太行山低山丘陵区3种树木叶温对土壤水分胁迫的响应
2015-07-01杨文卿张志铭杨喜田
杨文卿, 张志铭, 侯 娜, 赵 勇, 王 谦, 杨喜田, 樊 巍
(1.河南农业大学林学院,河南 郑州 450002; 2.河南省林业科学院,河南 郑州 450008)
太行山低山丘陵区3种树木叶温对土壤水分胁迫的响应
杨文卿1, 张志铭1, 侯 娜1, 赵 勇1, 王 谦1, 杨喜田1, 樊 巍2
(1.河南农业大学林学院,河南 郑州 450002; 2.河南省林业科学院,河南 郑州 450008)
以侧柏、刺槐和栓皮栎为试验材料,通过测定3种树木的叶温及气温,研究了3种树木在水分胁迫下叶温的日变化规律。结果表明,(1)侧柏、刺槐和栓皮栎的叶温日动态对土壤水分变化的响应十分显著。表现出随着水分胁迫的加剧,叶温呈升高趋势;重度水分胁迫下,叶温值在各个时间段都明显偏高。(2)3种树木在不同的水分胁迫下,叶温与气温日变化趋势表现一致,均为早晚低,午间高;且叶温均低于气温;但在重度水分胁迫下的10:00-16:00时,栓皮栎的叶温高于气温。(3)3种树木的叶温随气温变化的各有其特点,侧柏和栓皮栎呈“单峰型”曲线,而刺槐表现为“双峰型”曲线。
侧柏;刺槐;栓皮栎;叶温;水分胁迫
由于中国华北石质山区降水偏少、降水分布不均、土层浅薄,水分成为这一地区植物生长的限制因素。土壤水分的不足成为太行山低山丘陵地区植被恢复的主要障碍,即使是耐旱的造林先锋树种,如栓皮栎、侧柏、刺槐也常常受到土壤水分的胁迫,导致生长缓慢[1]。植物的叶面温度(简称叶温)作为植物体的一个重要生理特性,被认为是植物生理生态研究中的一个基本参数[2]。叶温的变化体现了叶片在外部环境变化和内部新陈代谢共同作用下叶片能量平衡的结果[3]。研究认为叶温与光合作用相关,叶面温度的变化会影响到光合作用,而叶片的结构和光合特性又受到环境条件的显著影响[4,5]。其次,植物叶温因叶片(如朝向、叶脉网络分布)的排列方向不同,受光照及风的影响也有差异,其热量明显不同,所以不仅不同的叶子温度不同,甚至同一片叶子的不同部位温度也有差异[6]。中国对叶温的研究起步较晚,多集中在农作物方面,并且多是集中在探讨冠层温度的变化,对于单叶片温度变化的研究较少[7,8]。因此,开展植物叶温的相关研究,探讨叶温在水分胁迫下的变化特性,对了解植物的生理生态规律有着重要意义。本研究以太行山地区常见的造林树种侧柏、刺槐和栓皮栎为对象,通过研究三种植物叶温在不同水分胁迫处理下的变化,讨论了叶温变化与气温日变化的响应关系,以期为太行山低山丘陵区的植被恢复和抗旱物种培育提供理论依据。
