石羊河下游7种沙生灌木的构型比较
2015-07-05郭春秀袁宏波徐先英刘虎俊王多泽李学敏刘开琳
郭春秀,袁宏波,徐先英,刘虎俊,王多泽,李学敏,刘开琳
(甘肃省治沙研究所&甘肃省荒漠化与风沙灾害防治重点实验室-省部共建国家重点实验室培育基地,兰州730070)
植物构型用来表述其不同构件在空间的排列方式,是植物体不同组分的空间位置及组合,植物构型狭义上就是枝系结构和分枝格局,最终决定了枝系的复杂化程度[1]。目前,植物构型研究主要集中于农作物和热带乔灌木,分析其构型与生物量或产量关系,揭示了不同构件之间的相互作用和空间生物量分配[1-4]。植物构型与其生态环境适应性研究较多,认为植物在适应不同生态环境条件下会产生趋同或趋异适应特征,特别是表现在外部形态特征[1]。在荒漠植物的构型研究中,进行了构型分类[5-6],提出了灌木空间结构是植物发育生长和适应的结果,认为灌木构型对其防风固沙能力有非常重要的影响[7-8]。研究沙生灌木构型对其个体构型与其防风固沙机理及效益,特别是应用其配置与建设防风固沙林具有重要意义。但是干旱区沙生灌木构型与其防风固少效能研究相对较少,仅观测分析了梭梭(Haloxylon ammodendron)、油蒿(Artemisia ordosica)和沙蒿(A.arenaria)的枝条空间分布及其防风功能[9-12]。为了比较相同环境下不同灌木的构型特征,揭示其防风固沙机理,也为研究仿真固沙灌木以及仿真模拟研究提供基础数据。在石羊河下游的民勤选择了7种常见灌木,观测分析其构型,为说明其生物学特征和仿制固沙灌木提供参考。
1 研究区概况
试验地选在巴丹吉林沙漠和腾格里沙漠交界处的石羊河下游的民勤(38°38′N,103°05′E),海拔1 378m,属温带干旱气候,具典型大陆性荒漠气候特征,日照强烈,昼夜温差大,干旱少雨,强烈辐射,蒸发量大,风大沙多,干燥度4.5以上,地下水位18 m 左右。年日照时数2 420h,初霜期10月上旬,终霜期4月下旬,无霜期171d,年平均降水量116.2 mm,年平均蒸发量2 643.9mm,年平均有风的记录占99.23%,≥5.0 m/s起沙风占全年有风日数的13.08%,4月份和5月份为大风和沙尘暴多发月。
2 材料和方法
在沙旱生灌木有叶期(5月),调查测定民勤的7种沙旱生灌木构型参数,被观测的7种灌木为:梭梭(Haloxylon ammodendron)、油蒿(Artemisia ordosica)、沙蒿(A.arenaria)、花棒(Hedisarum scoparium)、沙 拐 枣(Calligonum mongolicum)、白 刺(Nitraria tangutorum)、红 砂(Reaumuria kaschgarica)。因这7种灌木只有白刺的叶相对较大,其余6种的叶特化或完全退化,所以主要观测和分析指标包括高度、分枝角度、枝长、分枝率、分枝分维数、侧影面积和迎风面枝条密度。