温度、光照、盐分和干旱胁迫对华北驼绒藜种子萌发的影响
2015-07-01何青山魏玲玲刘国方杨慧玲
何青山, 魏玲玲, 张 旭, 刘国方, 杨慧玲
(1.河南农业大学生命科学学院,河南 郑州 450002;2.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093)
温度、光照、盐分和干旱胁迫对华北驼绒藜种子萌发的影响
何青山1,2, 魏玲玲1, 张 旭1, 刘国方2, 杨慧玲1,2
(1.河南农业大学生命科学学院,河南 郑州 450002;2.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093)
通过测定温度、光照、盐分和干旱胁迫(NaCl和PEG)对华北驼绒藜(Ceratoidesarborescens)种子萌发的影响,探究该物种对荒漠生境的适应对策。结果表明,华北驼绒藜种皮具有透水性,吸水2 h后开始萌发,低温(5/15和5 ℃)条件下种子平均萌发时间(MGT)延长,萌发速率(GR)及发芽指数(GI)降低,但在不同光照和温度处理下,所有梯度均可达到较高最终萌发率(95%~100%),因此,该物种种子无休眠;种子初始萌发率随溶液渗透势降低而下降,渗透势高于-1.4 MPa的等渗溶液对种子初始萌发无显著影响,但是发芽指数(GI)显著降低;而在渗透势-2 MPa,种子仍保持10%~14%的萌发率,高渗透势溶液(0 ~-2.0 MPa)处理下的种子具有较高的萌发恢复率,而-2.7 MPa NaCl处理下的种子死亡率高出与其等渗的PEG溶液处理的51%,高盐分胁迫会导致种子失去活力;华北驼绒藜种子具有快速的萌发特性及耐旱特性,有利于该物种在干旱荒漠环境的适应和种群维持。
华北驼绒藜;种子萌发;等渗溶液;温度;光照
种子萌发是群落物种更新的关键和基础,既影响着群落的建成、结构及动态,又直接影响着群落中物种的更新、分布及多度[1]。生长在荒漠或半荒漠地区的物种往往通过特有的传播方式及萌发策略适应恶劣的环境变化,从而保证种群在不利环境中的更新和延续[2,3]。种子萌发不仅受到种子内部因素的制约,而且受到外部环境因子的影响[4]。在环境恶劣地区(荒漠、沙化、盐碱地等),通常由于降水量少、蒸发量及风沙大等因素,盐分和水分成为影响种子萌发和幼苗生长的主要因素。因而,探究物种对这些地区环境的适应机制成为制定植被保护或恢复对策研究的首要问题[1]。华北驼绒藜(Ceratoidesarborescens)为藜科驼绒藜属多年生强旱生灌木,全株高1~2 m,果实管外中上部具4束长毛,下部具短毛;果实狭倒卵形,被毛;花果期为7~9月[5]。常生长于固定沙丘、沙地、荒地或山坡上,为中国特有植物。该物种营养丰富、适口性好、饲用价值高,是荒漠、半荒漠地区重要的牧草,并且具有良好的防风固沙作用。