侧柏-山毛桃混交林枯落物分解动态与养分归还的研究
2015-07-01赵佳宝孔玉华张盼盼杨喜田常海荣王怡博
吴 明, 赵佳宝, 孔玉华, 张盼盼, 杨喜田, 常海荣, 王怡博
(河南农业大学林学院,河南 郑州 450002)
侧柏-山毛桃混交林枯落物分解动态与养分归还的研究
吴 明, 赵佳宝, 孔玉华, 张盼盼, 杨喜田, 常海荣, 王怡博
(河南农业大学林学院,河南 郑州 450002)
以黄河小浪底库区内10 a生的侧柏、山毛桃纯林及侧柏-山毛桃混交林为研究对象,采用枯落物分解袋法对其枯落物的季节分解动态与养分归还进行分析。结果表明:(1)在气温高、降水多的5~11月,3种林分下枯落物的失重率变化较大,在气温低、降水较少的11月到次年3月,枯落物失重率变化较为平缓。不同林分间枯落物的分解速率大小顺序为:山毛桃纯林>侧柏-山毛桃混交林>侧柏纯林;(2)枯落物养分含量的变化趋势呈现出:N含量先升后降,为富集-释放的模式;P含量持续升高,出现富集现象;K含量不断下降,为直接释放的模式;(3)N、P、K在3种林分中的年归还量大小顺序分别为:侧柏-山毛桃混交林>山毛桃纯林>侧柏纯林;侧柏-山毛桃混交林>侧柏纯林>山毛桃纯林;山毛桃纯林>侧柏-山毛桃混交林>侧柏纯林。相对于纯林,侧柏-山毛桃混交林有利于提高枯落物的分解速率,显著增加N和P的归还量(P<0.05)。
侧柏;山毛桃;枯落物分解;养分归还
森林枯落物是森林生态系统的重要组成部分,是林木生长发育过程中的新陈代谢产物[1],在保持正常的养分循环及平衡、涵养水源和水土保持等方面起着重要的作用[2,3]。在陆地生态系统中,90%以上的地上部分净生产量通过枯落物的方式返回地表[4]。枯落物分解是森林生态系统中养分归还的主要途径,枯落物通过分解向土壤释放的养分元素是林木维持自身生长所需养分的主要来源之一,对土壤肥力的改善和森林生产力的提高具有十分重要的作用[5,6]。以往研究枯落物分解时往往只针对单一树种,而关于混合枯落物的研究较少[7]。有报道表明,较单一树种,其混交林的生物生产力与木材产量均表现出增加,如杨树(Populusalba)与刺槐(Robiniapseudoacacia)混交林[8]、白桦(Betulaplatyphlla)与山杨(Populusdavidiana)混交林[9]、杉木(Cunnighamialanceolata)和南酸枣(Choerospondiasaxillaris)混交林[10]等。关于生产力增加的机制,可能是一种枯落物与另一种枯落物混合分解时,分解速率与养分释放速率得到加强的结果[8-13]。侧柏(Platycladusorientalis)被广泛应用于北方干旱山地的植被恢复,喜光,较耐寒,抗风力较差;耐干旱,耐贫瘠,生长缓慢,寿命极长。