观测植株数5~7株,被观测植株均为散点分布或单株,95%植株高度为50~60cm,生长良好,株型相对整齐,受周围树木影响较小。植株的枝序按Strahler法[3-4]确定,即由外及内确定枝序,外层的第1小枝为第1级,2个第1级相遇为第2 级,2 个第2 级相遇后则为第3级,依此类推,如有不同枝级相遇,相遇后则取较高的作为枝级,计算各级枝数的相互比率。以10cm为分层高度,对所观测灌木的梯级高度宽度进行测量,并计算累积面积,计算侧影面积和迎风面枝条密度[6-8,13]。冠幅计算用椭圆面积公式,其余相关因 子的计算公式如下:

式中,Nt=∑Ni,为所有枝的总数;Ns为最高级枝条数;N1为第一级的枝条数;Ni和Ni+1是第i和第i+1级枝条总数。

式中,C为比例数(常数),LB 为枝长,D 分形维数,SB 与LB 在双对数坐标下存在一种线性关系,D 为该直线的斜率,lnSB=lnC+DlnLB,通过回归方程分析就可以求出分形维数D 值。

式中,Hi为植株第i段高度,Wi为第i段迎风面宽度。

式中,PS为面积S 内的枝条数目,S 为迎风面面积。
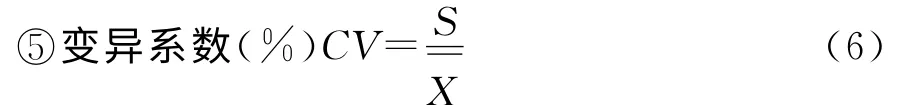
式中,CV(%)变异系数是标准差与其平均数的比,比较两组数据离散程度大小,是反映数据离散程度的绝对值。S 为统计数据的标准差,为统计数据的平均数。
文中所有数据统计、指标具体数值计算均在Excel 2007下完成,用SPPS 13.0进行数据差异性比较。
3 结果与分析
3.1 7种灌木的枝条分枝角度
所观测的7 种灌木分枝角度有10.2%的小于40°,有21.4%的分枝角为60°~70°。花棒和白刺二级枝角度的平均值最小,其余6种灌木的分枝角度约呈自冠层内向外的分枝角度逐渐变小趋势(表1)。油蒿各级枝的分枝角变异最大,其各级枝的分枝角变异系数达2.4%,其余灌木的均小于2%。梭梭的一级枝分枝角度平均较四级枝大7.8°。红砂和油蒿一级枝分枝角度平均较四级枝相差分别为15.6°和和24.2°,差异显著。沙蒿的一级枝分枝角度与四级枝的相差11.5°。沙拐枣三级枝分枝角度最大,一级枝和四级枝分枝角度平均值相等。白刺的分枝角度除一级枝分枝和二级分枝的小于油蒿外,三级分枝角度和四级分枝角度均大于其他灌木,是分枝角度较大的灌木。花棒的各级分枝角相对较小,是分枝角度较小的灌木。
3.2 7种灌木枝条的长度特征
所观测的7种灌木中只有油蒿的冠层外枝长较大,其余则均表现为越向冠层外,枝长度越短的趋势(表2)。其中沙拐枣的四级枝长与一级枝长相差3倍,红砂的四级枝长与第一级相差最小,为4.3cm,平均枝长也最短,只有6.9cm。沙蒿和梭梭各级枝长间的差异最小,沙拐枣的各级枝长变化较大,枝长变化是7种灌木的枝长中最大的。
从各级分枝长度变化以及所占比率分析,有叶的白刺有86.9%枝条长度为5~30cm,而花棒的枝条比较长,也比较分散,主要分布区间为30~50cm枝占37.8%。无叶的沙拐枣和梭梭的枝条长度中5~30cm 者达77.6%,棒状叶的红砂的枝长5~15 cm 有63.6%,沙蒿的枝长在15~30cm 范围之间所占比例较大,约为52.3%(图1)。