华北驼绒藜物种在优质牧草经济使用价值及荒漠化生态防护方面都具有较强的应用价值,但是长期的过度放牧和人为自然破坏等因素已使得华北驼绒藜野生种群日渐衰退[6]。目前对华北驼绒藜开展的研究工作相对较少,国内仅限于王普昶等[6]利用ISSR分子标记技术对6个野生地理分布区华北驼绒藜遗传多样性和遗传分化进行鉴定,表明这种变异主要存在于种间;王普昶等[7]研究了华北驼绒藜种群空间分布格局与环境的种群空间分布格局,表明种群盖度、密度、人为干扰等15个环境因子是引起其种群分布格局的主要因素;而易津等[8]通过对不同储存温度和包装方式的处理研究了2者对种子活力的影响,低温可显著延长种子寿命,使种子保持较高活力;孙海莲等[9]研究了华北驼绒藜关键成苗技术及大田管理建植技术以及其他方面的研究,而关于华北驼绒藜种子和幼苗阶段环境适应机制至今还未见报道。本研究通过测定光照、温度、盐分和干旱胁迫对华北驼绒藜种子萌发的影响,探究其对荒漠环境的适应机制,进而为华北驼绒藜植被恢复和优质牧草开发利用提供科学依据。
1 材料与方法
1.1 种子的采集
完全成熟的华北驼绒藜种子于2013年10月采集于内蒙古农牧科学研究院四子王旗基地西南4 km附近(41°44′N, 111°52′E, 海拔为1 400 m)的华北驼绒藜野生群落中。在种子收获1周内使用手工剥离法脱去苞片获得种子,用于测定萌发特性,而未使用的种子置于-20 ℃下干燥保存。
1.2 种子的大小和重量的测定
随机选取华北驼绒藜带苞片和无苞片种子各1 000粒为1组,用万分之一电子天平称量种子千粒重,取5组重复测量的平均值。随机选择20粒无苞片种子,用游标卡尺测定其长度和直径。其中具苞片的种子千粒重为(2.30±0.04) g ,去苞片种子千粒重为(1.29±0.02) g。去苞片种子直径和长度分别为(1.57±0.05) mm和(2.48±0.08) mm。
1.3 种子吸水特性的测定
种子吸水试验在实验室条件(温度:20~25 ℃,相对湿度:40%~50%)下进行。每重复25粒种子,共5个重复。将待测种子置于盛有3 ML蒸馏水且底部垫有2层滤纸的培养皿后,开始计时。分别在种子吸水时间0.5、1、2、3、4、5、6、7、8 h时将种子取出,快速用干燥滤纸擦干种子表面水分,置于万分之一天平称重,再放回培养皿中,直至8 h为止。种子吸水率为W,将种子原质量定义为W1,不同时间段种子吸水后的质量定为W2,则:
(1)
1.4 光照和温度对种子萌发的影响
种子萌发试验在鄂尔多斯生态站的培养箱(江南仪器GXZ-0328,100 μmol·m-2·s-1))中进行。将25粒完好种子置于直径为50 mm且垫有2层滤纸的培养皿中,用移液管滴加3 ML蒸馏水,再用6号自封袋将培养皿封装。每个处理设置4个重复。