甘南牦牛CYGB基因的克隆及生物信息学分析
2015-07-12娄延岳杜晓华罗玉柱
娄延岳,杜晓华,罗玉柱,刘 霞,张 磊
(1 甘肃农业大学 a 生命科学技术学院,b 动物医学院,甘肃 兰州730070;2 甘肃省草食动物生物技术重点实验室,甘肃 兰州 730070;3 河南农业职业学院 动物科学系,河南 郑州 451450)
甘南牦牛CYGB基因的克隆及生物信息学分析
娄延岳1a,2,杜晓华1b,2,罗玉柱2,刘 霞1a,2,张 磊3
(1 甘肃农业大学 a 生命科学技术学院,b 动物医学院,甘肃 兰州730070;2 甘肃省草食动物生物技术重点实验室,甘肃 兰州 730070;3 河南农业职业学院 动物科学系,河南 郑州 451450)
【目的】 探讨甘南牦牛CYGB基因的分子结构特征及生理功能。【方法】 提取甘南牦牛血样基因组DNA,以其为模板PCR分段扩增后拼接得到CYGB基因,测序后进行生物信息学分析。【结果】 克隆出甘南牦牛CYGB基因,其长度为8 484 bp,含有4个外显子和3个内含子,外显子长度分别为143,232,164和34 bp,内含子长度分别为5 175,280和2 379 bp,可编码190个氨基酸残基,氨基酸编码存在明显的密码子偏倚现象。与普通牛相比较,甘南牦牛CYGB基因存在73处核苷酸变异位点,其中2处发生在外显子2区域,氨基酸序列未发生改变;71处发生在内含子区域。甘南牦牛CYGB拥有“three-over-three”类型的螺旋“三明治”折叠结构。系统发育树分析表明,甘南牦牛CYGB与黄牛、绵羊等物种之间存在较高的同源性。【结论】 成功克隆了甘南牦牛CYGB基因,为进一步研究牦牛CYGB的遗传特性和生理机制奠定了基础。
甘南牦牛;胞红蛋白;基因;生物信息学
胞红蛋白(Cytoglobin,CYGB)是德国学者Burmester等[1]于2002年研究发现的一种新型携氧球蛋白,它是继血红蛋白、肌红蛋白和脑红蛋白后发现的第4类球蛋白,广泛分布于脊椎动物的多种组织和器官[2]。近年来,因其在机体缺氧保护方面的重要作用,CYGB已逐渐成为国内外学者研究的热点[3-6]。目前,国内针对CYGB基因的研究尚处于起步阶段,主要涉及人和啮齿类动物[7-9],对生活在高寒低氧环境下的特有畜种牦牛CYGB的研究国内外均未见报道。本试验选取甘肃甘南牦牛类群为研究对象,对其CYGB基因进行克隆测序[10],从分子水平研究牦牛CYGB基因的序列结构及相关生物信息学特征,以期丰富牦牛携氧球蛋白基因组学研究的基础数据,为进一步探讨牦牛CYGB的遗传特性及相关生理机制提供理论依据。
1 材料与方法
1.1 材 料
1.1.1 血 样 从甘肃省草食动物生物技术重点实验室冻存的600份甘南牦牛抗凝血中随机抽取2份,用于基因组DNA提取。
1.1.2 主要试剂与仪器 LATaqDNA 聚合酶、pMD18-T载体和DNA快速纯化回收试剂盒均购自Takara公司;DNA Marker和大肠杆菌DH5α感受态细胞、质粒提取试剂盒购自北京天根公司;X-gal和IPTG购自Amersco公司;其他常规试剂均为进口或国产分析纯级产品。梯度PCR仪购自美国ABI公司。
1.2 基因组DNA的提取与检测
采用PBS-酚/氯仿法提取甘南牦牛基因组DNA,ddH2O溶解。用1%的琼脂糖凝胶电泳进行检测,-20 ℃冻存备用。
1.3 引物的设计与CYGB基因的分段扩增
参照GenBank中普通牛CYGB基因序列(GenBank登录号:AC_000176)设计4对引物,交由北京华大公司合成,引物序列和扩增片段长度如表1所示。PCR反应的总体积为25 μL,其中含:各引物10 μmol,LATaqDNA聚合酶 2.5 U,dNTP Mixture各0.8 mmol/L,LATaqBuffer Ⅱ(含5 mmol/L Mg2+),模板100 ng/μL。反应条件为94 ℃预变性4 min;94 ℃变性40 s,退火30 s (引物P1、P2、P3、P4退火温度分别为:59,60,56 和57 ℃),72 ℃延伸3 min,30个循环;最后72 ℃延伸10 min,4 ℃保存。PCR产物用1%的琼脂糖凝胶电泳检测。

表1 引物序列和扩增产物的长度Table 1 Primer sequences and sizes of amplified products
注:引物位置参考序列为普通牛CYGB基因(GenBank登录号:AC_000176)。
Note:Reference sequence of primer position isBostaurusCYGBgene (GenBank accession number:AC_000176).