3.3 7种灌木枝条的分枝率
油蒿和沙蒿的总体分枝率相差近3倍,梭梭和沙拐枣的总体分枝率相差2.4倍,红砂的整体分枝率约是白刺的1/3。7种灌木的分枝率由树冠层外围向内呈逐步增加趋势。除梭梭外,其他6种灌木的总体分枝率都小于1,说明6种灌木的总体分枝是灌木树冠中心的分枝密度小于最外层。7种灌木中,差异性最大表现在三级和四级枝的数量,变异系数最大(0.93%),其次为二级和三级枝的数量,一级和二级枝的变异最小(表3)。
油蒿的逐步分枝率要比沙蒿小。沙蒿和白刺的二级分枝数量相对其他各级枝数最少,形成树冠层内疏,外层较紧密的树冠。梭梭的各级分枝数比较均匀,是7种灌木中各级分枝数变化最小的灌木(变异系数为0.12)。梭梭的分枝数是由树冠内向外增加,是一种较均匀的树冠。沙拐枣的二级枝数量相对来说较多,各级枝的分枝数量变异较大。白刺的一级和二级枝数量几乎相等,而三级枝的数量约为二级枝数量的2倍多,三级枝数量又是四级枝数量的72%,白刺是一种内密外疏型构型。花棒与白刺为有叶灌木,但花棒的分枝率为白刺的62%。红砂的总体分枝率是7种被观测灌木中最小的,是外密内疏的冠层结构。红砂总体分枝率约是白刺的1/3,红砂的枝数是白刺和花棒25%左右。

表1 7种灌木的不同分级枝条的平均分枝角度比较表Table 1 The average crotch angles of 7shrubs on different grade branches
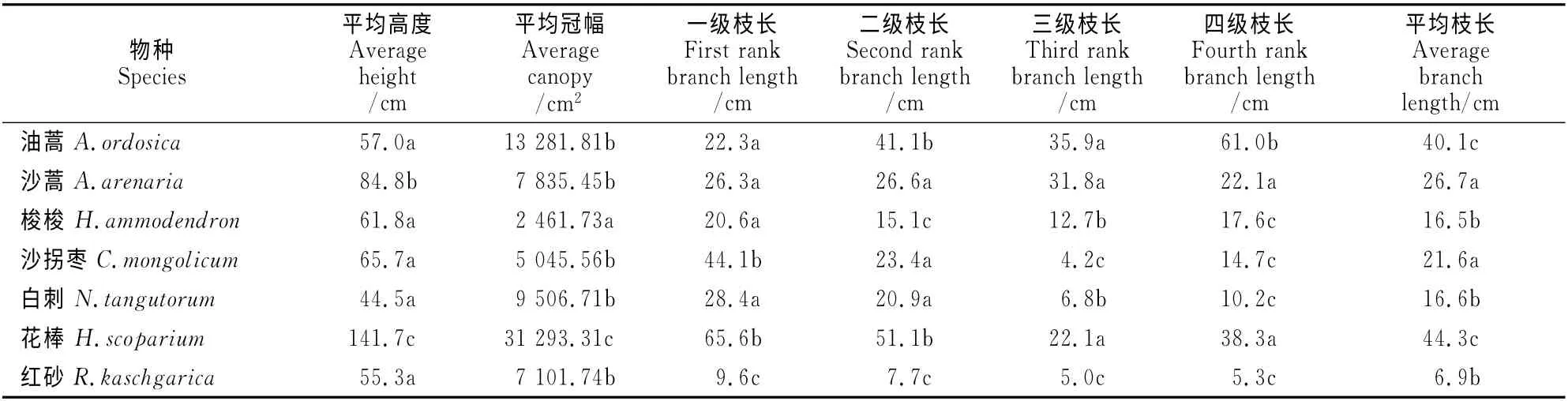
表2 7种沙旱生灌木不同分级枝的平均长度Table 2 The average branch length of 7shrubs on different grade branching
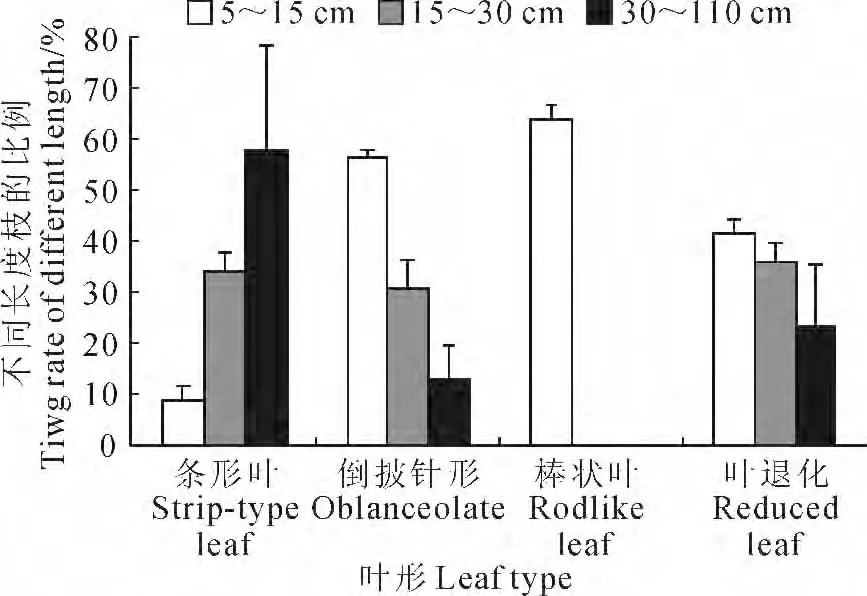
图1 叶形与不同长度枝条所占百分比关系图Fig.1 The relation between branch length ratio and shape of leaf
3.4 6种灌木枝条的分枝分维数
沙拐枣的枝长和枝数之间的相关性极显著(P<0.01),其拟合方程灵敏度也较高,说明被调查沙拐枣的枝数与其枝长间相互影响较强烈。红砂的分枝分维数最大,其枝系结构最复杂,其次为梭梭和白刺,沙拐枣的分枝分维数只是大于花棒的0.34(表4)。花棒的枝系结构相对较简单。梭梭和沙拐枣的叶都退化,但沙拐枣的分枝分维数只有梭梭的1/2,梭梭枝系结构相对来说较沙拐枣复杂。白刺、花棒和沙蒿三者中,花棒的分枝分形维数最小,白刺的枝系结构相对较为复杂。红砂的枝数与枝长的分形维数相关性不显著,其余灌木的枝数与枝长的分形维数的相关性均显著,而沙拐枣的差异性极显著,沙蒿和白刺次之。
3.5 灌木的侧影面积与迎风面几何形状