本试验的5个变温梯度分别为5/15、10/20、15/25、20/30、25/35 ℃(12 h黑暗/12 h光照)和6个恒温梯度5、10、15、20、25和30 ℃(全光照),代表生境中不同月份的最低/最高气温:其中5/15℃代表生境中4月和10月的气温,10/20 ℃代表5月和9月的气温,15/25 ℃代表6月和8月的气温,20/30 ℃代表7月上旬的气温,25/35 ℃代表7 月下旬的气温(参考鄂尔多斯生态站检测数据)。光照条件分别为:12 h黑暗/12 h光照及全黑暗和恒温全光照。全黑暗处理是将装有种子的培养皿置于黑色完全不透光的暗袋中。每天统计萌发率,全黑暗条件在完全黑暗安全绿光条件下观察,以胚根突破种皮1 mm为萌发标准,共统计10 d。为检测种子的萌发特性,计算不同温度、光照条件下的最终萌发率(Germination percentage,GP,%)、发芽指数(Germination index,GI)、萌发速率(Germination rate,GR)、平均萌发时间(Mean germination time,MGT,d)。各指标的计算公式分别为:
(2)
式中:a为萌发种子数;c为供试种子数。
(3)
式中:Gt为在时间T时的发芽数;Dt为发芽日数。
(4)
式中:Tt为相应的萌发日数;Gt为t时间的萌发百分数。
(5)
式中:G为每天萌发数;T为从萌发开始到结束时的天数;N为供试种子数。
1.5 等渗NaCl和PEG溶液对种子萌发和恢复萌发的影响
盐分和干旱梯度采用不同浓度的NaCl和与其等渗的PEG-6000进行设定。NaCl溶液的梯度设置为0、0.2、0.4、0.6、0.8 mol·L-1,按照LANG[10]的NaCl换算公式得到与其相对应的渗透势为0、-0.7、-1.4、-2、-2.7 MPa,再按照MICHEL等[11]的计算公式换算为等渗的PEG溶液。
为了防止PEG-6000随温度变化而产生渗透势变化,选择在恒温20 ℃下进行试验,试验方法同1.4恒温萌发特性试验。初始萌发10 d后,将未萌发的种子用蒸馏水清洗2次,移至盛有蒸馏水的培养皿中继续光照培养10 d。计算萌发恢复率、初始萌发率(同公式2)和最终总萌发率的公式分别为:
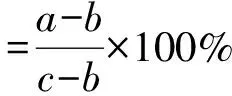
(6)
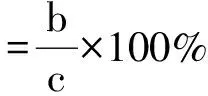
(7)
式中:a为在盐溶液中和恢复萌发过程中萌发的总种子数;b为在盐溶液中萌发的种子数;c为供试总种子数。种子生活力的判定方法为: 观察种子外形完好度、种胚颜色和坚实度,结合四唑染色法测定结果判断,将染色后胚为红色的视为有活力种子[3]。
1.6 数据处理
为保证方差齐性,数据分析之前先进行方差异质性检验,对于方差不齐的数据进行反正弦转换。采用单、双因素方差分析比较各处理之间的差异(5%水平);如果差异显著,再采用Tukey’sHSD方法进行多重比较。数据分析用SPSS17.0软件进行,图表中种子萌发率均采用平均值±标准误差来表示。
2 结果与分析
2.1 种子吸水特性