1.4 CYGB基因的分段克隆
PCR扩增产物经1%琼脂糖凝胶电泳后,用胶回收试剂盒回收目的片段,与 pMD18-T克隆载体在4 ℃条件下过夜连接;连接产物转化至50 μL DH5α感受态细胞,转化后加900 μL LB培养液摇匀复苏1 h。将复苏后的菌液涂布于含0.1 mg/mL 氨苄青霉素(Amp)、0.5% IPTG及0.04 mg/mL X-gal的LB平板培养皿表面,37 ℃避光培养12 h ;挑取白色阳性单克隆菌落,用LB(Amp+)培养液37 ℃扩大培养,之后进行菌液PCR 鉴定。PCR反应体系为20 μL:菌液1 μL,上、下游引物各0.4 μL,LATaqDNA聚合酶12 μL,水6.2 μL;反应程序同1.3节。PCR产物用1%的琼脂糖凝胶电泳检测。将PCR检测呈阳性的菌液送北京六合华大基因科技股份有限公司进行测序。
1.5 生物信息学分析
运用Lasergene 7.1软件包中的EditSeq、SeqMan和GeneQuest对CYGB基因序列进行拼接、校对和分析;使用Bioedit和Protparam(http://web.expasy.org/protparam/)、PredictProtein (http://www.predictprotein.org/)、SWISS-MODEL(http://swissmodel.expasy.org/)等在线分析软件对CYGB的氨基酸序列进行分析,利用PyMol软件修饰并输出三维结构[11]。利用Lasergene 7.1软件包中的Megalign软件[12]及BLASTp程序(http://www.ncbi.nlm.nih.gov/BLAST/)对甘南牦牛和黄牛的CYGB基因及其编码氨基酸序列进行比对,并进行多序列的比对和同源性分析。
2 结果与分析
2.1 CYGB基因的分段扩增
对甘南牦牛基因组进行分段扩增,得到4条特异性较高的目的条带(图1),且大小与预期产物相符。克隆测序结果表明,P1-F/P1-R引物的扩增产物由3 127个核苷酸组成,P2-F/P2-R引物的扩增产物由1 523个核苷酸组成,P3-F/P3-R引物的扩增产物由2 052个核苷酸组成,P4-F/P4-R引物的扩增产物由2 438个核苷酸组成。对4段序列手动校对、拼接,使用NCBI数据库进行Blast搜索,结果表明扩增得到的序列是甘南牦牛CYGB基因,长度为8 484 bp,其中包含有4个外显子和3个内含子,外显子长度分别为143,232,164和34 bp,内含子长度分别为5 175,280和2 379 bp,该序列已提交至GenBank数据库(GenBank登录号:KF669897)。