山毛桃(Prunusdavidiana)为落叶小乔木或灌木,其根系发达,适应性强,耐旱,耐寒,是黄土丘陵和北方石质山区自然分布的物种,也是植被恢复的优良灌木树种[14]。本研究选择处于植被演替初期阶段的10 a生侧柏纯林、山毛桃纯林及侧柏-山毛桃混交林为研究对象,探讨不同枯落物组成对枯落物分解速率、养分归还速率等影响,为混交树种的合理搭配提供理论依据,从而为营建强健的森林生态系统以及保障生态功能的发挥提供依据。
1 研究地概况
试验区位于河南省济源市黄河小浪底库区(34°57′N、112°20′E),海拔在350~700 m之间,该区属于暖温带大陆性季风气候,年平均气温13.1 ℃,7月平均气温26.2 ℃。1月平均气温-0.5 ℃,极端气温为﹣17.2 ℃和43.7 ℃。全年无霜期平均215 d。年降雨量一般在600~800 mm,60%左右的降水量集中在7-9月。年平均蒸发量1 800~2 100 mm。年平均相对湿度约为60%。
试验区地形破碎,主要由低山和丘陵等地貌类型组成。土壤结构不良,石砾较多,养分含量低,且多属粗骨土,易侵蚀,土层浅薄。其区内植被类型丰富,乔木群落主要由落叶阔叶林组成,多数为人工次生林,灌木群落主要有酸枣和荆条。
2 试验设计与方法
2.1 试验设计
试验地位于南坡中上坡位,坡度约为30°。本研究选取的树种侧柏和山毛桃均为10 a生,株行距50 cm,密度约为6万株·hm-2,混交林中侧柏与山毛桃混交比为1∶4。 侧柏为常绿针叶树种,树高分布在6~9 m,胸径为4~9 cm,树型直立,生长发育比较缓慢,林下枯落物较少;山毛桃为落叶阔叶树种,树高分布在3~7 m,胸径为7~11 cm,其树杈较多,树枝分散,生长发育较快,林下枯落物较多。
2013-05-01在小浪底库区试验基地,选取立地条件一致(坡向、海拔、坡位、坡度)的侧柏、山毛桃纯林及其侧柏-山毛桃混交林(简称混合)中,分别设置1 m×1 m的小样方18个,共54个。收集地面新凋落和半分解的枯落物,装入自封袋内带回实验室,并采取样方内表层0~20 cm的土壤,混合均匀,以备装盆用,为枯落物分解提供微生物菌源。
分别按林分称取50 g(烘干重)枯落物装入尼龙网分解袋中(网眼为1 mm,规格为30 cm×30 cm),每林分18个分解袋。其中,侧柏-山毛桃混合枯落物中侧柏和山毛桃枯落物的比例是1∶4。2013-05-02,将采取的表层土壤装入盆钵内(口径32 cm、 深度14 cm),装土深度为12 cm,并将分解袋分别置于相应盆钵中的土壤表层,使枯落物直接与土壤接触并将盆钵随机放置于相应林分中。
2.2 采样与分析