1 材料与方法
1.1 试验材料
试验地位于济源市大沟河林场的国家林业局小浪底生态站。以侧柏、刺槐、栓皮栎为测试树种。选择生长良好,长势一致的植物各20盆,苗龄3年。盆高50 cm,口径45 cm,土层深度40 cm(土壤为褐土),每盆栽1株植物。
1.2 试验方法
1.2.1 土壤含水量的测定与控制 2014-06-21开始对3种树木进行土壤水分控制。将每个树种的20盆分为4组(即4个处理),每个处理5个重复。按照土壤含水量为田间最大持水量的75%~80%(正常水分,对照组CK)、60%~65%(轻度胁迫,LS)、45%~50%(中度胁迫,MS)、20%~25%(重度胁迫,SS)进行控水处理。人工控制盆栽的浇水量,以获得不同的土壤水分梯度。利用时域反射仪(Time Domain Reflectometry,TDR) 测量土壤含水量,测量时将探头直接插入土壤,插入深度为10、20、30 cm处以测得土壤水分。每层土壤重复测量3次。田间持水量采用土壤环刀法测量。
1.2.2 叶温的测定 连续控水20 d 后,每个土壤水分梯度下选取3盆植株,对同一植株中底部不同方位的2片叶子进行测量,取其平均值。植株叶片的测量位置为完全展开且阳光能够充分照射的水平叶片正面。选择晴朗天气,从8:30到17:30,采用CB-0231热电偶测温仪,对3种树木的叶片温度进行测量,每1.5h测量并记录数据1次。
1.3 数据统计分析
采用Excel 2007和SPSS17.0软件对试验数据进行分析处理。
2 结果与分析
2.1 树木叶温对水分胁迫的响应
从图1可以看出,不同程度土壤水分胁迫下,3种树木叶温日变化基本趋势大致为早晚低,午间高。其中叶温在一日内的变化侧柏、栓皮栎为单峰型曲线,刺槐为双峰型曲线;随土壤胁迫性增大,叶片温度逐渐升高。叶温值在正常供水情况下的各个时段均最低;而重度水分胁迫下均最高。晴天侧柏、刺槐和栓皮栎3种树木叶温的日均值随着土壤水分胁迫程度增加均表现出逐渐增加的趋势。在正常供水下(对照)侧柏的叶温日均值为31.79 ℃,在轻度和中度水分胁迫下较对照分别升高0.8%和1.7%;重度水分胁迫的叶温日均值较对照升高2.4%为32.57 ℃。重度水分胁迫下,刺槐和栓皮栎的叶温日均值分别较对照处理升高2.6%和5.1%。综上分析,随着水分胁迫的加剧,针叶树种侧柏叶温升幅较小,阔叶树种刺槐和栓皮栎的叶温升幅比侧柏大。方差分析可知,3种树木各水分胁迫处理间均表现为显著性差异,即3种树木叶温均对水分胁迫响应显著。
