小秦岭国家级自然保护区维管植物群落结构与组成
2015-07-01韦博良贾宏汝韩军旺叶永忠袁志良
韦博良,陈 云,许 宁,贾宏汝,韩军旺,叶永忠,袁志良
(1.河南农业大学生命科学学院, 河南 郑州 450002;2.郑州大学离子束生物工程省重点实验室,河南 郑州 450052;3.河南小秦岭国家级自然保护区管理局, 河南 灵宝 472500)
小秦岭国家级自然保护区维管植物群落结构与组成
韦博良1,陈 云1,许 宁1,贾宏汝2,韩军旺3,叶永忠1,袁志良1
(1.河南农业大学生命科学学院, 河南 郑州 450002;2.郑州大学离子束生物工程省重点实验室,河南 郑州 450052;3.河南小秦岭国家级自然保护区管理局, 河南 灵宝 472500)
沿海拔梯度设置56个样地,对小秦岭植物群落组成与结构进行调查分析。结果表明,(1)小秦岭群落物种丰富,样地中共调查到有143科330属522个类群;(2)通过对乔木,灌木,草本的重要值分析可知,华山松、连翘、莎草占优势;(3)对乔木层进行径级结构分析,葛萝槭和领春木呈倒“J”形,说明森林更新良好。华山松、秦岭冷杉等呈正态分布和多峰特征,说明面临着后代更新困难的局面,属于衰退种群;(4)对乔木层的植物进行树高和胸径的拟合作图,所建立的二元分布模型可以为研究树木胸径、树高的实际关系和动态提供信息。
海拔梯度;群落结构;径级结构;多度分布;拟合优度
植物群落结构可以揭示物种生态适应性成因,有助于生态保护。植物群落的组成与结构是植物群落生态学研究的基础,植物群落主要受控于群落物种的生态、生物学特性及它们的特殊配置方式。研究群落结构有利于理解群落的物种组成特点,功能特征,物种与环境之间的关系等,是制定植物群落多样性保护与管理措施的重要理论参考依据[1]。研究群落结构和物种组成对阐明种群生态特性、更新对策乃至群落的形成及其稳定性与演替规律等都具有重要意义[2,3]。植被数量生态学中重要值(Important Value, IV)是由CURTIS等[4]1951年首先提出来的,它是反映某个物种在森林群落中作用和地位的综合数量指标。重要值作为一种综合性指标,在衡量某种植物于群落中相对重要性的同时也指出这种植物分布的最适生境。重要值是反映种群在群落中的地位和作用的相对数量指标。特别在种类繁多,优势种不明显的群落中,重要值分析显得尤为重要。本研究通过对不同功能群植物重要值变化的研究,旨在为进一步研究生物多样性提供理论依据。胸径、树高是森林调查和经营中最重要的测量因子[5],是常用来计算立木材积、立地指数、森林生长和收获、演替和生物量等相关的重要变量[6,7]。在植物生态研究中,群落年龄结构能近似地表示为径级大小,它综合反映了植物数量的变化趋势及与环境间的相互关系,能较好地解释种群动态变化,因而得到广泛的应用[8,9]。种群的径级结构不仅可以反映种群动态及其发展趋势,并可在一定程度上反映种群与环境间的相互关系以及种群在群落中的作用和地位。但由于树木胸径测定简单、方便、准确,而树高测量费时费力,还会产生很大的测量误差,因此在目前的森林调查和试验研究中,常常只测量部分树木的树高,缺失的树高则通过不同的树种的树高——胸径模型来预测[7,10]。研究和探讨树高与胸径的相关关系,是揭示森林成分和生长规律的基础[11,12]。