图1 甘南牦牛CYGB基因的分段扩增M.DNA标准DL4500;1~4.分别为引物P1-F/P1-R、P2-F/P2-R、P3-F/P3-R、P4-F/P4-R扩增产物Fig.1 Separate amplification of CYGB gene of Gannan yakM.DNA Marker DL4500;1-4.Amplified products of P1-F/P1-R,P2-F/P2-R,P3-F/P3-R,P4-F/P4-R
2.2 甘南牦牛CYGB一级结构及其编码产物的理化性质分析
利用Lasergene 7.1软件包中的EditSeq和GeneGuest软件对甘南牦牛CYGB基因中核苷酸组成及其序列结构特征进行分析。结果(表2)发现,甘南牦牛CYGB基因序列(剪切掉5′和3′侧翼序列)A+T含量低于G+C含量,且其4个外显子区域(cDNA)的G+C含量远远高于A+T含量(G+C含量为63%,A+T含量为37%)。
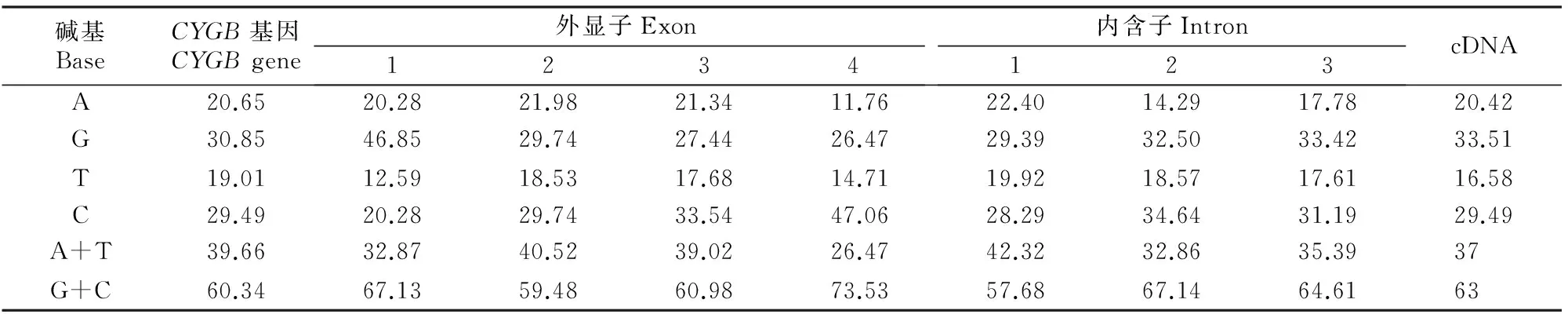
表2 甘南牦牛CYGB基因各核苷酸含量Table 2 Content of each nucleotide in Gannan yak %
对甘南牦牛CYGB基因编码区进行翻译,得到一条由190个氨基酸残基组成的多肽(表3)。由表3可知,CYGB多肽链包含所有的20种标准氨基酸,其中天冬氨酸、半胱氨酸、色氨酸数目最少,为3个;谷氨酸数目最多,为22个,占整个氨基酸组成的11.58%。在编码谷氨酸的过程中,密码子GAG使用了19次,GAA仅使用了3次,前者的使用频率是后者的6倍多;而在20个缬氨酸密码子中,GUG使用了16次,GAA仅使用1次,存在明显的密码子偏倚现象。由表3还可知,甘南牦牛CYGB氨基酸序列中含有29个碱性氨基酸(Lys、Arg、His)、25个酸性氨基酸(Asp、Glu)、71个非极性氨基酸(Ala、Ile、Leu、Phe、Trp、Val)、39个极性氨基酸(Asn、Cys、Gln、Ser、Thr、Tyr)。
通过Protparam和Bioedit软件对CYGB理化性质的分析,结果(表4)表明,甘南牦牛CYGB的相对分子质量为21 459.6,半衰期为30 h;不稳定系数为48.43(不稳定系数大于40即为不稳定性蛋白[13]),由此预测甘南牦牛CYGB具有不稳定性;总平均亲水性为-0.301,说明CYGB整体呈现亲水性,是可溶性蛋白[14]。推测该编码产物在pH 7.0环境下其电荷量为-1.143,理论等电点为6.618。

表3 甘南牦牛CYGB一级结构中各氨基酸和密码子的使用频率Table 3 Using frequency of amino acids and codons of the primary structure of CYGB gene of Gannan yak
注:密码子数据括号中的数字为密码子的使用次数
Note:The number in parentheses of codon is usage times.