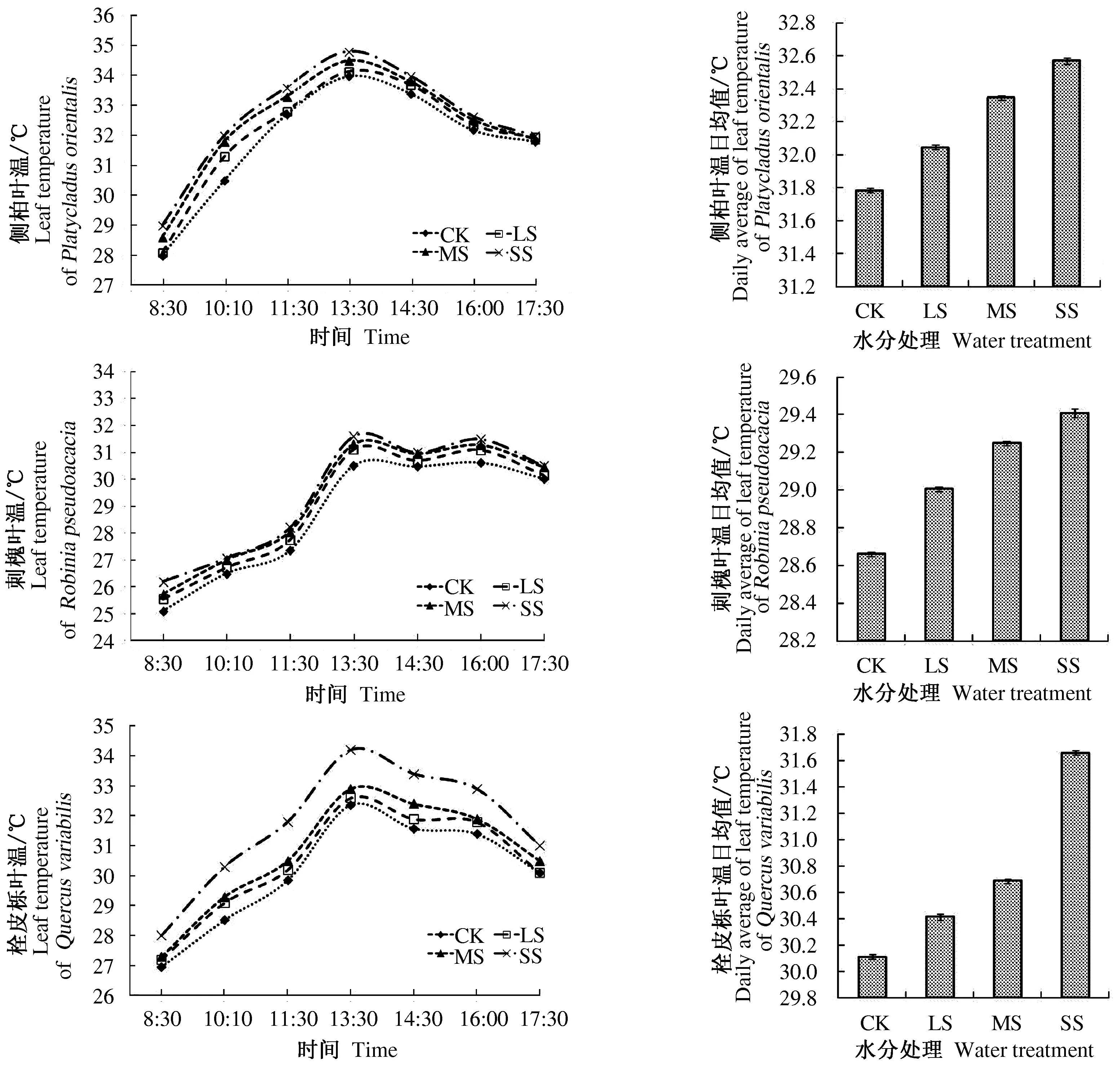
图1 3种树木叶温和叶温日均值对水分胁迫的响应
3种树木的叶温日变化在各个水分处理间均表现明显,3种树木的叶温和叶温日均值均呈现出随水分胁迫程度增加而升高的趋势。叶温对水分胁迫响应十分显著,尤其在10∶00~16∶00之间。因此,在同一时间和相近的环境条件下,叶温变化可以作为3种树木受到土壤水分胁迫程度的重要响应指标之一。
图1中显示,刺槐的叶温日变化区别于侧柏和栓皮栎的“单峰型”曲线,其叶温变化呈“双峰型”曲线;第1个峰出现在13∶00,第2个峰出现在16∶00。刺槐的这种叶温日变化特点类似一些植物的光合“午休”的现象。在其他条件一致时,土壤水分胁迫越高叶温越高,这可能是因为植物叶片吸收太阳辐射后将一部分能量转换成热能。当气温升高促使植物蒸腾加快时,如果植物供水充足,土壤水分能够满足植物叶片蒸腾的需要,使叶温下降且此时温度低于其在蒸腾抑制时;但当植物受到水分胁迫时,叶片的保卫细胞会对气孔起到调节作用,此时蒸腾作用减少,相对于土壤水分充足的叶片,植物蒸腾的潜热减少,显热增加,叶温相应上升。刺槐叶温的这种类似植物光合“午休”的特点也进一步说明植物的叶温与植物蒸腾有着密切的关联。
2.2 气温日变化对树木叶温的影响
通过对晴天对应高度气温的同步观测发现,由图2可知,1 d中,侧柏、刺槐和栓皮栎3种树木的叶温随时间的变化与周围气温随时间的变化趋势是一致的,有着明显的起伏变化规律,都呈现近似的单峰和双峰曲线。随着太阳升起,气温逐渐升高,叶温也逐渐增大。在13∶00左右,气温和3种树木的叶温均达到最大值。达到极值后,气温和叶温开始逐渐降低,但气温和3种树木的叶温在8∶30左右均低于17∶30。从图2可知, 随着水分胁迫程度的加剧,3种树木的叶温日均值逐渐升高,并且逐渐接近气温日均值,但是3种树木各个水分处理的叶温日均值均低于气温日均值;在重度土壤水分胁迫下,栓皮栎的叶温日均值最接近气温日均值。
由图2可知,1 d内3种树木的叶气温差也随时间的变化而变动。3种树木在上午的叶气温差普遍高于下午。随着上午气温的快速上升,叶气温差逐步缩小;下午气温缓慢下降,叶气温差绝对值变化相对平缓。同时随着水分胁迫程度的加剧,叶气温差呈现正常供水>轻度胁迫>中度胁迫>重度胁迫的特点。在重度水分胁迫的情况下,侧柏在中午气温最高点处的叶气温差基本趋近于0,栓皮栎在重度水分胁迫下在11∶00左右叶温即超出气温为负值。刺槐的叶温变化规律呈“双峰型”曲线,其叶温变化出现类似光合“午休”的现象,因此其叶气温差在出现“午休”期间较大。重度水分胁迫条件下,刺槐叶气温差最小的时点并不在气温最高的正午,而在类似光合“午休”现象结束后的第二个峰。3种树木在叶气温差的表现大致相近但各有特点。
