别将每份叶片分为6组放入密封塑料袋置于超低温冰箱进行低温处理,温度设定梯度为0、-3、-6、-9、-12、-15 ℃,每 2 个温度间降温时间30 min,并分别在各个处理温度持续24 h[10]。香榧树属常绿树种,3—5月份为抽梢萌发新叶期,新叶充分伸展后在其余月份进行光合作用,为保证叶样的相对一致性,本试验所采叶片均为梢头充分伸展的叶片。
相对电导率的测定和半致死温度的计算。分别称取低温处理的叶片0.5 g(重复3次),放入盛有20 mL去离子水的烧杯中,震动摇匀浸泡24 h,用雷磁DDS—237电导仪测定电导率,再放入100℃沸水浴中煮30 min,冷却至室温后摇匀再用电导仪测定电导率,相对电导率=(浸泡电导率/煮沸电导率)×100%;根据不同低温处理下的相对电导率变化与温度的对应关系,利用 LogisticY=K/(1+ae-bx)拟合,(其中,Y为低温处理下的相对电导率;x为处理温度;K、a、b均为参数,K为Y的最大极限值,a表示曲线对原点的相对位置,b反映了x与Y之间的对应关系,当b>0时,Y值随着x的增加而增加,当b<0时,Y随x的增加而减少)求其方程的二阶导数,并令其为零,则可获得曲线的拐点X=(lna)/b,即半致死温度 LT50[8-9];丙二醛质量摩尔浓度测定采用硫代巴比妥酸法[15];可溶性糖质量分数测定采用蒽酮比色法[15]。
3 结果与分析
3.1 叶片半致死温度
从表1看出,一年中不同时期香榧叶片的半致死温度在各个树龄阶段都呈现基本相同的变化规律,即10月份的半致死温度值显著高于其他时期,而1月份的半致死温度值则显著低于其他时期。11月份与3月份的半致死温度和12月份与2月份的半致死温度,分别两两相比较无显著差异,但第一组(11、3月份)的半致死温度则显著高于第二组(12、2月份)的对应值。可见,一年中的不同时期,香榧叶片的抗寒能力存在显著差异,其由高到低的顺序为1、2、12、11、3、10 月份。
从表1还可以看出,一年中的相同时期,不同树龄阶段的香榧叶片半致死温度存在差异。其中1、2、12月份的叶片半致死温度在不同树龄树体之间的差异最为显著。从表2可见,各个时期香榧叶片半致死温度与其树龄之间存在相同的回归模型,即二次函数关系。通过对回归方程的极值求解(表2)可以看出,一年中的不同时期半致死温度与其树龄存在不同的对应关系,但其变化规律具有相似的趋势,即在一定的树龄范围内,树龄越大,其叶片半致死温度越低,但达到一定树龄后,叶片半致死温度则随树龄增大而变化缓慢或略有升高。综合不同时期测定结果可知,香榧树体叶片的半致死温度达到最低值的树龄为17~21 a。可见,树龄17~21 a可能是香榧抗寒能力最强的时期。
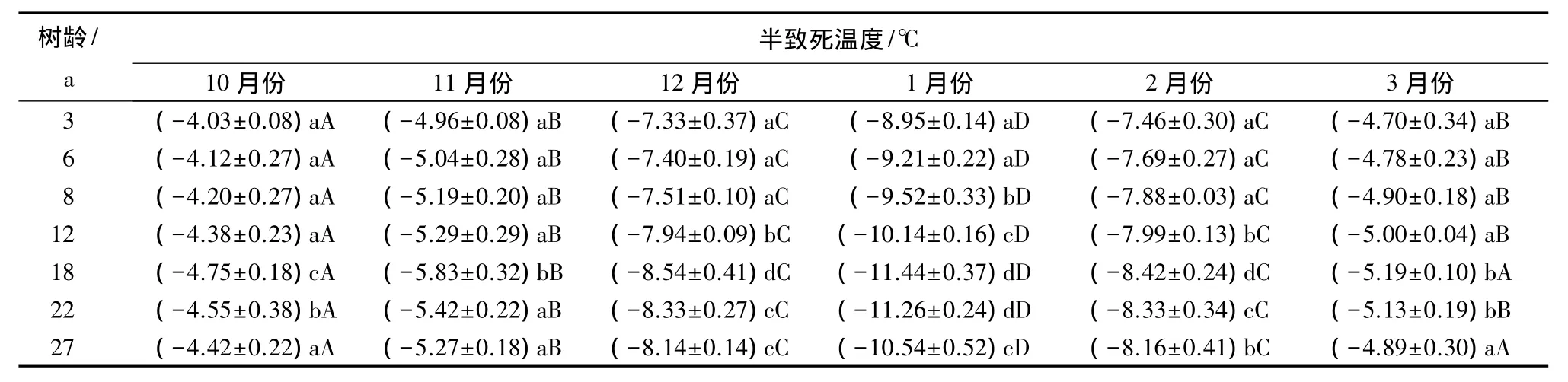
表1 香榧叶片半致死温度变化
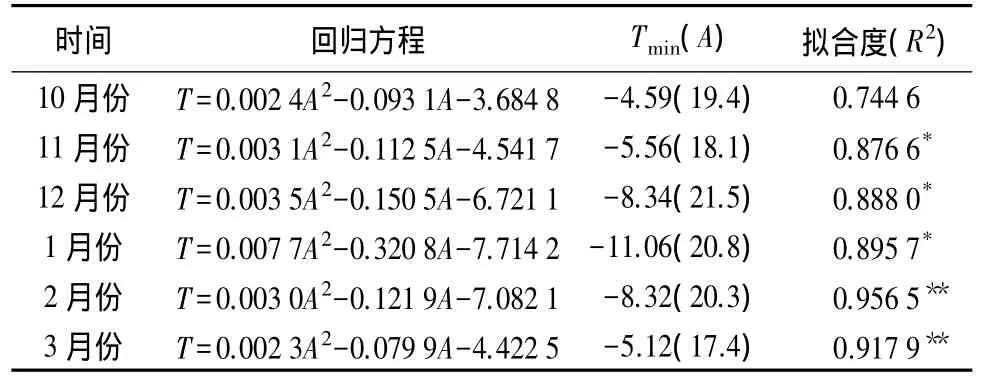
表2 一年中的不同时期半致死温度(T)与树龄(A)的回归方程