目前,通过胸径和树高等生长指标之间的相关关系建立数学模型来估算森林植被生产力的方法,得到了普遍应用。小秦岭自然保护区已有的研究工作主要基于植被带的划分、植物资源考察、植物区系分析等方面开展,而运用定量的数学分析手段对小秦岭森林群落进行深入、细致的研究还很少。因而,本研究从群落外貌、群落结构、群落的数量特征、乔木层树高和胸径的拟合分析这4个方面对小秦岭森林群落植物多样性和群落结构进行研究,从而加深对本区植物群落结构、性质的认识,以期为本区生物多样性的保护和持续利用提供参考或依据。
1 研究区概况与研究方法
1.1 研究区概况
小秦岭横跨陕西与河南2省,为华山东延部分,止于灵宝市区南弘农涧两岸。小秦岭国家级自然保护区位于豫陕2省交界的灵宝市西部、小秦岭北麓,属于暖温带大陆性季风型半干旱气侯。地理坐标为北纬34°23′~34°31′,东经110°23′~110°44′,属森林生态类型自然保护区。小秦岭保护区是我国生物多样性保存最好的区域之一,森林覆盖率高达81.2 %[13],保护区内共有维管束植物137科708属1 958种。该地区年平均气温在11.2~14.2 ℃[14],年降水量505.0~719.5 mm[15,16],海拔大多在1 200~2 000 m[17],主峰老鸦岔垴高达2 413.8 m[18],为河南最高峰。
1.2 样地的设置与调查
本研究依据研究区域内物种的生物学特性以及生活史特征,将群落内的维管植物分为3个层次:乔木层(树高≧5 m)、灌木层(0.5 m≦树高<5 m)和草本层(0 m≦树高<0.5 m)。在保护区内选择具有代表性的山体,从海拔1 020 m开始,直至海拔2 413.8 m,沿海拔梯度选择具代表性的群落设置样地,海拔每升高50 m设置2个20 m×20 m的乔木样方,在每个乔木样方中随机选择典型群落设置1个5 m×5 m 的灌木样方,并在每个乔木样方的4个角分别设置1个1 m×1 m 的草本样方,共设置56套样方。其中乔木样方调查目标为树高≧5 m的乔木层个体,这些个体主要为成年的乔木;灌木样方调查目标为0.5 m≦树高<5 m的灌木层个体,这些个体主要为成年的灌木以及乔木的幼树;草本样方调查目标为树高<0.5 m的草本层个体,这些个体主要为草本以及灌木和乔木的幼苗。调查记录内容包括:物种的生物学特征如:盖度、高度、胸径,多度等,以及群落综合特征如:海拔、坡度、坡向、经纬度等。
1.3 数据分析方法
依据基本调查数据,分别计算各样地内不同高度层出现的植物种类的重要值,并对其进行排序。目前应用较多的是经CURTIS等修改的重要值指数,是相对密度或相对多度、相对频度和相对显著度3项指数的综合。本研究物种重要值依下式计算[19]:
乔木重要值=(相对密度+相对频度+相对优度)/ 3
灌木重要值=(相对密度+相对频度+相对盖度)/ 3
草本重要值=(相对多度+相对盖度)/ 2
其中,相对多度是指某植物种的个体数所占群落种类总个体数的百分率。其计算公式为:相对多度=(某种的多度/所有种的多度之和)×100%。相对频度是某种植物出现的次数与样方总数的百分比求得的,其计算公式为:相对频度=(某种的频度/所有种的频度之和)×100 %。相对盖度,即相对显著度,是指单位面积中某种植物显著度与全部树种显著度的百分比求得的,其计算公式为:相对显著度=(某树种显著度/全部树种显著度之和)×100 %。