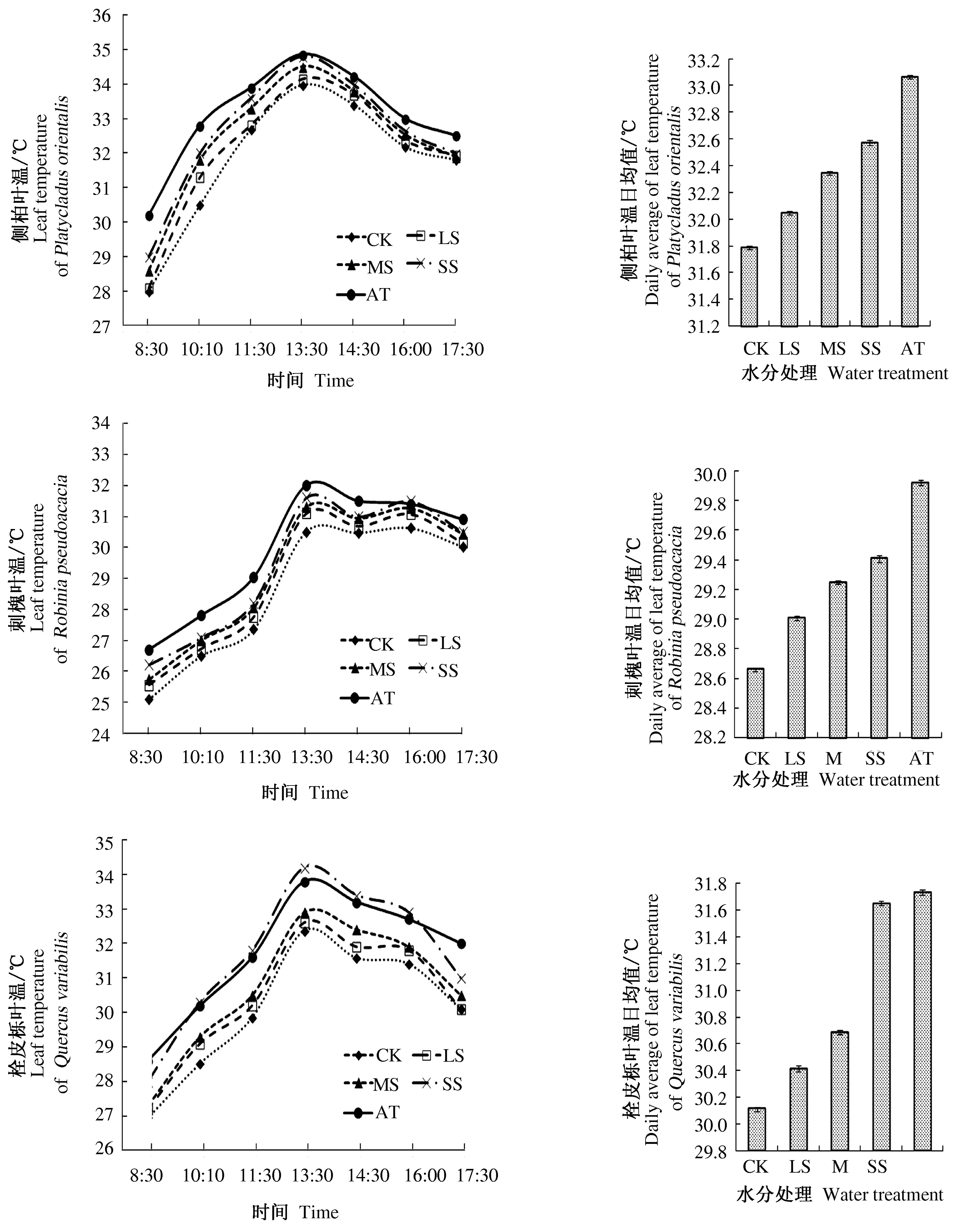
图2 3种树木叶温和空气温度的日变化
通常情况下,植物的叶温一般低于晴天白天的空气温度。当气温升高,植物通过蒸腾作用保持叶温使其低于气温,避免叶片受到灼伤。但当光照过强、气温过高或受到水分胁迫时,植物叶片会关闭部分气孔以减少蒸腾,此时植物叶温将可能高于气温。
植物对于叶温保持能力的不同代表着其对水分胁迫的耐受程度。当植物叶温高于气温说明该植物水份状况或保持叶温的能力较差,对水分胁迫环境的适应能力不足。因此,在重度水分胁迫下,在10∶00至16∶00之间栓皮栎叶温高于气温,而侧柏与刺槐的叶温并未高于气温,可能是由于相对侧柏和刺槐而言栓皮栎不耐受水分胁迫。
3 结论与讨论
1)侧柏、刺槐和栓皮栎叶温日变化为早晚低,午间高。其中,侧柏和栓皮栎叶温日变化呈“单峰型”曲线,刺槐叶温日变化呈“双峰型”曲线。
2)3个树种的叶温对土壤水分变化的响应十分明显,正常供水情况下的叶温值在各个时段均最低;而在重度水分胁迫下,各个时段的叶温值均最高。
3)在水分胁迫下侧柏、刺槐和栓皮栎的叶温日变化曲线均低于气温的日变化曲线,只有在重度水分胁迫下在10∶00至16∶00之间栓皮栎的叶温高于气温。在不同土壤水分处理下,侧柏、刺槐和栓皮栎3种树木的叶温随时间的变化与周围气温随时间的变化趋势是一致的,但不同树种在具体表现上存在差异。
3种树木叶温都表现为随土壤水分胁迫的加剧而升高的趋势。豆胜等[9]研究表明,叶温的变化可以随着水分的变化而变化。当水分供应减少时,叶片温度就会上升。因此其他条件一致时,叶温可以作为这3种树木土壤水分状况的的反应指标之一。姚振坤等[10]研究表明,在进一步的试验中可以根据3种植物的主要生理特征和环境特点,建立通过叶温判断植物水分胁迫程度的模型,以方便监测植物受土壤水分胁迫的程度,进而采取合理的抗旱措施,提高保育效果。