白刺和红砂的侧影面积在高度10cm 内最大,白刺和红砂树冠近似成坛状形态。花棒、梭梭和沙蒿的侧影面积最宽处则为高度20~40cm 范围,略成梭形(图2)。花棒侧影面积随高度的变化幅度最小,在40cm范围侧影面积均是10、20和30cm范围侧影面积的1.1倍,达到高度50~70cm 则侧影面积逐渐缩小,70~90cm 的侧影面积约为20~40 cm 的1/2。梭梭侧影面积变化趋势与花棒相似,20~40cm 高度范围的侧影面积约为10cm 范围侧影面积的1.2倍,50~70cm 高度范围的侧影面积约是10cm 范围侧影面积的1/3。梭梭和沙蒿侧影面积最宽处在30cm,花棒在高约10cm 处。沙蒿侧影面积相对变化较小,最大值在80cm2左右。沙拐枣的侧影面积则随高度变化幅度与其他几种灌木相反,在中 间30cm 处 较20cm 和40cm 处 反 而 宽 度较大,几乎成扫帚形态。
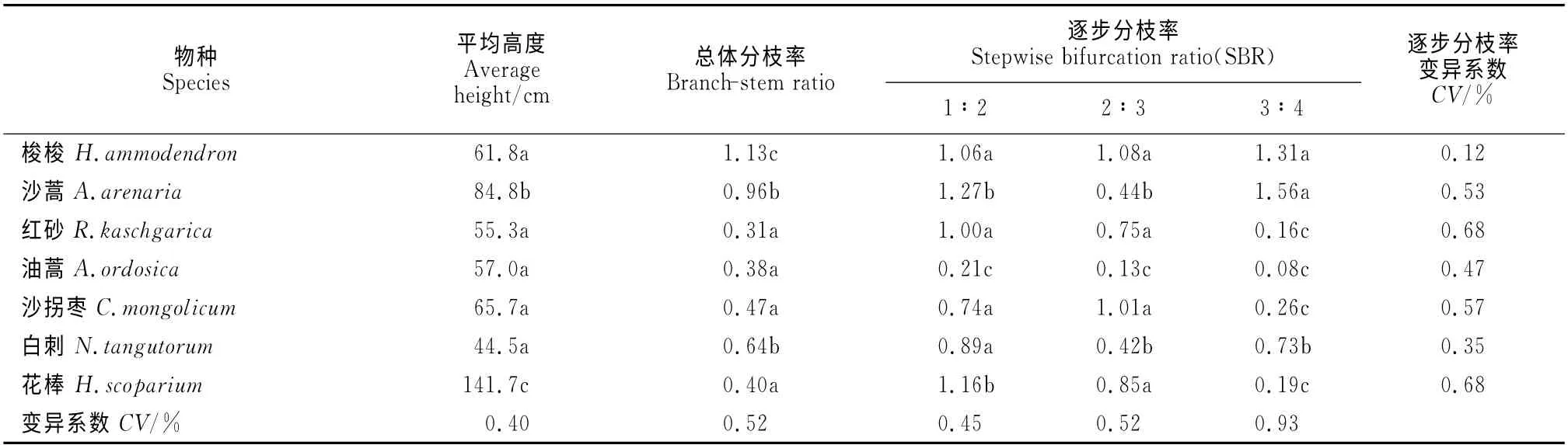
表3 7种灌木的平均总体分枝率和逐步分枝率Table 3 The branch-stem ratio and the stepwise bifurcation ratio of seven shrubs

表4 6种灌木的枝长和枝数的对数线性关系Table 4 Logarithm relation between length and number of branches of six shrubs
3.6 迎风面枝条密度
在所有观测的树高相近的梭梭和沙拐枣,平均枝条密度相差5倍多,红砂枝条总数是白刺的2倍左右。除红砂之外,所观测灌木的枝条最密集的高度都在树高0.5倍处左右。红砂、沙蒿、白刺、梭梭、沙拐枣、花棒的枝条密度最大值依次出现在树高的0.54~0.72、0.47~0.59、0.44~0.89、0.32~0.49、0.31~0.45、0.28~0.42倍处(图3)。只有花棒枝条密度最小的高度在0~20cm 高度,其余均在0~10cm 范围内。沙蒿的40~50cm 高度范围内的枝条数占总数的45.6%,红砂的30~40cm 高度范围内的枝条数占总数的67.9%,沙拐枣的20~30cm高度范围内的枝条数占总数的57.1%,白刺的20~30cm 高度范围内的枝条数占总数的55.1%,花棒的20~30cm 高度范围内的枝条数占总数的31.6%,梭梭的20~30cm 高度范围内的枝条数占总数的46.2%。从不同高度枝条密度的变异系数分析均匀度,沙拐枣(0.70)>梭梭(0.66)>花棒(0.47)>红砂(0.47)>沙蒿(0.30)>白刺(0.20),白刺的均匀度最好。
4 讨 论