表4 甘南牦牛CYGB的基本理化性质Table 4 Basic physicochemical properties of CYGB gene of Gannan yak
2.3 甘南牦牛与普通牛CYGB基因序列及氨基酸序列比对
将甘南牦牛与普通牛(GenBank登录号:AC_000176)的CYGB基因及其编码的氨基酸序列进行比对,结果显示,甘南牦牛CYGB基因存在73处核苷酸变异位点,其中有46处转换、20处颠换、4处插入和3处缺失;73处变异位点有71处发生在内含子区,有2处发生在外显子2区,其中外显子2区的2处突变都是同义突变,甘南牦牛CYGB氨基酸序列未发生改变;内含子区域在内含子1和内含子3中分别存在62处和9处核苷酸变异位点。
2.4 甘南牦牛CYGB二级结构和三级结构的预测与分析
利用PredictProtein软件对CYGB二级结构进行预测,结果显示α-螺旋(Helix)占64.21%,无规则卷曲(Coil)占35.79%,无β-折叠,根据α-螺旋比例大于45%,同时β-折叠比例小于5%的可定义为α-类蛋白质的原则[15]可知,甘南牦牛CYGB属于全α-类蛋白质。利用 SWISS-MODEL在线软件进行同源建模,得到CYGB三级结构模型。CYGB作为细胞内的单体球蛋白,属于脊椎动物6配位、低螺旋球蛋白中的一种,它拥有“three-over-three”类型的螺旋“三明治”折叠结构。
2.5 甘南牦牛与其他物种的CYGB同源性及系统进化分析
将甘南牦牛CYGB氨基酸序列与从GenBank中获得普通牛(AC_000176)、绵羊(NC_019468)、人(NC_000017)、猕猴(NC_007873)、野猪(NC_010454)、马(NC_009154)、家鼠(NC_005109)、斑马鱼(NC_007114)8个物种的CYGB氨基酸序列进行相似性分析,并利用MegAlign软件构建系统发育树。氨基酸序列相似性分析结果(表5)表明,甘南牦牛与黄牛CYGB同源性高达100%,与其他哺乳动物的同源性也较高(93.2%~98.4%),甘南牦牛与这些物种在系统发育树中距离较近,其亲缘关系较密切;与斑马鱼同源性较低,为48%,后者在系统发育树中处于另外的分支。CYGB系统发育树(图2)与生物进化的物种树基本一致,说明CYGB基因编码区在物种间比较保守,而且它们都有非常相近的生物学功能。

表5 各种动物CYGB同源性比较Table 5 Identity and divergence of CYGB genes of different animals

图2 基于CYGB基因编码区构建的各物种的系统发育树Fig.2 Phylogenetic tree of different species based on CYGB gene coding region
3 讨 论
目前,国内外针对牦牛CYGB的研究尚属空白。本试验利用PCR克隆及核酸测序技术获得甘南牦牛CYGB基因的核苷酸序列,并采用生物信息学方法分析了甘南牦牛CYGB的结构与特性[16]。结果表明,在甘南牦牛CYGB多肽链的组成中,不同的氨基酸数目相差较大,而相同的氨基酸倾向于使用一种或几种特定的同义密码子,说明甘南牦牛CYGB基因编码氨基酸的过程中存在明显的密码子偏倚现象[17]。饶友生等[18]研究认为,密码子选择由可供使用的tRNA的丰度所决定。通过对牦牛密码子使用偏性的研究,利用其偏性特点,可以筛选出牦牛CYGB基因的最优密码子,有针对性地设计相应的基因表达载体,提高CYGB基因的表达量。
高原低氧适应性研究一直备受科学家们的关注,但其适应机制十分复杂,目前尚未能找到与高原低氧适应直接相关的基因。对CYGB基因结构的研究发现,其与缺氧关系密切,是缺氧可调节型表达指示器[19],这为高原低氧适应性的研究提供了新的思路。CYGB在中枢神经系统内分布广泛,其中海马、丘脑和下丘脑表达量较高,这3个区域能迅即感应缺氧应激,表明CYGB可能在缺氧损伤中起到重要作用[20-21]。这些研究为探讨CYGB与高原低氧适应性之间的关联提供了有价值的理论依据。本试验通过对甘南牦牛与黄牛CYGB基因序列进行比对,发现牦牛CYGB基因内含子存在4个碱基的插入和3个碱基的缺失,这可能与高原低氧环境下CYGB的表达调控具有一定联系。