种子吸水0.5h后种子质量增加(20.39±2.25)%,在1h内吸水迅速,种子吸水率可达(36.46±2.11)%;1h后种子吸水率随时间呈直线上升趋势(吸水率=16.08×时间+16.31,R2=93.3%,P<0.001,图1)。在2h后出现萌发,种子质量增加53.60±3.32%。8h后种子萌发率达到(40±2.17)%,种子质量增加(122.37±2.76)%。
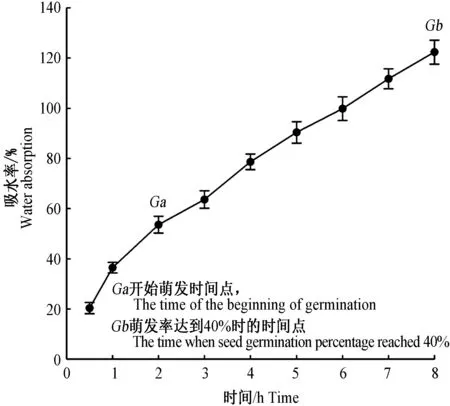
图1 实验室条件(20~25 ℃、45%~50%相对湿度)华北驼绒藜种子吸水率曲线
2.2 光照、温度对华北驼绒藜种子萌发特性的影响
双因素方差分析表明,光照条件对种子GP、GI、GR和MGT均无显著影响(表1、图2、图3)。不同变温条件对GP没有显著影响(F1,30=1.15,P=0.351)(表1),种子均达到相同的累积萌发率,高达95%以上 (图2)。同时,温度和光照2者交互作用对种子累积萌发率仍无显著影响(F4,30=0.72,P=0.587)(表1)。然而,GI(F4,30=8.31,P<0.001)、GR(F4,30=7.29,P<0.001)和MGT(F4,30=60.32,P<0.001)均受到温度的显著影响(表1、图3)。其中,5/15℃处理的GI、GR值显著低于其他变温温度处理,而MGT值显著高于其他处理,而且变温对华北驼绒藜种子萌发特性的影响不依赖于光照条件(表1、图3)。
同样,在不同的恒温条件下,种子也均达到相同的累积萌发率(F5,18=1.62,P=0.206) (图4A)。不同恒温条件对种子GI(F5,18=102.46,P<0.001)、GR(F5,18=194.69,P<0.001)和MGT(F5,18=21.44,P<0.001)有显著影响(图4B、C、D)。在5℃恒温处理下种子GI,GR值显著低于其他温度处理,而MGT值高于其他温度处理(图4-B、C、D)。
综合以上结果,华北驼绒藜种子在各个温度与光照处理中均有相当高的萌发率(95%~100%),因此该种子不具休眠性。
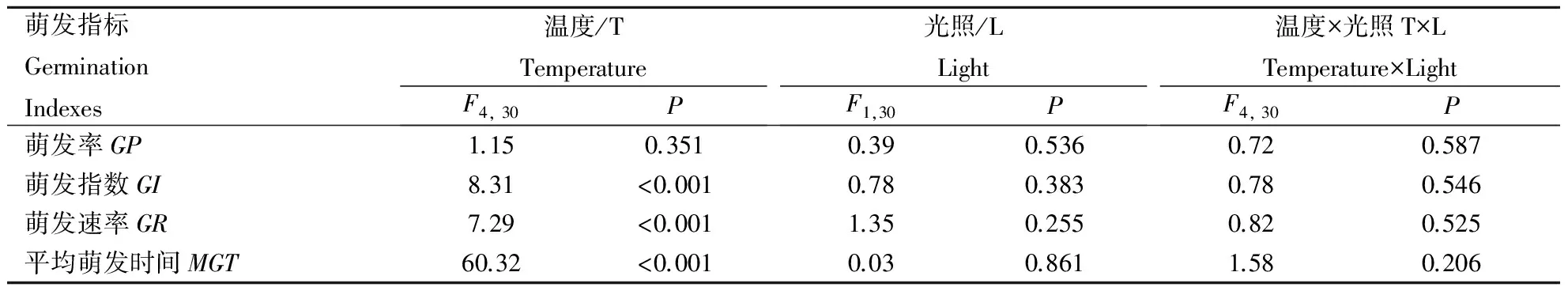
表1 温度、光照及其交互作用对华北驼绒藜(Ceratoides arborescens)种子萌发指标的影响
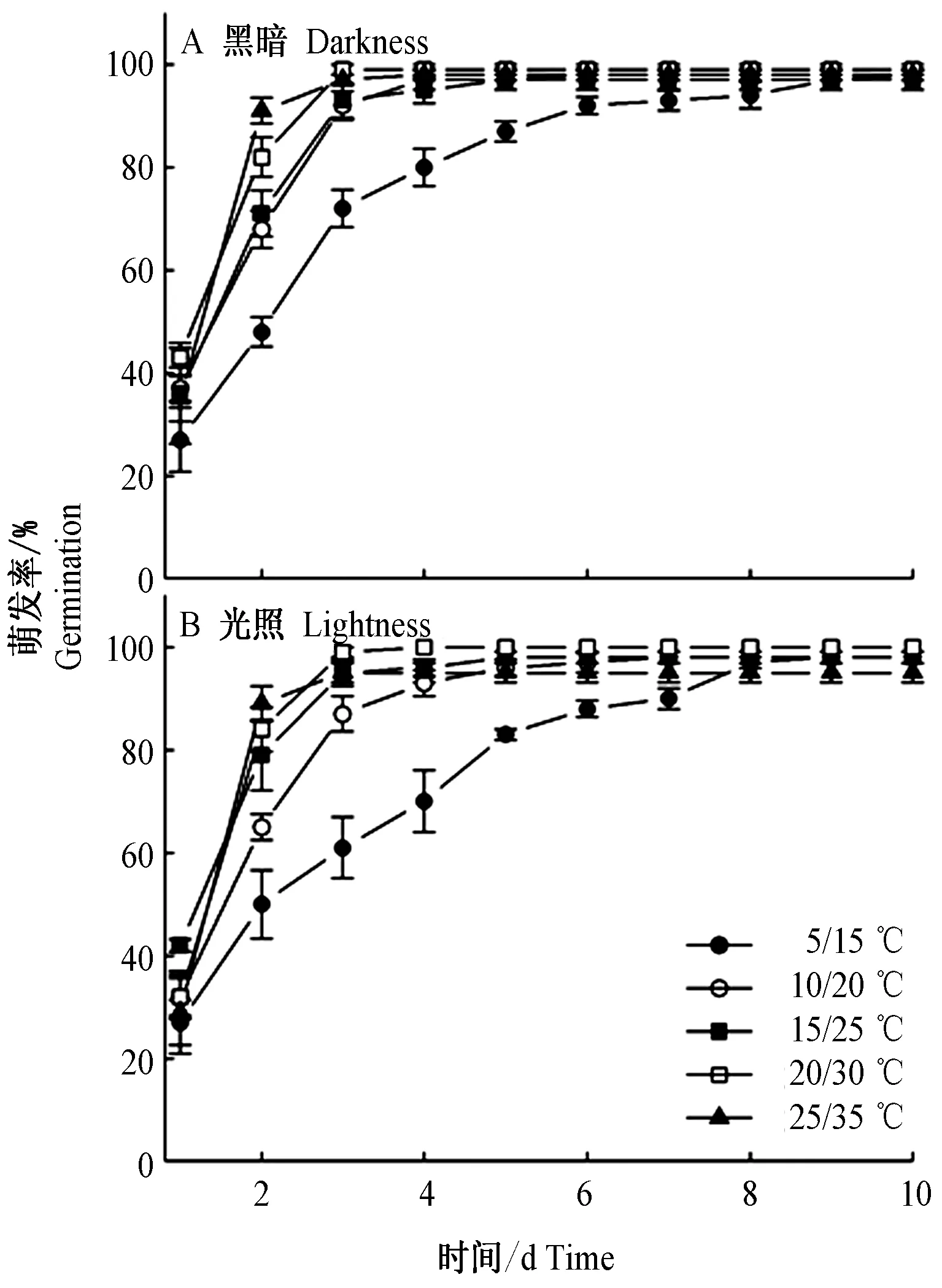
图2 温度和光照对华北驼绒藜种子日累计萌发率的影响
2.3 等渗NaCl和PEG溶液对种子萌发的影响

注:数据为平均值±标准误差,不同字母表示不同萌发指标之间的差异显著(P<0.01) 。下同。
Note: Data are mean±SE. Different letters indicate significant differences between different seed ripening times(P<0.01) .The same as below.