3.2 叶片丙二醛质量摩尔浓度
表3表明,一年中的不同时期,各个树龄阶段的香榧叶片丙二醛质量摩尔浓度由高到低的顺序基本上表现为1月份、2月份、12月份、11月份、3月份、10月份,且其差异程度均达到显著水平,与其同期半致死温度基本呈现相反的变化规律。
各个树龄阶段的香榧叶片丙二醛质量摩尔浓度在一年中的各个时期分别存在不同的变化规律,其中3月份和10月份的各个树龄阶段的香榧叶片丙二醛质量摩尔浓度不存在显著差异,而一年中的其他时期,香榧叶片丙二醛质量摩尔浓度大体可以分3~12 a和12~27 a两个树龄阶段,其中第一树龄阶段(3~12 a)的丙二醛质量摩尔浓度显著高于第二树龄阶段(12~27 a),而两组内部叶片的丙二醛质量摩尔浓度则均不存在显著差异。
植物组织丙二醛质量摩尔浓度与其受低温危害程度存在密切关系,丙二醛质量摩尔浓度越高,表明受低温危害的程度越重。由此可见,在本试验区,1月份香榧叶片受低温危害的响应最明显,其次是2月份和12 月份,在冬季(1、2、1、11 月份),3~12 a树龄的香榧叶片受低温的胁迫程度明显大于12~27 a树龄的香榧叶片。
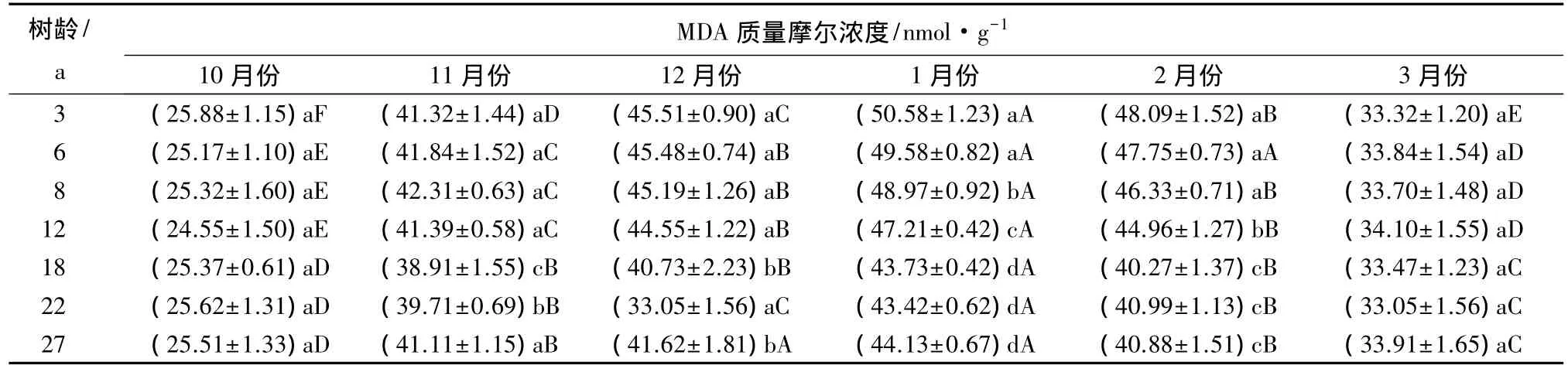
表3 香榧叶片丙二醛(MDA)质量摩尔浓度变化
3.3 叶片可溶性糖质量分数
由表4显示,一年中的不同时期,各个树龄阶段的香榧叶片可溶性糖质量分数存在不同程度的差异,其中,3月份和10月份显著低于一年中的其他时期,其他时期各个树龄阶段的香榧叶片可溶性糖质量分数由高到低的顺序则为1月份、12月份、2月份、11月份,且其差异程度均达到显著水平。
一年中的相同时期,不同树龄叶片的可溶性糖质量分数分别存在不同的变化规律,其中3月份和10月份,叶片可溶性糖质量分数在不同树龄之间不存在显著差异,而一年中其他时期不同树龄叶片的可溶性糖质量分数大体可以分为3~12 a和12~27 a两个树龄阶段,其中第一树龄阶段(3~12 a)的可溶性糖质量分数显著低于第二树龄阶段(12~27 a)的可溶性质量分数。这与丙二醛质量摩尔浓度有相同的变化规律。
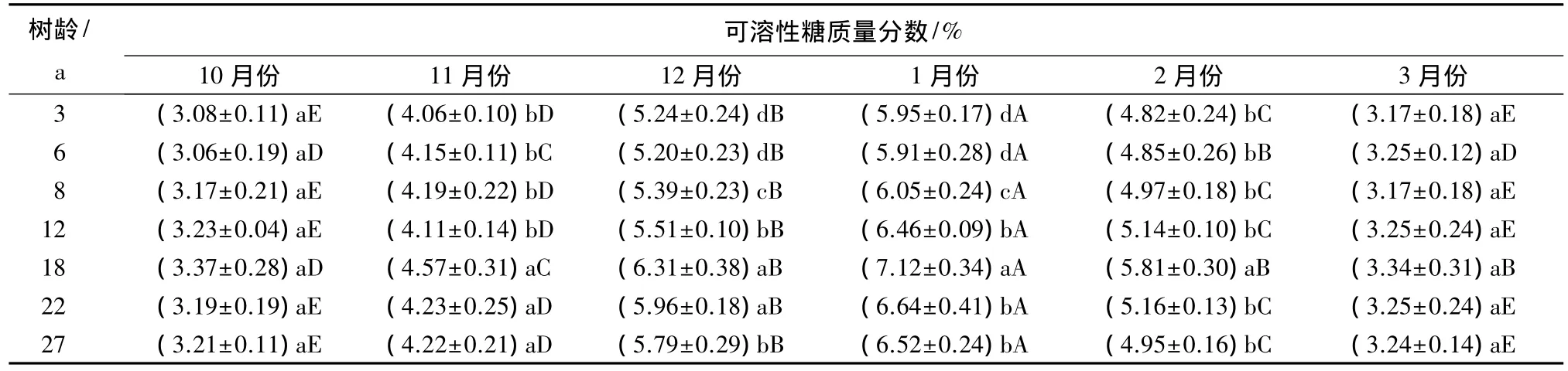
表4 香榧叶片可溶性糖质量分数的变化
3.4 叶片半致死温度与可溶性糖质量分数的相关性
依据香榧叶片不同季节半致死温度与其同期可溶性糖质量分数的相关性分析可知,11月份至翌年2月份,香榧叶片半致死温度与其可溶性糖质量分数的相关性均达到极显著相关(依次为-0.923 0**、-0.968 4**、-0.952 0**和-0.915 7**),而此期也是香榧叶片半致死温度较低、抗寒性较强的季节,说明香榧叶片可溶性糖质量分数的高低是影响其抗寒能力的重要因子。
4 结论与讨论
