香蓼种子休眠与萌发特性1)
2015-08-08何淼常进刘娇
何淼 常进 刘娇
(东北林业大学,哈尔滨,150040)
香蓼(Polygonum viscosum)是蓼科(Polygonaceae)蓼属的一年生草本植物,主要分布在黑龙江省、吉林省、辽宁省、台湾地区,俄罗斯、朝鲜、日本、印度等地也有分布[1]。香蓼为一年生草本,植株具香味,总状花序呈穗状,花期 7—9月份,瘦果宽卵形[2]。香蓼作为蓼属植物中较为珍贵的芳香植物,目前处于野生状态且资源相对稀少。蓼属植物中部分种子存在休眠现象且不易萌发。其种子萌发是复杂的生理过程,亦是保证植物生长的重要阶段,环境条件中的温度、水分、光照、土壤及种子自身条件中的酶、激素等都能够影响种子的萌发[3]。对蓼科蓼属种子休眠原因的研究中表明:种皮障碍、胚形态发育不完全、化学抑制物质[4-6]等均可导致种子休眠。目前有关香蓼的挥发油[7]已有初步研究,但有关香蓼种子休眠与萌发特性还尚未见报道。本研究通过对香廖种子休眠与萌发特性进行定性试验,探索促进其萌发的条件,为深入探讨这种植物的生态适应机制提供依据,同时为生产中获得大量种苗,建立种植资源圃,以及为进行进一步的杂交育种及品种选育工作提供理论基础和技术支持,对香蓼的工厂化生产具有一定现实意义。
1 材料与方法
1.1 试验材料
2012年9月份于黑龙江省尚志市帽儿山(45°20'~45°20'N,127°30'~127°34'E)采集野生香蓼种子。空瘪粒采用水选法漂出,然后将饱满种子于室温下阴干后装入牛皮纸袋中,室温储藏备用。
1.2 香蓼种子休眠特性的测定
种皮机械障碍测定:将香蓼未破皮种子和破皮种子分别在蒸馏水中浸泡48 h,75%乙醇消毒30 s,0.1%HgCl2消毒 6 min,放入 MS+30 g·L-1蔗糖+8 g·L-1琼脂的培养基中。试验条件为(25±2)℃,光照强度 1 500~2 000 lx,光照时间16 h·L-1。每处理50粒种子,重复3次。计算发芽率。发芽率=(已发芽种子数/总粒数)×100%。
种皮透水性测定:未破皮种子和破皮种子称质量并浸泡于蒸馏水中,分别在 1、2、3、4、8、12、24、36、48 h时取出,用滤纸吸干种子表面水分称质量,直至种子质量不再变化为止。每处理50粒种子,重复3次,计算吸水率。吸水率=[(吸水后质量-吸水前质量)/吸水前质量]×100%。
1.3 香蓼种子萌发特性的测定
不同温度处理:将香蓼种子种皮划破后,200 mg·L-1GA3浸种 48 h,分别置于常温 5、15、20、25、30℃,5℃ /25℃(夜12 h/昼12 h)、20℃ /30℃(夜12 h/昼12 h)下进行发芽试验,光照强度为1900~2000 lx。
不同光照强度处理:将香蓼种子种皮划破后,200 mg·L-1GA3浸种 48 h,在 1 900~2 000 lx、400~500 lx、黑暗条件下进行发芽试验,温度为25℃。