2 结果与分析
2.1 物种组成
植物种类组成是植物群落最重要的特征之一,是决定群落性质和鉴别不同群落类型的主要因素[20,21]。据此次调查统计,样方内共有维管植物6 686株,隶属于143科330属522种。样地内乔木类群比较多的是松科(Pinaceae)、漆树科(Anacardiaceae)、杨柳科(Saliaceae)、壳斗科(Fagaceae)和槭树科(Aceraceae);灌木比较多的是木犀科(Oleaceae)、蔷薇科(Rosaceae)和卫矛科(Celastraceae);草本比较多的科是莎草科(Cyperaceae)、禾本科(Gramineae)、菊科(Compositaesae)和毛茛科(Ranunculaceae)。
2.2 优势度
优势度表示植物群落内各植物种类处于何种优势或劣势状态的群落测定度。重要值的数值大小可以作为群落中植物种优势度的一个度量标志[22],重要值也可以体现群落中每种植物的相对重要性及植物的适宜生境。
2.2.1 乔木层 乔木层的优势树种不仅影响群落的外貌和结构,而且最能客观地体现该种植物对该地生境的适应度,通常以其重要值的大小顺序来确定[23,24]。样地内乔木层植物共126种。
从物种多度来看(表1),华山松个体数最多,其相对多度为12.71 %;其他个体数超过100的类群按个体数多少排序依次为领春木、东陵山柳、漆树、葛萝槭、油松和糙皮桦,这6个树种的个体数之和占乔木层总个体数的24.71 %。而蒿苹四蕊槭、君迁子、华北卫矛和金花忍冬等18个物种在乔木层中均仅有1株。从平均胸径可以发现(表1),样地内所有乔木层个体的平均胸径为9.79 cm。其中,水曲柳平均胸径最大,达25.88 cm,其余较大的是糙皮桦、秦岭冷杉、槲栎和蒙古栎等。样地内乔木层个体的总胸高断面积为59.44 m2·hm-2。其中胸高断面积大于1 m2·hm-2的树种不到20种,从高到低依次为华山松、糙皮桦、秦岭冷杉、槲栎、油松等。这几个树种的胸高断面积之和占总胸高断面积的79.19 %(表1),由此可见它们的优势性。
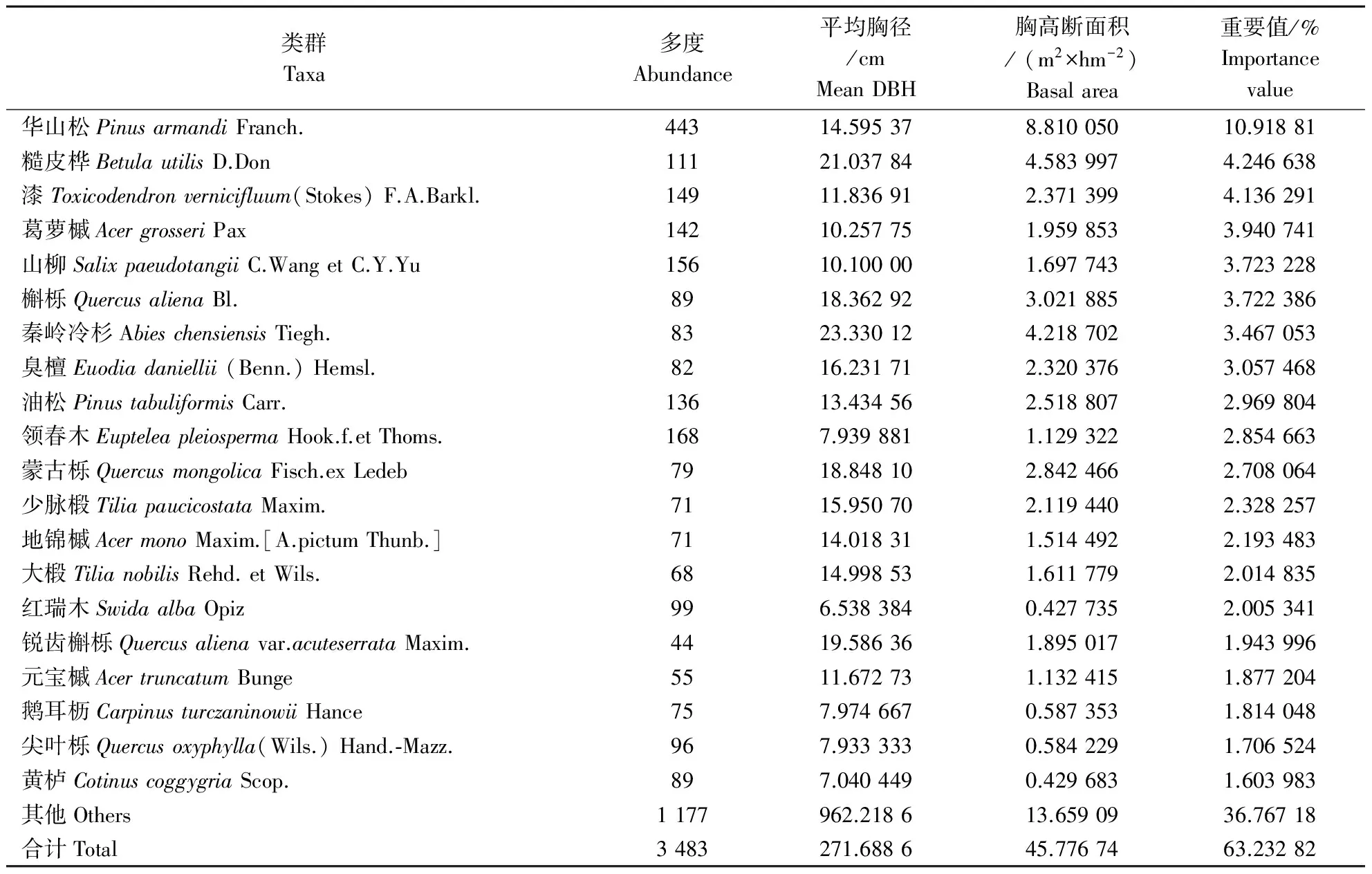
表1 乔木层植物重要值
从物种的重要值来看(表1),华山松的重要值最大为10.92 %,既是个体数最多又是胸高断面积最大,在乔木层中占有绝对优势。重要值大于2 %的树种有15种,这些树种的多度和胸高断面积分别占乔木层物种总多度和总胸高断面积55.90 %和69.23 %。
综上所述:从物种多度、平均胸径和胸高断面积这3个指标可以了解到:一些树种既有较多的个体,也有较大的平均胸径,因此胸高断面积也大,这些树种就是乔木层中的优势种,如华山松、糙皮桦等;有一些物种个体数很多,但平均胸径小,胸高断面积也小,这些种是乔木下层的优势种,如葛萝槭、东陵山柳等;还有一些平均胸径大,但个体数很少,胸高断面积小,这些属于群落中的偶见种,如水曲柳、红桦等。
2.2.2 灌木层 从物种多度来看(表2),连翘的个体数最多,共有103个,占到总个体数的6.89 %。个体数超50的类群依次为华北绣线菊、花楸树、山梅花和栓翅卫矛等,这4个树种的个体数之和占所有物种总个体数的28.36%。而白皮松等32个物种只有1个个体。