植物组织半致死温度的高低是衡量其抗寒能力强弱的一项重要直接指标[7]。随着外界环境温度条件的变化,树种的抗寒能力也会产生相应的变化,特别是随着季节变化的外界低温的诱导可不同程度地提高树种的抗寒性[16-17]。从一年中的不同时期看,香榧叶片的半致死温度由低到高依次为1月份、2月份、12月份、11月份、3月份、10月份,且其半致死温度最低值(1月份)与半致死温度最高值(10月份)相差5℃左右。半致死温度最低值与试验区最冷月平均温度出现的时间相吻合,说明在试验区最冷月(1月份)香榧的抗寒能力达到了最高水平。入秋以后,随着气温的降低,树体生理代谢逐渐诱导运输、转化和积累更多的可溶性糖等内含物于叶片中,从而提高了叶片的抗寒能力,这是香榧适应试验区温度变化的生理响应,与其他学者对相关树种的研究结论一致[4-7]。
植物组织丙二醛质量摩尔浓度的高低是其在逆境下受害程度的重要体现[7-9]。从测定结果看,香榧叶片丙二醛质量摩尔浓度由高到低的年变化与试验区冬季气温由高到低的月份呈完全相反的吻合关系,说明试验区冬季的低温对香榧叶片产生了明显的胁迫。不同树龄叶片丙二醛质量摩尔浓度的差异表明,树龄为17~21 a的香榧叶片受到的低温胁迫较轻,而树龄在12 a以下的香榧幼树叶片受到的低温胁迫较重。可见,不同树龄其抗寒能力存在差异,这可能是香榧在幼年时期生理代谢较弱,叶片适应低温而转化提高其可溶性糖等内含物的能力较差,因此,其抗寒能力较低。所以,做好幼树的防寒管理和抗寒锻炼是培育香榧丰产林的一项重要措施。
提高植物组织可溶性糖质量分数是提高其抗寒能力的重要生理机制[7-9]。香榧叶片可溶性糖质量分数的年变化与气温的年变化相关,特别是与试验区冬季的气温呈现完全相反的吻合关系,这是香榧叶片受低温诱导而降低细胞冰点减轻低温危害的生理响应[13-14]。
香榧叶片可溶性糖质量分数在试验区最冷月最高,而此期的半致死温度最低,表明可溶性糖是香榧叶片提高抗寒能力的重要机制之一,而丙二醛质量摩尔浓度在试验区最冷月最高,表明试验区最冷月的低温对香榧叶片产生的受害反应仍然最重。比较其他树种的半致死温度可以看出[5-6,12],香榧属于喜温树种。可见,抗寒锻炼和冬季防寒是培育香榧优质丰产林的重要环节之一。
[1] 万小金,吴晓明,程小东.香榧采穗圃营建技术[J].江西林业科技,2006,23(3):23-25.
[2] 黎章矩,程晓建,戴文圣,等.香榧品种起源考证[J].浙江林学院学报,2005,22(4):443-448.
[3] 戴文圣,黎章矩,曹福亮,等.我国榧树研究文献分析[J].浙江林业科技,2006,26(4):51-56.
[4] 张雪,刘鹏,王兴华,等.喜树种源间抗寒能力比较[J].东北林业大学学报,2011,39(1):14-16.
[5] 谢晓金,郝日明.南京地区12种常绿阔叶树冬季抗寒性动态变化[J].生态学报,2009,29(4):2149-2154.
[6] 张家洋,郭辉,张莉,等.不同低温胁迫对南京10种道路绿化树木形态及电导率的影响[J].东北林业大学学报,2012,40(5):25-28.