图3 温度和光照对华北驼绒藜种子萌发指标的影响
Fig.3 Effects of light and temperature on germination indexes ofCeratoidesarborescensseeds
华北驼绒藜种子在NaCl和PEG溶液中培养10 d后,萌发情况如图6所示。随着等渗NaCl和PEG溶液渗透势降低,初始萌发率表现为:在0~-1.4 MPa下降缓慢,-1.4~-2.0 MPa下降迅速,直至在- 2.7 MPa处理下为0(图6)。
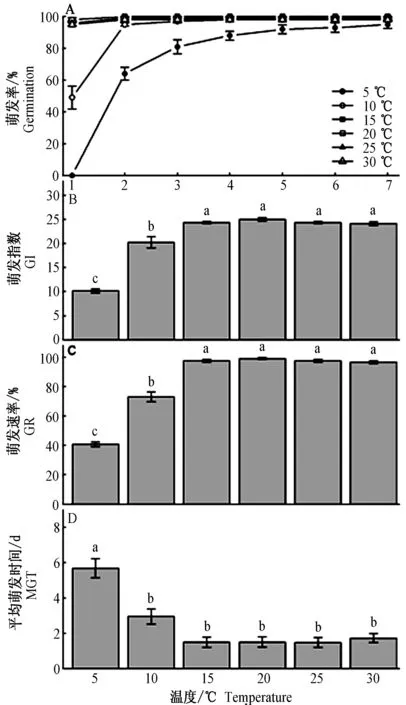
图4 不同恒温条件对华北驼绒藜种子日累计萌发率(A)和萌发指标的影响(B~D)

图5 等渗NaCl和PEG对华北驼绒藜萌发指数的影响
双因素方差分析结果表明,等渗NaCl和PEG溶液处理下对初始萌发结果的影响无显著差异(F1,30=0.43,P=0.520),而不同渗透势对种子GI有显著影响(F4,30=369.56,P<0.001 )。随着两种溶液渗透势的降低,种子GI显著下降(图5)。
在不同浓度的溶液中未萌发的种子转移至蒸馏水中的萌发恢复情况如图6所示。相对高渗透势的NaCl溶液保持着较高的恢复萌发率,可达到91%,而低渗透势 -2.7 MPa的NaCl溶液处理后,恢复萌发率降低至30%。PEG渗透势在 -0.7~-2.7 MPa梯度上,华北驼绒藜种子恢复萌发率相对平稳,能维持在60%~80%。PEG引起的干旱胁迫对最终萌发率影响变小,而NaCl引起的盐分胁迫对华北驼绒藜种子的最终萌发率影响变大,较低渗透势(-2.7 MPa)的NaCl溶液培养的种子死亡率高出与其等渗的PEG溶液处理51%(图6)。
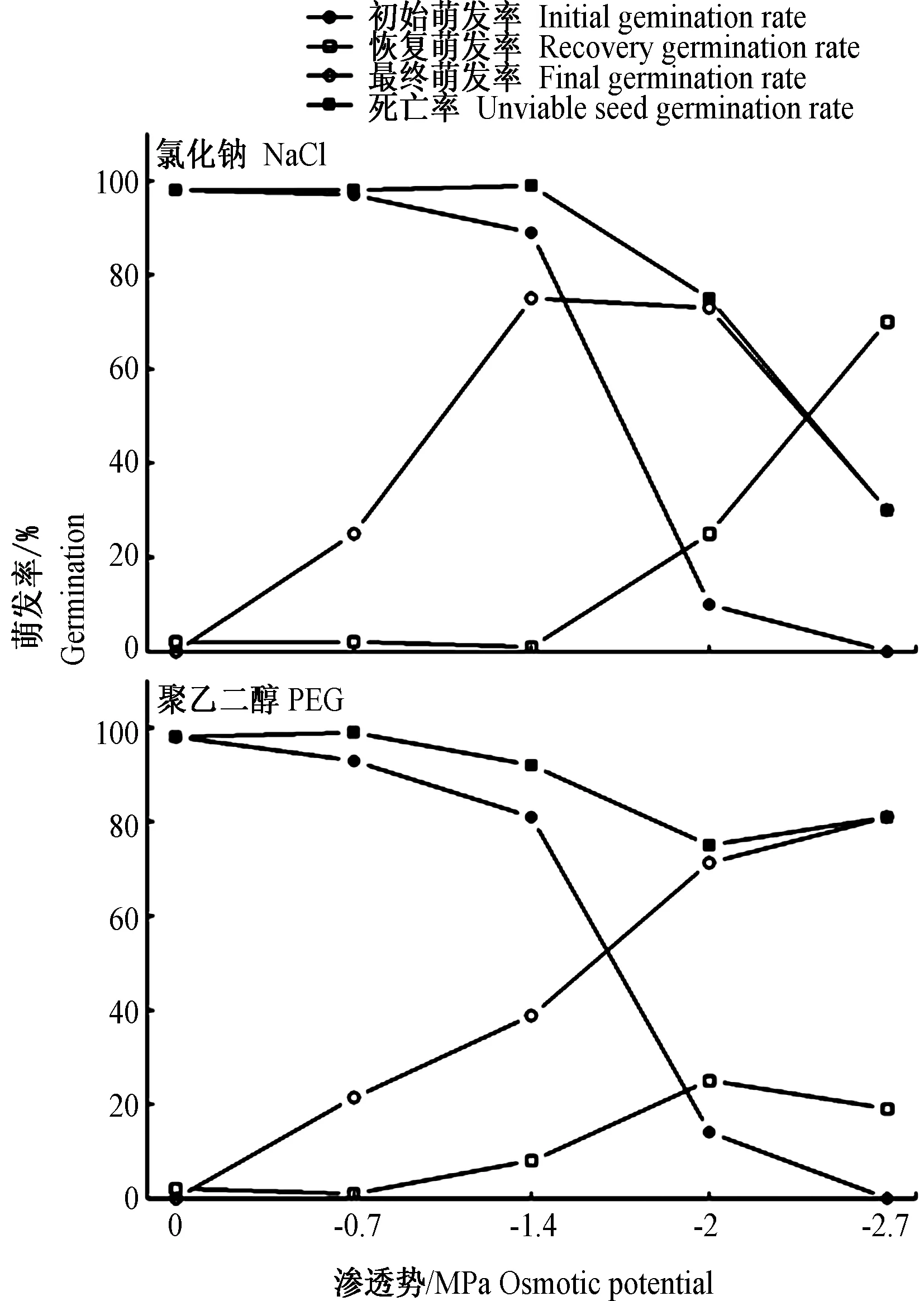
图6 等渗NaCl溶液和PEG溶液对华北驼绒藜(Ceratoides arborescens)种子萌发、恢复萌发率和最终萌发率的影响
3 结论与讨论