在不同环境生长的同种植物构型有所差异[1,3]。张德魁等[9]、詹 科 杰 等[10]和 李 建 刚 等[11]研究了油蒿、沙蒿、梭梭、柠条、花棒和沙拐枣构件组成特征,并对其固沙机制和抗风蚀能力进行了分析。他们所描述的上述灌木分枝角度与本研究观测结果略有不同。这可能与其生长区域相关,张德魁等和詹科杰所观测沙蒿和油蒿都分布半干旱区,生长茂盛。李建刚所测梭梭和花棒具有较强的分枝能力,其分枝角度一般都在70°~85°,这主要是李建刚所选择的梭梭、花棒和沙拐枣均为120~160cm,观测植株较高。本研究所观测梭梭、花棒的高度没有超过100cm,树龄较小,枝序较少,枝长也较短。这也进一步说明,不同生命期,植物构型略有不同。从枝系格局分析,叶特化灌木的各级枝分枝角变化较小,有叶灌木的冠层外一级枝条的分枝角度大于叶特化灌木。油蒿的逐步分枝率比沙蒿的低,这是由于油蒿与沙蒿这两种半灌木一般具有3~4级分枝,进行光合作用的大部分叶片着生在新枝(一级和二级枝)上,二级枝上着生极少数的叶片。白刺沙埋后生长更加旺盛,三级和四级枝数量较多有利于植物生长和生存。最高级(四级)的枝数量约为其余各级枝数量的总和。花棒外层(一级和二级)枝数量相对较少,是一种内密外疏型构型,这可能与其枝上生长叶进行光合作用相关。红砂是一类过渡类型,这正好与其叶型相一致。由于红砂的当年生枝端冬季枯死,且有二次发枝现象,所以最外层枝相对还是比较多。灌木的构型主要决定于其生物学特性[4-5],当然也受其树龄和生境变化影响。为了说明观测灌木防风固沙效能,本项研究没有根据植物年龄,而是以植株的高度为标准,所观测灌木不同植株尽可能选择高度相近,以缩小系统误差。这也是本观测分析结论与其他研究结果存在差异的主要原因。本研究分析的7种灌木种生活在相似的环境下,其构型是植物发育生长和适应沙漠环境结果。生长在风沙环境灌木形成了适应环境的构型,防风固沙效能各异,为仿真固沙灌木选择提供了参考。

图3 灌木枝密度随高度变化关系图Fig.3 The change of density ratio of shrub with the height
灌木防风固沙效能与其树冠构型与枝系格局关系密切,为了说明两者关系,Lancaster等[12]提出了侧影面积和粗糙元密集度等概念。研究表明,植物防风效应主要来自于迎风方向的侧影面积,树冠形态决定其侧影面积,从而影响其防风阻沙能力[7,14]。本研究分析结论最大枝条密度高度最低是白刺,这与其容易形成沙堆相一致[15]。不同形态的植物防风阻沙强弱不同,半球体树冠的植物比锥体和梭形树冠的植物阻沙能力更大,可截留更多的沙物质[15-16],这也为仿真固沙灌木制作提供了参考。
7种灌木的分枝角度是自冠层内向外逐渐变大,枝序级都为四级,只有油蒿的冠层外枝较短。除梭梭外,6种灌木的枝分布都是外密内疏。红砂、白刺、油蒿和花棒为密生型灌木,梭梭、沙拐枣和沙蒿为疏生型灌木。侧影面积随高度变化为:白刺>梭梭>红砂>沙拐枣>沙蒿>花棒。白刺和红砂的树冠近似坛状形态,花棒、梭梭和沙蒿略成梭形,沙拐枣成扫帚形态。
[1] CHEN B(陈 波),SONG Y CH(宋永昌).Woody plant architecture and its research in plant ecology[J].Chinese Journal of Ecology(生态学杂志),2002,21(3):52-56(in Chinese).