侧柏和栓皮栎各个水份处理叶温日变化的为“单峰型”曲线,这与宋晓明等[11]、董果等[12]研究结果一致。而刺槐的叶温日变化规律呈“双峰型”曲线。谢心等[13]研究表明,在日照强烈的中午,由于植物叶片的过量蒸腾,使植物体内的水分迅速下降,于是叶片的气孔便会暂时关闭或半关闭,以抑制体内水分的损失强度,从而以减少光合物质为代价换取减小水分胁迫的伤害,这样便形成了光合“午休”的现象。在本研究中,刺槐的叶温日变化表现出类似光合“午休”的现象。综上分析,在通过叶温判断植物水分胁迫的实践中,应根据具体树种的叶温变化特点选择合适的测量时间。
叶温存在时空间的变异性,不同生长阶段、地理环境、植株之间的表现不同,即使同一植株,不同部位的叶面温度也存在差异。以往对叶温与气温之间关系的研究也有很多,但并没有一致的结论。李强征等[14]研究发现充分供水和水分亏缺条件下温室内黄瓜叶温变化的差异及其与相关作物生理信息的关系结果表明,叶温与叶面蒸腾的 Pearson 相关系数达到了0.7以上叶面蒸腾是影响叶温变化的内在因素供水条件的不同影响了作物蒸腾的变化从而导致作物叶温变化的差异。赵晓彤等[15]通过对开粳1号叶温、气温及生理性状的研究表明,叶温与气温变化一致,但叶温低于气温。水分胁迫严重的叶温始终高于轻度胁迫的叶温,并随控水时间的延长差距拉大,轻度水分胁迫叶温随控水时间变化不大,复水后各水分胁迫处理叶温基本相同。以上研究表明面对不同梯度的水分胁迫,不同物种呈现了不同的特征变化[14-17],而造成这种差异的原因可能与植物物种自身遗传特性以及其对环境变化的应变策略有关。
[1] 万 林.不同树种对华北石质山区困难立地水分限制的响应[D].北京:中国林业科学研究所, 2013.
[2] 赵立新,荆家海,王韶唐.陕西渭北旱源土壤-植物-大气连续体中水分运转规律的研究——I.生态环境对植物叶温的影响[J].西北植物学报,1996,16(4):345-350.
[3] 蔡焕杰,康绍忠.棉花冠层温度的变化规律及其用于缺水诊断研究[J].灌溉排水,1997,16(1):1-5.
[4] ANDERSON J M. Photoregulation of the composition,function and structure of thylakoid membranes[J].Annual Review of Plant Physiogy,1986,37: 93-136.
[5] BAKER N R. A Possible role for Photosystem II in environmental perturbations of photosynthesis[J]. Physiologia Plantarum, 1991,81:563-570.