从物种的重要值来看(表2),连翘的重要值最大为6.20%。既是个体数最多又是重要值最大的树种,在灌木层中占有优势。重要值大于2 %的树种有9种,这些树种的多度占灌木层物种总多度的40.80 %。

表2 灌木层和草本层物种的重要值
2.2.3 草本层 从物种多度来看(表2),莎草的个体数最多,为2 357个,占到总个体数的15.46 %,个体数超500的其他物种大小顺序依次为宽叶薹草、活血丹、求米草、早熟禾等,这4个草本的个体数之和占草本层总个体数的20.21 %。而卷叶黄精等25个物种只有1个个体。
从物种的重要值来看(表2),莎草的重要值最大为7.32 %,既是个体数最多又是重要值最大,在草本植物中占有优势。重要值大于2 %的树种有11种,这些树种的多度占乔木层物种总多度的44.83%。
2.3 径级结构与更新
径级结构是指林木株数按径级大小的分配状况,是植物群落稳定型和生长发育状况的重要指标[25,26]。径级结构不仅对群落结构有直接影响,而且能客观地体现群落的发展趋势。因此,研究森林种群径级结构对阐明种群生态特性、更新对策乃至群落的形成及演替规律等都具有重要意义。
对小秦岭乔木层重要值前9种物种的径级分布研究(图1)发现,葛萝槭、领春木的径级分布基本上呈倒“J”型,即小径级的个体数量居多,随着胸径增大,株数减少,个体径级分布连续。表明这些森林类型的天然更新状况良好,物种的幼苗更新能力较强,属于进展型种群。华山松、糙皮桦、漆树、油松、槲栎的径级结构近似于正态分布,为乔木上层种,个体主要集中在中径级,大径级与小径级个数则相对较少,反映出样地中的这5个优势种的种配置不均匀,面临着后代更新困难的局面,属于衰退种群。臭檀的径级分布没有明显的形状分布,个体主要集中在小径级和中径级而且个体数比较多,介于衰退型种群和进展型种群之间。秦岭冷杉径级分布呈现多峰特征,它的胸径主要以中径级居多,各径级分布极不合理,径级缺失严重,人为的干扰破坏活动大,大径级的被采伐,干形不好、有病虫害的、天灾致畸形的残留下来,再加上气候的变化等因素的影响,其数量急剧减少,目前正处于极度濒危状态,属于衰退型种群。
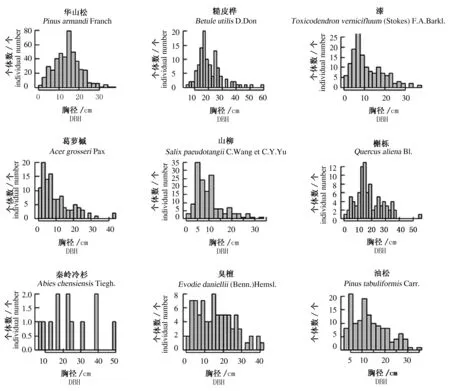
图1 乔木层重要值前9位物种的径级结构
2.4 乔木层树高和胸径的拟合
本研究以小秦岭重要值前10位的乔木层植物中的9种为研究对象,评价9个常见树高-胸径模型,作树高与胸径分布散点图,利用R语言分析软件,以个体树高为y,个体胸径为x,选择合适的Linear(线性)曲线模型进行直线拟合。利用R语言分析软件,可以得出散点图的截距、斜率、相关系数、以及显著性,结果如图2所示。

图2 乔木层重要值前9位物种的树高与胸径分布散点图
此9种乔木的胸径和树高拟合方程分别为:
华山松:y=0.503 27x+1.984 65
糙皮桦:y=0.176 02x+5.990 69
漆树:y=0.317 84x+5.152 15
葛萝槭:y=0.330 24x+3.849 42
山柳:y=0.278 07x+2.848 62
槲栎:y=0.297 83x+5.041 15
秦岭冷杉:y=0.494 7x-0.556 8