不同酸蚀时间处理:分别用浓硫酸浸泡种子0、3、5、8、10 min 后用蒸馏水冲洗干净,用 200 mg·L-1GA3浸种48 h后进行发芽试验。温度为25℃,光照强度 1 900~2 000 lx。
不同质量浓度及种类的外源植物生长调节剂处理:用 GA3(100、200、300、500 mg·L-1)、6-BA(50、100、150、200 mg·L-1)、NAA(10、50、100 mg·L-1)处理未破皮种子,以蒸馏水浸泡48 h未破皮种子为对照;将香蓼种子种皮划破再经过上述处理,对照为破皮种子。温度为25℃,光照强度1 900~2 000 lx。
1.4 数据分析
采用Excel和SPSS19.0软件进行方差分析及多重比较分析(Duncan’s法)。
2 结果与分析
2.1 香蓼种子休眠影响因素
2.1.1 种皮机械障碍特性
相同光照条件下,在无菌培养基中未破皮种子不萌发,而破皮种子的萌发率为36.67%,最早萌发时间为第3 d。由此说明,香蓼种子的萌发受到种皮的机械阻碍作用,破皮种子更易萌发。
2.1.2 种皮吸水性测定分析
试验结果如图1所示。在吸水过程中,香蓼未破皮种子与破皮种子的吸水趋势大致相同,但就整个吸水过程而言,破皮种子的吸水率均略高于未破皮种子,在4 h之前比较明显,这说明破皮有助于香蓼种子快速吸水。在4 h之后吸水曲线呈缓慢上升趋势,饱和状态出现在36 h左右,在饱和状态下未破皮种子与破皮种子的吸水率差异不显著,吸水率分别为 25.01%、26.03%。
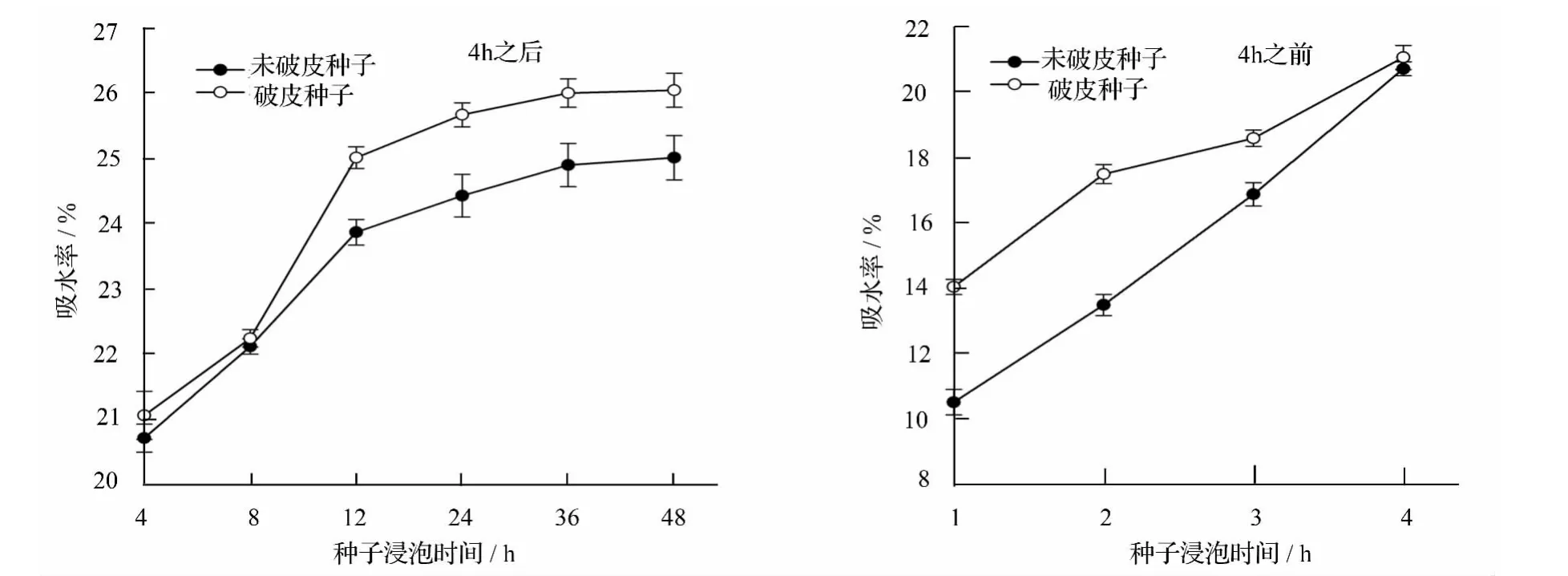
图1 香蓼种子吸水曲线
2.2 香蓼种子萌发特性
2.2.1 温度对种子萌发的影响
不同温度处理对种子发芽率、发芽势、平均发芽时间进行方差分析(表1),25℃条件下,种子发芽率和发芽势均为最高,分别达到58.00%和31.33%。这与其他6个温度处理下的结果相比,差异极显著(P<0.01);在25℃条件下,平均发芽时间最短,仅为4.36 d,说明种子发芽速度在25℃条件下最快,同时在5℃条件下香蓼种子几乎不萌发。
2.2.2 光照强度对种子萌发的影响
不同光照强度处理均在第2 d开始发芽(表2)。不同光照强度条件对香蓼种子发芽率、平均发芽时间影响差异不显著(P>0.05)。在光照强度为1 900~2 000 lx和黑暗条件下,平均发芽时间差异显著(P<0.05)。在光照强度为1 900~2 000 lx条件下,种子的发芽率和发芽势均最高,分别为58.00%和 31.33%,平均发芽时间最短,仅为 4.36 d。所以,香蓼种子在光照强度为1 900~2 000 lx时萌发效果最好。
2.2.3 不同时间浓硫酸处理对种子萌发的影响