[6] 蒋高明.植物的温度及测定[J].植物杂志,1998(1):30-31.
[7] 粱银丽,张成娥. 冠层温度-气温差与作物水分亏缺关系的研究明[J].生态农业研究,2000,8(1):24-26.
[8] 袁国富,罗 毅,孙晓敏,等.作物冠层表面温度诊断冬小麦水分胁迫的试验研究[J].农业工程学报,2002,l8(6):13-17.
[9] 豆 胜,马成仓, 陈登科. 4种常见双子叶植物蒸腾作用与叶温关系的研究[J].天津师范大学学报:自然科学版,2008,28(2):11-13.
[10]姚振坤, 罗新兰, 李天来, 等.日光温室番茄叶温的模拟及与环境因子的关系[J].江苏农业学报, 2010,26( 3 ): 587-592.
[11]宋晓明,董 果,赵 勇,等.土壤水分胁迫对栓皮栎叶温及叶绿素荧光特性的影响[J].河南农业大学学报, 2013,47(6): 691-697.
[12]董 果,戴 勐,赵 勇,等.侧柏叶温及叶绿素荧光特性对土壤水分胁迫的响应[J].中国水土保持科学, 2014,12( 1 ): 68-74.
[13]谢 心,赵 忠.不同土壤水分条件下刺槐幼苗光合日变化研究[J]. 陕西林业科技,2013(1):5-9.
[14]李强征,李国臣,于海业,等.温室内黄瓜叶温变化特性的试验研究[J].农机化研究, 2006 (8):144-146.
[15]赵晓彤,韩亚东,高继平,等.水稻穗分化期不同土壤水势叶温及生理性状变化[J].湖北农业科学,2011,50(1):33-36.
[16]刘云鹏,申 思,潘余强,等. 干旱胁迫下玉米叶-气温差与叶温差日变化特征及其品种差异[J]. 中国农业大学学报,2014,19(5):13-21.
[17]廖 金,周 泓,刘 冰, 等. 夏季城市绿地中不同遮阳条件下杜鹃叶温变化及其生理响应[J]. 北方园艺,2013(7):74-77.
(责任编辑:蒋国良)
Responses of leaf temperature of three types of trees to soil water stress in hilly areas of Taihang Mountain
YANG Wenqing1,ZHANG Zhiming1, HOU Na1, ZHAO Yong1, WANG Qian1,YANG Xitian1,FAN Wei2,
(1.College of Forestry,Henan Agricultural University, Zhengzhou 450002,China; 2.Henan Forestry Academy, Zhengzhou 450008,China)
The experiment was conducted withPlatycladusorientalis,RobiniapseudoacaciaandQuercusvariabili, the leaf temperature and air temperature were measured. In this paper, the daily variation and responding mechanism of the leaf temperature of the three types of trees under water stress was analysed and discussed systematically. The results indicated that: (1) the daily dynamic of the leaf temperature ofPlatycladusorientalis,RobiniapseudoacaciaandQuercusvariabiliwas remarkably responding to the variation of the soil moisture. The leaf temperature of all the three types of trees was obviously higher than that of the contrast, and was increasing gradually while the water stress was intensifying, the leaf temperature was especially high all through under serious water stress; (2) The daily variation of the leaf temperature of the three types of trees under water stress was matching the variation of the air temperature during the day, both of which were lower during morning and night, meanwhile higher during noon. In addition, except that the leaf temperature ofQuercusvariabiliwas higher than air temperature under serious water stress between 10:00 and 16:00, the curve of the daily variation of leaf temperature ofPlatycladusorientalis,RobiniapseudoacaciaandQuercusvariabiliwas lower than that of the air temperature under any other water stress level; (3)The variation of leaf temperature followed that of the air temperature, howere it showed variousiy on different trees.PlatycladusorientalisandQuercusvariabilioffered unimodal type curves, whileRobiniapseudoacaciaoffered a bimodal pattern.
Platycladusorientalis;Robiniapseudoacacia;Quercusvariabili; leaf temperature;water stress
2014-12-11
国家自然科学基金项目(31270750);“十二五”国家科技支撑计划(2011BAD38B00)
杨文卿(1988-),男,河南南阳人,硕士研究生,主要从事植被恢复方面的研究。
赵 勇(1962-),男,河南商丘人,教授.博士研究生导师。
1000-2340(2015)03-0325-06
Q945. 78
A