贾浩等[22-23]研究认为,蛋白质半衰期与其稳定性之间存在密切联系,一般来说,半衰期长则蛋白质稳定性高。本研究结果显示,CYGB具有较长的半衰期,但却属于不稳定蛋白,这一特性与其生理功能的发挥是否存在某种联系有待更进一步的研究和探讨。甘南牦牛CYGB二级结构和三级结构的预测与分析表明,牦牛CYGB属于全α-类蛋白质,呈“three-over-three”类型的螺旋三明治折叠结构,这与前人研究结果[24]基本一致。
利用Megalign软件对CYGB编码产物进行同源性比对,发现甘南牦牛与牛、绵羊等物种在系统发育树中距离最近,其亲缘关系最为密切。CYGB基因编码产物的系统发育树与生物进化的物种树基本一致,这一结果与动物分类学的观点相一致,也说明CYGB基因编码区在这些物种间比较保守。
4 结 论
本研究克隆出8 484 bp的甘南牦牛CYGB基因,其中包含有4个外显子和3个内含子,外显子长度分别为143,232,164和34 bp,内含子长度分别为5 175,280和2 379 bp。甘南牦牛外显子存在2处核苷酸变异位点,二者都是同义突变,未导致氨基酸序列发生改变;内含子区域存在71处核苷酸变异位点。甘南牦牛系统发育树与生物进化的物种树基本一致,说明CYGB基因编码区在物种间比较保守。
[1] Burmester T,Ebner B,Weich B,et al.Cytoglobin:A novel globin type ubiquitously expressed invertebrate tissues [J].Mol Biol Evol,2002,19:416-421.
[2] Trent J T Ⅲ,Hargrove M S.A ubiquitously expressed human hexacoordinate hemoglobin [J].J Biol Chem,2002,277:19538-19545.
[3] Hodges N J,Innocent N,Dhanda S,et al.Cellular protection from oxidative DNA damage by over-expression of the novel globin cytoglobininvitro[J].Mutagenesis,2008,23(4):293.
[4] Fordel E,Thijs L,Martinet W,et al.Anoxia or oxygen and glucose deprivation in SH-SY5Y cells:A step closer to the unraveling of neuroglobin and cytoglobin functions [J].Gene,2007,398(1/2):114.
[5] Fordel E,Thijs L,Martinet W,et al.Neuroglobin and cytoglobin overexpression protects human SH-SY5Y neuroblastoma cells against oxidative stress-induced cell death [J].Neurosci Lett,2006,410(2):146.
[6] Ostojic J,Grozdanic S D,SYED N A,et al.Patterns of distribution of oxygen-binding globins,neuroglobin and cytoglobin in human retina [J].Arch Ophthalmol,2008,126(11):1530.