臭檀:y=0.350 95x+5.029 82
油松:y=0.275 17x+6.095 81
由方程可知:华山松、秦岭冷杉的斜率很大,但它的截距很小;糙皮桦、油松的斜率较小,但它的截距比较大;东陵山柳、葛萝槭的斜率较小,它的截距也较小;漆树、臭檀、槲栎的斜率较大,它的截距也较大。其中,华山松的斜率最大,但它的截距最小。
通过分析比较拟合方程的R2,检验方程的线性显著性水平。其中,R2等于误差平方和除以总离差平方和的商;误差平方和等于观测值与拟合值的差的平方的和;总离差平方和等于观测值与该观测数据的样本均值的差的平方的和。R2值越大,相应回归方程的拟合效果越好,观测数据与理论模型的偏离程度越小;由表可知:相关系数由大到小为臭檀、葛萝槭、槲栎和糙皮桦。相关系数越大说明拟合程度越好,可以用胸径较准确地来预测树高。本研究中乔木层重要值前9种的物种的拟合都极为显著。
3 结论与讨论
乔木层群落中,葛萝槭种群径级结构呈倒“J”形的增长型,幼苗个体数占有绝对优势,个体径级分布连续,说明其具有前期增长,后期稳定的特点,能够进行天然更新。油松、华山松在群落中表现为正态分布的衰退型种,目前处于成熟而渐现衰退的时期。这些在群落演替过程中发挥了重要作用,尽管后备资源不足,但并不意味着它们很快就会被其他优势种群所取代。秦岭冷杉的胸径主要以小径级居多,各径级分布极不合理,径级缺失严重,由于人为和气候等因素,目前正处于极度濒危状态。
灌木层群落内的优势种为连翘、花楸树和华北绣线菊。它们的共同特点是稍耐荫,抗寒,抗旱,喜温暖湿润的气候和深厚肥沃的土壤。萌蘖力和萌芽力均比较强。从物种多度这一指标可知:多度个数基本上涵盖每个数量段,总体来说很均衡。一般地,多度个数多的重要值也大,多度个数少的重要值就小。草本层群落内的优势种为莎草、宽叶薹草、活血丹。它们都是多年生草本植物,且其共同特点为多匍匐枝茎,枝茎上多不定根,蔓延能力强,能很好地适应环境。
研究发现,在小秦岭自然保护区,乔木层的主要物种的胸径和树高有很强的相关关系,胸径和树高拟合后得到的相关系数(R2)除了糙皮桦都相对较高。所有模型的方差检验都在0.01的水平上显示为极显著。这为我们以后研究提供了良好的基础。但影响树高的因素很多,如树种之间的竞争、温度、坡度、湿度、人为干扰等,这些都还需要进一步的研究。
本研究通过对小秦岭自然保护区植物群落的结构和组成的调查分析,确定了乔木层,灌木层以及草本层的优势物种,揭示了不同高度层内主要物种的生长趋势,并对乔木层的主要物种树高胸径进行了拟合,这为我们科学管理森林、维持群落稳定以及保护生物多样性等方面了提供理论依据。
[1] 傅志军,张行勇,刘顺义,等.秦岭植物区系和植被研究概述[J].西北植物学报,1996,16(5): 93-106.
[2] SOUL M E.Conservation biology: the science of scarcity and diversity[M].Sunderland: Sinauer Associates Inc,1986: 1-598.
[3] 白雪娇,李步杭,张 健,等.长白山阔叶红松林灌术物种组成、结构和空间分布[J].应用生态学报,2010,21 (8): 1899-1906.