在荒漠或半荒漠地区,植物种子萌发对光照有着不同的响应。有些物种是在光照条件下萌发能力更高,如白沙蒿[12],而有些物种则是在黑暗条件下具有较高的萌发率[3],如沙蓬[13]、沙芥和斧翅沙芥[14]等。本试验结果表明,华北驼绒藜种子萌发与同科无休眠物种囊果碱蓬[15]、梭梭[16]等其他灌木物种相同,即其萌发在黑暗中和光照条件下的萌发能力没有差异。
温度对荒漠植物种子萌发有重要的调节作用,它影响着物种的萌发速率等指标。本试验结果表明,温度变化对华北驼绒藜种子萌发没有显著影响,华北驼绒藜种子有较宽的萌发温度范围 (5~30 ℃),即说明其可能在任何一个生长季节萌发。但是,低温(5/15和5 ℃)推迟了种子平均萌发时间,萌发速率和发芽指数也显著降低。
荒漠或半荒漠地区的特点是少量而不可预测的降水和很大的潜在蒸发量。因此,物种在萌发过程中往往会遭受盐分和干旱的双重胁迫。本试验表明,华北驼绒藜种子萌发受到等渗NaCl和PEG胁迫时,会降低种子萌发率和萌发指数。随着等渗NaCl和PEG渗透势的降低,种子萌发率呈降低趋势,而在高浓度处理下具有较高萌发率。在-2 MPa等渗NaCl和PEG溶液中仍具有萌发能力(萌发率分别为10%和14%),表明华北驼绒藜有较强的抗旱、抗盐能力。
盐分对种子萌发的影响一般归结为渗透胁迫与离子毒害,而PEG 溶液处理对种子萌发的影响仅表现为渗透胁迫。通过对种子恢复萌发试验,低浓度溶液(-0.7~-2 MPa)中的种子具有较高恢复萌发率,这表明是溶液的渗透压抑制了种子的萌发而不是金属离子毒害了种子,这种现象在同科植物梭梭[16],紫翅猪毛菜[17]中也有出现。而在高浓度(-2.7 MPa)处理下,种子在等渗NaCl和PEG溶液之间的恢复萌发率有显著差异,PEG溶液恢复率要高于等渗NaCl溶液51%,因此高浓度处理中NaCl溶液对种子萌发不仅产生了渗透胁迫,也有离子毒害。因而,华北驼绒藜种子具有较强的抗旱性,在水分缺乏的荒漠地区,受到渗透胁迫的影响时,仍能保持一定的萌发率。在雨水降低土壤溶液渗透势后,能够解除胁迫,继续恢复萌发,最终能够顺利建立种群。