于2013-05-02、2013-07-02、2013-09-02、2013-11-02、2014-01-02、2014-03-02,经历春、夏、秋、冬4个季节,分别取侧柏、山毛桃、侧柏-山毛桃混交林枯落物各3袋带回实验室,清除分解袋表层泥土,用去离子水冲洗干净,置于80 ℃恒温条件下烘干至恒重,计算其失重率和分解速率,粉碎并过60目的筛之后供化学分析。植株样品全氮采用半微量凯氏定氮法测定(全自动凯氏定氮仪),全磷采用H2S04-H2O2消煮钼锑抗比色法测定(分光光度计),全钾采用H2SO4-H2O2消煮火焰光度计法(分光光度计)[16]。
枯落物养分归还速率由分解前叶枯落物中养分元素浓度及其重量(50g),求得分解袋内不同林分枯落物分解前的养分数量,再由最后1次取样时测得的枯落物养分元素浓度及其残留量求得分解末期3种林分枯落物中养分元素的残留量。最后由二者之差求得在整个分解过程中各林分枯落物的养分归还速率[17]。
2.3 指数衰减模型的建立
依据OLSON[15]提出的指数衰减模型(公式1)进行模拟。如定义枯落物分解50%事所需时间为分解半衰期t0.5,分解95%时所需时间为t0.95,根据所测各林分中枯落物的失重率,应用该模型对各林分枯落物的分解参数进行估测。
Xt/X0=e-kt
(1)
式中:t为X0与Xt之间的时间间隔,d;X0为枯落物的初始干重,g;Xt为t时刻的枯落物残留干物质重,g;k为枯落物的分解速率常数,d-1。
2.4 数据处理
采用SPSS 21.0 软件进行数据分析,运用Microsoft Excel 软件制作图表。
3 结果与分析
3.1 枯落物分解速率的季节性动态变化
枯落物在分解的过程中,通常用枯落物的失重率来表示质量的变化。由图1可以看出,3种枯落物的累积失重率的变换范围为12.4%~47.7%,均呈现上升的趋势,其中以山毛桃的为最高。其中,5至7月,侧柏、山毛桃及混合枯落物的月平均失重率分别为:6.2%、11.3%及10%,变化较大;7月至次年1月,侧柏和山毛桃月平均失重率变化较平缓,分别为3.6%、3.9%;次年1月,混合枯落物的累积失重率为36.7%,较11月的38.3%下降了1.6%;次年1~3月,侧柏、山毛桃及混合枯落物平均月失重率变化大小分别为:6.1%、0.8%及1.4%。这一变化可能是由于5~11月气温较高,降水较多,致使微生物数量增多、活性加强,有利于枯落物的分解;另一方面,5~7月,枯落物中易分解成分的含量相对高一点,平均月失重率变化较为明显。11月至次年3月,由于试验地出现雨雪天气,导致枯落物分解较为平缓,尤其是混合枯落物的分解基本处于停滞状态。
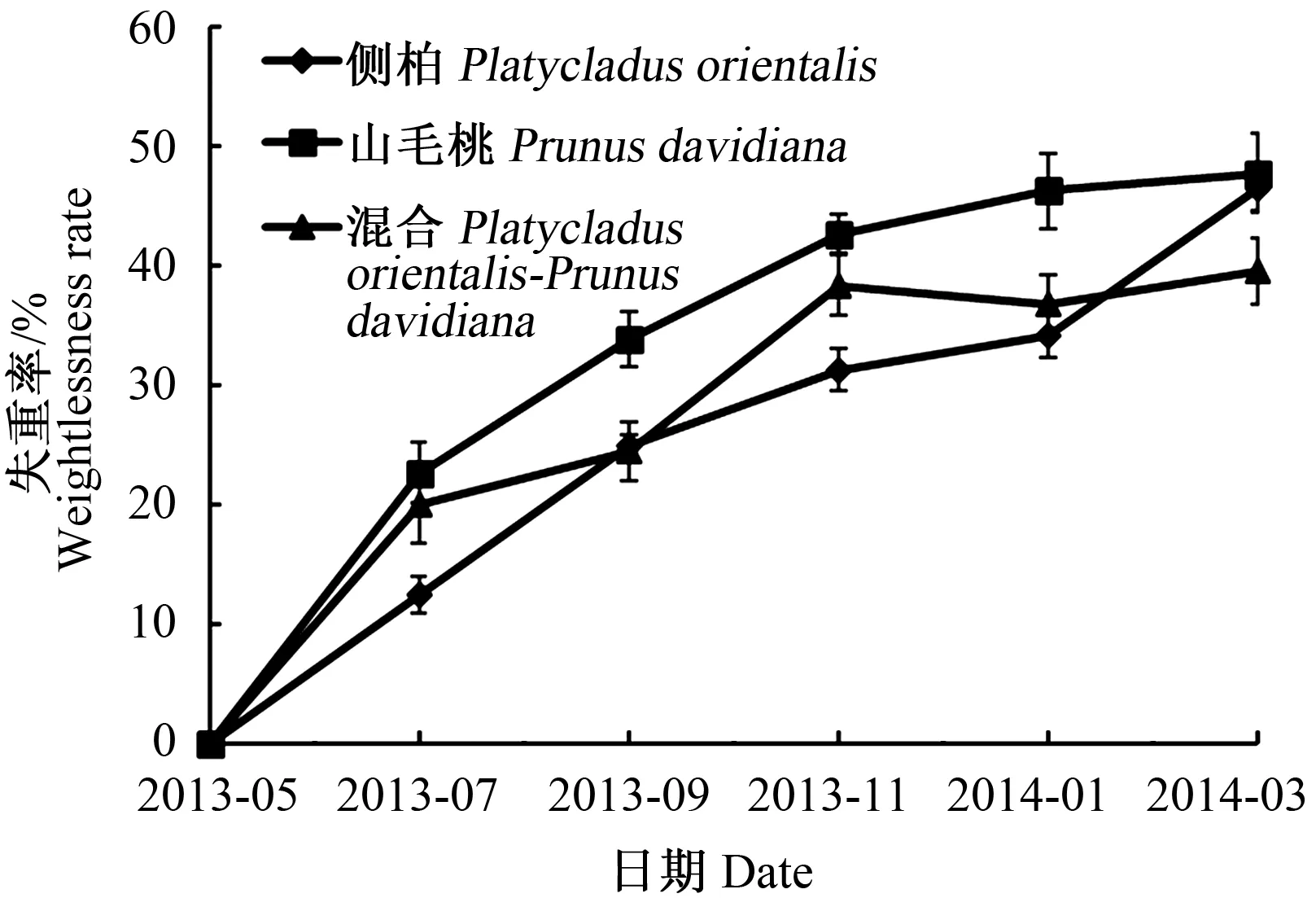
图1 3种林分枯落物分解过程中的失重率变化