[4] CURTIS J T,MCINTOSH R P.An upland forest continuum in the prairie-forest border region of Wisconsin[J].Ecology,1951,32(3): 476-496.
[5] LEI X,PENG C,WANG H,et al.Individual height-diameter models for young black spruce (Piceamariana) and jack pine (Pinusbanksiana) plantations in New Brunswick,Canada[J].The Forestry Chronicle,2009,85(1): 43-56.
[6] CASTEDO DORADO F,DI GUE-ARANDA U,BARRIO ANTA M,et al.A generalized height-diameter model including random components for radiata pine plantations in northwestern Spain[J].Forest ecology and management,2006,229(1): 202-213.
[7] SHARMA M,PARTON J.Height diameter equations for boreal tree species in Ontario using a mixed-effects modeling approach[J].Forest Ecology and Management,2007,249(3): 187-198.
[8] 田世广,张金屯,张钦弟,等.庞泉沟地区山地杨桦林优势种种群格局与动态[J].北京师范大学学报: 自然科学版,2011,47(2): 175-180.
[9] 臧润国,杨彦承,蒋有绪.海南岛霸王岭热带山地雨林群落结构及树种多样性特征的研究[J].植物生态学报,2001,25(3): 270-275.
[10]COLBERT K C,LARSEN D R,LOOTENS J R.Height-diameter equations for thirteen midwestern bottomland hardwood species[J].Northern Journal of Applied Forestry,2002,19(4): 171-176.
[11]CURTIS R O.Height-diameter and height-diameter-age equations for second-growth Douglas-fir[J].Forest Science,1967,13(4): 365-375.
[12]HUANG S,TITUS S J,WIENS D P.Comparison of nonlinear height-diameter functions for major Alberta tree species[J].Canadian Journal of Forest Research,1992,22(9): 1297-1304.