植物在长期的进化过程中,形成独特的方式以适应环境的变化,从而保证种群的建成和生存。华北驼绒藜具有高萌发率(95%~100%)、萌发迅速(恒温15 ℃~30 ℃条件下MGT仅仅需2 d)、单株种子个体小而量大(21 992粒[18])且种子寿命短(自然条件下,种子寿命仅为8~10个月,常规贮存一年的种子,70%以上丧失发芽力[19])、萌发对温度及光照不敏感、不具休眠性等特性,而这些特征属于典型的“R对策”[20],这种萌发策略有利于适应荒漠、半荒漠地区多变的环境。温度、光照、土壤盐分和干旱等因素共同决定了华北驼绒藜种子的不同萌发策略,而这种响应也正是华北驼绒藜物种对荒漠、半荒漠、干旱等异质性环境中的存活适应。
[1] HARPER J L. Population biology of plants [M].New York, San Francisco, London: Academic Press,1977.
[2] GUTTERMAN Y. Seed Germination in Desert Plants [M]. New York: Springer-Verlag,1993.
[3] BASKIN C C, BASKIN J M. Seeds, Ecology, Biogeography, and Evolution of Dormancy and Germination[M]. San Diego: Academic Press,1998.
[4] 黄振英,曹 敏,刘志民,等.种子生态学:种子在群落中的作用[J].植物生态学报, 2012,36 (8):705-707.
[5] 马毓泉.内蒙古植物志[M].呼和浩特:内蒙古人民出版社,1979.
[6] 王普昶,赵丽丽,易 津,等.内蒙古野生华北驼绒藜种群遗传多样性和遗传分化研究[J]. 草业学报, 2011,20(4):287-292.
[7] 王普昶,易 津,赵丽丽,等.华北驼绒藜种群空间分布格局及其环境依赖性研究[J].种子,2009,28(3):5-9.
[8] 易 津,曹自成,乌仁其木格. 几种不同贮藏条件对华北驼绒藜种子寿命和活力的影响[J]. 内蒙古农牧学院学报, 1994,15(1):18-22.