[2] CHANG J(常 杰),CHEN G(陈 刚),GE Y(葛 滢).A new method of quantitative study on plant structure and morphology-fractal simulation[J].Chinese Bulletin of Botany(植物学通报),1996,13(2):57-62(in Chinese).
[3] 常 杰.植物结构的分形特征及模拟[M].杭州:杭州大学出版社,1995:10-55.
[4] SPRUGEL D G,HINCKLEY T M,SCHAAP W.The theory and practice of branch autonomy practice of branch autonomy[J].Annual Review of Ecology and Systematics,1991,22:309-334.
[5] 何明珠.荒漠植物枝系构型分类研究[C]//中国科学院寒区旱区生态与工程研究所沙坡头沙漠试验研究站.中国沙漠研究与治理50年.北京:海洋出版社,2005:285-292.
[6] HE M ZH(何明珠),WANG Y H(王摇辉),ZHANG J G(张 景 光).Classification of the branching architectures of the desert plants in Minqin County[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2005,25(9):1 827-1 832(in Chinese).
[7] TANG Y(唐 艳),LIU L Y(刘连友),QU ZH Q(屈志强),et al.Research review of capacity of plant for trapping blown sand[J].Journal of Desert Research(中国沙漠),2011,31(1):44-48(in Chinese).
[8] SHU J M(苏金梅),LI G T(李钢铁),QIN F C(秦富仓),et al.A prediction study on aboveground biomass of artificial Haloxylon ammodendronforest[J].Inner Mongolia Forestry Science and Technology(内蒙古林业科技),1997,(3):9-11(in Chinese).
[9] ZHANG D K(张德魁),WANG J H(王继和),MA Q L(马全林),et al.Study on branch module characteristics of Artemisia ordosica and A.arenaria[J].Grassland and Turf(草原与草坪),2009,(1):43-46(in Chinese).
[10] ZHAN K J(詹科杰),WANG J H(王继和),MA Q L(马全林),et al.The space construction of of Artemisia arenariaand A.ordosicaand the research of dune-fixating[J].Journal of Gansu Forestry Science and Technology(甘肃林业科技),2005,30(5):1-4(in Chinese).
[11] LI J G(李健刚),WANG J H(王继和),JIANG ZH R(蒋志荣),et al.Study on the tree spatial structure and the wind break function on psammophyte in Minqin County[J].Research of Soil and Water(水土保持研究),2008,15(3):121-124(in Chinese).
[12] LANCASTER N,BASS A.Influence of vegetation cover on sand transport by wind:field studies at Owens Lake.Califorma[J].Earth Surface Processes and Landforms,1986,11:505-514(in Chinese).
[13] HUANG F X(黄富祥),WANG M X(王明星),WANG Y S(王跃思),et al.Recent progress on the research of vegetation protection in
soil erosion by wind[J].Chinese Journal of Plant Ecology(植物生态学报).2002,26(5):627-633(in Chinese).
[14] 董治宝,苏志珠,钱广强,等.库姆塔格沙漠风沙地貌[M].北京:科学出版社,2011:272-287.
[15] 董治宝.中国风沙物理50年[C]//中国科学院寒区旱区生态与工程研究所沙坡头沙漠试验研究站.中国沙漠研究与治理50年.北京:海洋出版社,2005:87-105.
[16] WU L(乌 拉),ZHANG G Q(张国庆),XIN ZH M(辛智鸣).Mechanism and effect of wind break and sand blocking for single natural shrub[J].Inner Mongolia Forestry Science and Technology(内蒙古林业科技),2008,34(2):36-39(in Chinese).