[13]王海亮,姚冠忠,韩军旺,等.河南小秦岭自然保护区绣线菊属植物资源及栽培利用研究[J].南阳师范学院学报,2006,5(6): 61-64.
[14]袁志良,叶永忠,贾宏汝.河南小秦岭自然保护区蕨类植物区系研究[J].西北植物学报,2006,26(8): 1677-1681.
[15]李冬林,王宝松,阮宏松,等.河南小秦岭自然保护区生态评价[J].安徽农业大学学报,2007,34(1): 97-102.
[16]王海亮,李冬林,韩军旺,等.河南小秦岭自然保护区槭属植物的分布与区系分析[J].中国野生植物资源,2006,25(4): 47-50.
[17]黄 栋,王 谨,李冬林,等.河南小秦岭忍冬属植物的分布及资源利用探讨[J].中国野生植物资源,2010,29(1): 28-31.
[18]王海亮,李冬林.河南小秦岭卫矛属植物资源的分布与开发利用研究[J].河北林果研究,2010,25(4): 348-351.
[19]王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报: 自然科学版,2010,33(2): 312-316.
[20]胡正华,于明坚,余志良,等.古田山国家级自然保护区甜槠群落物种多样性研究[J].中国生态农业学报,2005,13(1): 35-37.
[21]胡正华,于明坚,丁炳扬,等.古田山国家级自然保护区常绿阔叶林类型及其群落物种多样性研究[J].应用与环境生物学报,2003,9(4): 341-345.
[22]宋永昌.植被生态学[M].上海: 华东师范大学出版社,2001: 127-134.
[23]刘克旺,石道良,杨旭红.湖南绥宁县神坡山穗花杉群落特征初步研究[J].武汉植物学研究,1999,17(2): 137-145.
[24]张金屯.植被数量生态学方法[M].北京: 中国科学技术出版社,1995.
[25]叶万辉,曹洪麟,黄忠良,等.鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J].植物生态学报,2008,32(2): 274-286.
[26]孟宪宇,张 弘.闽北杉木人工林单木模型[J].北京林业大学学报,1996,18(2): 1-8.
(责任编辑:朱秀英)
Plant community structure and composition of the Xiaoqinling National Nature Reserve
WEI Boliang1, CHEN Yun1, XU Ning1, JIA Hongru2, HAN Junwang3, YE Yongzhong1, YUAN Zhiliang1
(1.College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China; 2.Henan Provincial Key Laboratory of Ion Beam Bio-engineering, Zhengzhou University, Zhengzhou 450052, China; 3.Administration Bureau of Xiaoqinling National Nature Reserve of Henan, Lingbao 472500, China)
56 sample plots along the altitudinal gradient were set to investigate and analyze the composition and structure of the Xiaoqinling plant community. The results showed that: (1) There are abundant community species in plots, belonging to 143 families, 330 genera and 522 taxa; (2) By analyzing the important value of trees, shrubs and herbaceous we find thatPinusarmandi,ForsythiaandSedgesare in the dominant position; (3) The analysis of the size structure of tree layer suggests the following results:PizmapleandEupteleapresent the reversed “J” shape, which showed good forest regeneration; On the contrary, Pinusarmandi,Abieschensiensisand others showed normally distributed and multi-modal features, indicating that these future generations are facing a difficult update situation, belonging to the decline populations; (4) By building a bivariate distribution model based on the analysis of the fitting of tree height and DBH, we can provide information for the study of actual relationship and dynamic between tree height and DBH.
elevatio gradient; community structure; size structure; abundance distribution; goodness of fit
2014-10-19
河南省科技攻关项目(132102110133);河南省教育厅科学技术研究重点项目(14A180013)
韦博良(1991-),男,河南平顶山人,硕士研究生,主要从事森林生态方面的研究。
叶永忠(1957-),男,湖北黄冈人,教授,博士生导师。
1000-2340(2015)03-0335-08
S718
A