经过不同时间浓硫酸处理后,香蓼种子的发芽率和发芽势与对照组相比,均差异显著(P<0.05)(表3)。发芽率和发芽势在不同处理时间下差异显著(P<0.01),但平均发芽时间并未达到显著水平(P>0.05)。随浓硫酸处理时间的延长,其发芽势呈现先升高后降低的趋势。在1 900~2 000 lx光照强度下经浓硫酸处理5 min时,其发芽率和发芽势均达到最高,其发芽率为 24.67%,较其他处理差异显著(P<0.05)。

表1 温度对种子萌发的影响
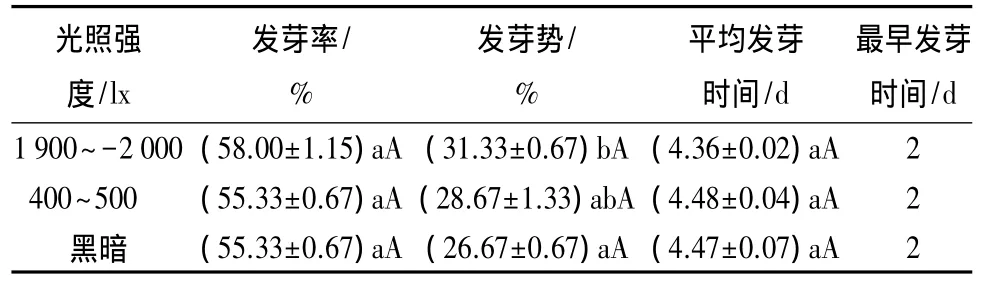
表2 光照强度对种子萌发的影响

表3 不同时间浓硫酸处理对种子萌发的影响
2.2.4 外源植物生长调节剂对种子萌发的影响
不同外源植物调节剂处理种子,其发芽率、发芽势均显著高于对照组,平均发芽时间均低于对照组(除 150、200 mg·L-16-BA 外)(P<0.05)(表4)。

表4 外源植物生长调节剂对破皮种子萌发的影响
随着GA3质量浓度上升,种子发芽率和发芽势先升高后降低,而平均发芽时间与之相反;随6-BA质量浓度上升,其发芽率和发芽势下降,而平均发芽时间则增大。随NAA质量浓度上升,其发芽率、发芽势、平均发芽时间均上升。在11种外源激素处理中,香蓼种子在200 mg·L-1GA3处理下,发芽率和发芽势最高,分别为58.00%和31.33%,与其他处理间差异极显著(P<0.01)。
3 结论与讨论
蓼属植物部分种子具有休眠特性,这种休眠特性和种子本身的性质有关,并且因植物种类及萌发条件差异而不同[8-9]。前期研究工作发现,香蓼种胚(胚根、子叶)不存在休眠现象,但其萌发需要营养作为动力。本研究发现香蓼种子虽具有较厚蜡质角质层,但种皮透水性较好,所以种子透水性不佳不是引起休眠的原因。在种皮障碍测定中发现,破皮种子发芽率为36.67%。而未破皮种子不萌发,因此,其不易萌发的原因之一是种皮对种子萌发的机械阻碍作用,使得种胚不能突破种皮正常生长。这与红蓼、核桃(Amygdalus persica)、杜仲(Eucommia ulmoides Oliv.)等植物存在的机械阻碍类似[10]。
温度是影响蓼属植物种子萌发的一项重要因素,其过高过低都不利于种子萌发[11-12]。本试验中香蓼种子在温度25℃下最适宜萌发,发芽率、发芽势分别为58.00%和31.33%,与其他6个试验结果相比差异极显著(P<0.01),同时变温条件未能有效提高种子发芽率,这说明温度与香蓼种子的萌发有着密切联系;在25℃下进行不同光照试验,结果表明,种子发芽率、平均发芽时间差异不显著,由此说明,光照不是影响香蓼种子萌发的主要因素;酸蚀处理是降低厚种皮种子硬实率常用的方法[4,13],本试验中处理时间为5 min时,其发芽率和发芽势均最高,且较其他处理差异显著,发芽率为24.67%。