[7] Kawada N,Kristensen D B,Asahina K,et al.Characterization of a stellate cell activation-associated protein(STAP) with pemxidase activity found in rat hepatic stellate cells [J].J Biol Chem,2001,276(84):25318-25323.
[8] Shigematsu A,Adachi Y,Matsubara J,et al.Analyses of expression of cytoglohin by immunohistochemical studies in human tissues [J].Hemoglobin,2008,32(3):287-296.
[9] 尹 宏,白振忠,靳国恩,等.藏羚羊脑组织胞红蛋白基因的表达 [J].青海医学院学报,2012,33(1):47-50.
Yin H,Bai Z Z,Jin G E,et al.Expression of cytoglobin genes in brain tibetan antelopes [J].Journal of Qinghai Medical College,2012,33(1):47-50.(in Chinese)
[10] 石宁宁,杜晓华,刘 霞,等.甘南牦牛NGB基因克隆及序列分析 [J].西北农林科技大学学报:自然科学版,2013,41(4):1-7.
Shi N N,Du X H,Liu X,et al.Cloning and sequence analysis of NGB gene in Gannan yak [J].Journal of Northwest A&F University:Natural Science Edition,2013,41(4):1-7.(in Chinese)
[11] 陶士珩.生物信息学 [M].北京:科学出版社,2007.
Tao S H.Bioinformatics [M].Beijing:Science Press,2007.(in Chinese)
[12] 刘 菲,赵婷婷,程安春,等.天府肉鹅IFN-α基因的克隆及生物信息学分析 [J].中国兽医科学,2010,40(12):1271-1275.
Liu F,Zhao T T,Cheng A C,et al.Cloning and bioinformatics analysis of IFN-α gene of Tianfu meat-type goose [J].Chinese Veterinary Science,2010,40(12):1271-1275.(in Chinese)
[13] Guruprasad K,Reddy B,Pandit M.Correlation between stability of a protein and its dipeptide composition:A novel approach for predictinginvivostability of a protein from its primary sequence [J].Protein Eng,1990,4:155-161.
[14] Kyte J,Doolittle R F.A simple method for displaying the hydropathic character of a protein [J].J Mol Biol,1982,157:105-132.
[15] Rost B,Sander C.Combining evolutionary information and neural networks to predict protein secondary structure [J].Protein Sci,1994,19(1):105-132.
[16] 李盛杰,杜晓华,罗玉柱,等.天祝白牦牛NGB基因的克隆及生物信息学分析 [J].畜牧兽医学报,2013,44(3):395-398.
Li S J,Du X H,Luo Y Z,et al.Cloning and bioinformatics analysis ofNGBgene of Tianzhu white yak [J].Acta Veterinaria Et Zootechnica Sinica,2013,44(3):395-398.(in Chinese)
[17] 刘庆坡,冯 英,董 辉.20个物种同义密码子偏性的比较分析 [J].西北农林科技大学学报:自然科学版,2004,32(7):67-71.
Liu Q P,Feng Y,Dong H.Comparative studies on synonymous codon usage bias in twenty species [J].Journal of Northwest Sci-Tech University of Agriculture and Forestry:Natural Science Edition,2004,32(7):67-71.(in Chinese)
[18] 饶友生,梁菲菲,王樟凤,等.鸡Z染色体基因表达的密码子偏性 [J].华南农业大学学报,2010,31(1):70-74.
Rao Y S,Liang F F,Wang Z F,et al.Study on the codon bias of genes expression in GGAZ [J].Journal of South China Agricultural University,2010,31(1):70-74.(in Chinese)
[19] 郑淮武,史雪川.胞红蛋白在缺氧下的表达 [J].现代生物医学进展,2008,8(3):580-582.