[7] 王宁,吴军,夏鹏云,等.大叶冬青对低温胁迫的生理响应及抗寒性分析[J].华南农业大学学报,2011,32(3):82-86.
[8] 王玲,王春雷,马喜娟,等.锦带花新品种抗寒性[J].东北林业大学学报,2012,40(12):43-46.
[9] 芦建国,杨金红,武翠红.山东地区引种的5种石楠属植物抗旱性比较[J].南京林业大学学报:自然科学版,2008,32(5):153-156.
[10] 刘艳萍,朱延林,康向阳,等.不同类型广玉兰的抗寒性评价[J].林业科学,2013,49(6):178-183.
[11] 刘辉,张钢.短日照对金叶女贞茎抗寒性和电阻抗图谱参数的影响[J].华北农学报,2008,23(2):173-179.
[12] 田如男,薛建辉.6个常绿阔叶乔木树种抗寒性研究[J].西南林学院学报,2005,25(4):110-112.
[13] Turner J,Mitchell S J.The effect of short day treatments on containerized Douglas-fir morphology,physiology and phenology[J].New Forests,2003,26:279-295.
[14] Mancuso S,Nicese F P,Masi E,et al.Comparing fractal analysis,electrical impedance and electrolyte leakage for the assessment of cold resistance in Callistemon and Grevillea spp.[J].Journal of Horticultural Science and Biotechnology,2004,79:627-632.
[15] 李合生,孙群,赵世杰.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[16] Beck E,Hem R,Hansen J.Plant resisstance to cold stress mechanisms and environmental signals triggering frost hardening and dehardening[J].Journal of Bioscience,2004,29:440-459.
[17] Kang S K,Motosugi H,Yonemori K,et al.Freezing injury to persimmons(Diospyros kaki Thunh)and four other Diopyros species during deacclimation in the spring as related to bud development[J].Scientia Horticulturae,1998,77:33-43.