随着浓硫酸处理时间不断延长,种子发芽率先升高后降低,原因可能为在浓硫酸处理3~5 min时,种皮透性逐渐增强,而5 min后浓硫酸放热对种子萌发有影响,这说明浓硫酸处理对种子萌发有一定的促进作用;试验过程中发现,只有在破皮与外源激素同时处理香蓼种子时,种子才会萌发,这与本试验中影响种子休眠因素的研究结果相一致。在11组外源激素的处理中,经200 mg·L-1GA3处理的破皮香蓼种子,其发芽率、发芽势均最高,分别为58.00%和31.33%,较其他处理差异达到极显著水平(P<0.01)。这说明种子萌发需要动力,GA3等外源激素的使用,可以为香蓼种子的萌发提供所需的营养。GA3等外源激素的使用在某种程度上可能会解除某种抑制物质的抑制作用,从而促使种子萌发。
综上所述,香蓼种皮坚硬对种子萌发有一定的机械阻碍,是否还存在其他影响香蓼种子萌发的因素,如种皮中存在抑制物质等,还有待进一步研究。
[1] 聂绍荃,袁晓颖.黑龙江省植物资源志[M].哈尔滨:东北林业大学出版社,2003:215.
[2] 王雁,岳桦,汤一方.中国黑龙江野生花卉[M].北京:中国林业出版社,2008:40.
[3] 乌凤章,刘桂丰,姜静,等.种子萌发调控的分子机理研究进展[J].北方园艺,2008(2):54-58.
[4] 许桂芳,刘明久,席世丽,等.破除红蓼种子休眠研究[J].种子,2005,24(1):24-25.
[5] 曹贵荣,王建荣.GA3打破红蓼种子深休眠的试验[J].植物生理学通讯,1996,32(6):426-427.
[6] 沈法莱,杨大,杨跃雄,等.影响珠芽蓼种子发芽诸因素及其生态[J].资源开发与市场,1999,315(3):131-132.
[7] 张德志,杨文胜,张伟森,等.香蓼挥发油化学成分的研究[J].吉林林学院学报,1992,8(1):6-9.
[8] [美]卡恩·AA.种子休眠和萌发的生理生化[M].王沙生,译.北京:农业出版社,1989:33-68.
[9] 唐安军,龙春林,刀志灵.种子休眠机理研究概述[J].云南植物研究,2004,26(3):241-251.
[10] 杨朝霞.厚朴种子解除休眠及萌发生理的研究[D].南京:南京林业大学,2008.
[11] 孙昌高,方坚,徐秀瑛.蓼科药用植物种子发芽的研究[J].资源开发与市场,1999,15(3):131-132.
[12] Bouwmeester H J,Karssen C M.The dual role of temperature in the regulation of the seasonal changes in dormancy and germination of seeds of Polygonum persicaria L[J].Oecologia,1992,90:88-94.
[13] 史晓华,徐本美,黎念林,等.青钱柳种子休眠与萌发的研究[J].种子,2002,15(1):5-7.