Zheng H W,Shi X C.Expression of cytoglobin under hypoxia [J].Progress in Modern Biomedicine,2008,8(3):580-582.(in Chinese)[20] Geuens E,Brouns I,Flamez D,et al.A globin in the nucleus! [J].J Biol Chem,2003,278(33):30417-30420.
[21] Mammen P P,Shelton J M,Ye Q,et al.Cytoglobin is a stress-responsive hemoprotein expressed in the developing and adult brain [J].J Histochem Cytochem,2006,54(3):1349-1361.
[22] 贾 浩.基于序列特征的胞内蛋白稳定性计算分析 [D].南京:南京航空航天大学,2011.
Jia H.Calculation and analysis of intracellular protein stability based on sequence features [D].Nanjing:Nanjing University of Aeronautics and Astronautics,2011.(in Chinese)
[23] 贾 浩,张小白,宋晓峰.人类胞内蛋白半衰期与其亚细胞定位的相关性研究 [J].计算机与应用化学,2011,28(4):411-414.
Jia H,Zhang X B,Song X F.Relationship between intracellular protein half-life and subcellular localization in human cells [J].Computers and Applied Chemistry,2011,28(4):411-414.(in Chinese)
[24] Sugimoto H,Makino M,Sawai H,et al.Structural basis of human cytoglobin for ligand binding [J].J Mol Biol,2004,339(4):873.
Cloning and bioinformatics analysis ofCYGBgene of Gannan yak
LOU Yan-yue1a,2,DU Xiao-hua1b,2,LUO Yu-zhu2,LIU Xia1a,2,ZHANG Lei3
(1 aCollegeofLifeScienceandTechnology,bCollegeofVeterinaryMedicine,GansuAgriculturalUniversity,Lanzhou,Gansu730070,China;2GansuKeyLaboratoryofHerbivorousAnimalBiotechnology,Lanzhou,Gansu730070,China;3DepartmentofAnimalScience,HenanVocationalCollegeofAgriculture,Zhengzhou,Henan451450,China)
【Objective】 This study aimed to elucidate the structure and physiological function ofCYGBgene. 【Method】 TheCYGBgene of Gannan yak was amplified through PCR using extracted genomic DNA sequence from blood samples as template.Then the gene was sequenced and analyzed by bioinformatics tools.【Result】 The length of cloned Gannan yakCYGBgene was 8 484 bp,including four exons with lengths of 143,232,164 and 34 bp,and three introns with lengths of 5 175,280 and 2 379 bp,respectively.A total of 190 amino acids were encoded and the usage of synonymous codons was biased.Compared with taurine cattle (Bostaurus),there were 73 nucleotide mutations inCYGBgene of Gannan yak,among which 2 occurred in exon region and 71 occurred in intron region.There were no changes in amino acids.Gannan yakCYGBgene had a spiral sandwich like folded structure with the “three-over-three” type.Phylogenetic analysis showed that Gannan yakCYGBgene had a high homology withBostaurus,Ovisariesand other species.【Conclusion】 Gannan yakCYGBgene was successfully cloned and could be used for further study on genetic characteristics and physiological mechanism.
Gannan yak;cytoglobin;gene;bioinformatics
2013-11-23
甘肃省自然科学基金项目(1107RJZA148);甘肃省教育厅研究生导师项目(1102-04);甘肃省高等学校基本科研业务费资助项目(2013);甘肃农业大学科技创新基金项目(GAU-CX1037,GAU-CX1038)
娄延岳(1986-),女,河南南阳人,在读硕士,主要从事动物遗传与分子生物学研究。E-mail:1023498596@qq.com
刘 霞(1975-),女,甘肃兰州人,副教授,博士,硕士生导师,主要从事动物遗传与分子生物学研究。 E-mail:liuxia@gsau.edu.cn
时间:2015-03-12 14:17
10.13207/j.cnki.jnwafu.2015.04.003
S823.85
A
1671-9387(2015)04-0001-06
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20150312.1417.003.html