[9] 孙海莲.华北驼绒藜成苗特性与建植技术研究[D].呼和浩特:内蒙古农业大学, 2007.
[10]LANG ARG. Osmotic coefficients and water potentials of sodium chloride solutions from 0 to 40°C[J].Australian Journal of Chemistry, 1967,20(9):2017-2023.
[11]MICHEL B E, KAUFMANN M R. The osmotic potential of polyethylene glycol-6000[J]. Plant physiology, 1973,51(5):914-916.
[12]YANG X, BASKIN C C , BASKIN J M, et al. Seed mucilage improves seedling emergence of a sand desert shrub[J]. PLoS ONE, 2012,7(4):e34597.
[13]GAO R, YANG X, YANG F, et al. Aerial and soil seed banks enable populations of a desert annual to cope with an unpredictable environment[J]. Annals of Botany , 2014,114:279-287.
[14]宋兆伟,郝丽珍,黄振英,等.光照和温度对沙芥和斧翅沙芥植物种子萌发的影响[J].生态学报, 2010,30(10): 2562-2568.
[15]王 雷,田长彦,张道远,等.光照, 温度和盐分对囊果碱蓬种子萌发的影响[J].干旱区地理,2005, 28(5):670-674.
[16]黄振英,张新时,GUTTERMAN Y,等.光照、温度和盐分对梭梭种子萌发的影响[J]. 植物生理学报, 2001,27(3):275-280.
[17]魏 岩.准噶尔荒漠几种藜科植物的种子多型性及其萌发行为研究[D].乌鲁木齐:新疆农业大学, 2007.
[18]王学敏,易 津,张 鹏.管理方式对华北驼绒藜生长及种子产量、质量的影响[J].草地学报,2003, 11(2):139-145.
[19]王普昶,易 津,赵丽丽.华北驼绒藜种群数量动态与生殖特性的相关性[J].生态学杂志, 2010,29(6):1081-1086.
[20]VENABLE D L. The evolutionary ecology of seed heteromorphism[J]. The American Naturalist. 1985,126:577-595.
(责任编辑:李 莹)
Effects of temperature, light, salt and drought stress on seed germination ofCeratoidesarborescens
HE Qingshan1,2, WEI Lingling1, ZHANG Xu1, LIU Guofang2, YANG Huiling1,2
(1.College of Life Science,Henan Agricultural University, Zhengzhou 450002, China; 2.State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China)
We tested the effects of temperature, light and osmotic stresses (NaCl and PEG) on seed germination ofC.arborescensto investigate the adaption strategy in desert habitat. The results showed that, seed coat ofC.arborescenswas water-permeable and the seed began to germinate after 2 hours of water absorption. The germination rate (GR), germination index (GI), and mean germination time (MGT) decreased more significantly in low temperature conditions (e.g. 5/15, 5 ℃) than in the other temperature regimes, and its seed germination rate could reach an extremely high value (95%~100%) in all tested temperature regimes under both light and dark conditions. Therefore,C.arborescenswas considered as a non-dormant species. With initial germination percentage decreased with the decrease of osmotic potential, germination was not affected by osmotic potential (NaCl and PEG) at concentrations higher than -1.4 MPa; but the germination index was significantly reduced with the increase of osmotic potential; The seed germination rate was decreased to 10%-14% when osmotic potential was lower than 2.0 MPa. Germination percentage in high osmotic potentials concentrations(0~-2.0 MPa)has high recovery germination. But the seed mortality percentage under -2.7 MPa NaCl was higher than that under PEG with the same osmotic potential by 51%, which indicates that some seeds have lost their viability in high NaCl solution. Thus,C.arborescensowns relatively faster seed germination fraits and drought resistance that coutribute to the species adaption and persistence to the arid desert environment.
Ceratoidesarborescens; seed germination; osmotic potential; temperature; light
2014-12-28
“十二五”国家科技支撑计划课题(2012BAD16B03),(2012BAC01B05)
何青山(1989-),男,河南信阳人,硕士研究生,主要从事种子生态学方面的研究。
杨慧玲(1976-),女,河南郑州人,副教授,博士;刘国方(1980-),男,河南驻马店人,副研究员,博士。
1000-2340(2015)03-0343-06
S 548
A