由表1可以看出,3种林分枯落物的分解速率大小为:山毛桃纯林>侧柏纯林>侧柏-山毛桃混交林。3种林分枯落物分解50%所需时间分别为114、83、107 d;分解95%需要时间分别为2.4 、1.7、2.2 a。由此可知,山毛桃枯落物的分解速率最大,需要分解的时间最短;侧柏枯落物的分解速率最小,需要分解的时间最长;混合枯落物的分解速率以及需要分解的时间处于中间值。影响枯落物分解速率的因素主要有森林生态系统类型、枯落物组成和环境条件等,而枯落物碳氮比及木质素含量在分解过程中意义重大[18]。另外,枯落物形态可能也会影响其分解速率。维持枯落物一定的分解速率,即可为植物生长持续提供营养,又可保证适当的土壤表层覆盖,因此混交林对于现实森林经营具有十分重要的价值。
3.2 3种分解残留物中养分含量动态变化
由图2可以看出,在3种林分的枯落物分解过程中,N含量整体上为先升后降的趋势,呈富集-释放的迁移模式。山毛桃枯落物的全N含量的变化幅度最大,侧柏枯落物的全N含量的变化幅度最小,混合枯落物的全N含量的变化幅度则基本介于两者之间。5~11月,侧柏、山毛桃及混合枯落物的全N平均含量的变化范围分别为0.460~0.973、0.560~2.338、0.680~1.876 g·kg-1;山毛桃的变化幅度最大,侧柏的变化幅度最小。其中,枯落物全N平均含量在11月出现最大值,分别为0.973、2.338、1.876 g·kg-1。结果表明,5~11月,N含量出现富集,可能是由于这期间,雨热同期,土壤生物活动频繁,从而导致枯落物中的N元素出现富集现象。从11月开始,N含量大量释放。其中,11月至次年1月,3种林分枯落物的N含量剧烈下降,1~3月,N含量缓慢上升。

表1 3种林分枯落物的Olson模型及分解速率
注:k:分解速率常数;t0.5:50%干物质分解需要的时间(t);t0.95:95%干物质分解需要的时间(t)。
Note:k: constant of decomposition rate;t0.5: time (t) needed for 50% of dry mass decomposed;t0.95: time (t) needed for 95% of dry mass decomposed.

图2 3种林分枯落物分解过程中全N含量变化
在3种林分的枯落物分解过程中,随着时间的延长,P含量呈现升高的趋势,出现富集现象(图3)。山毛桃枯落物的全P变化幅度最大,侧柏枯落物的全P变化幅度最小,混合枯落物的全P含量的变化幅度基本介于两者之间。本次试验中,侧柏、山毛桃及其混合枯落物中全P平均含量变化幅度很小,分别为0.021、0.047 、0.012 g·kg-1。这可能是由于分解过程较短,富集量没有达到阈值,分解率较低所致。
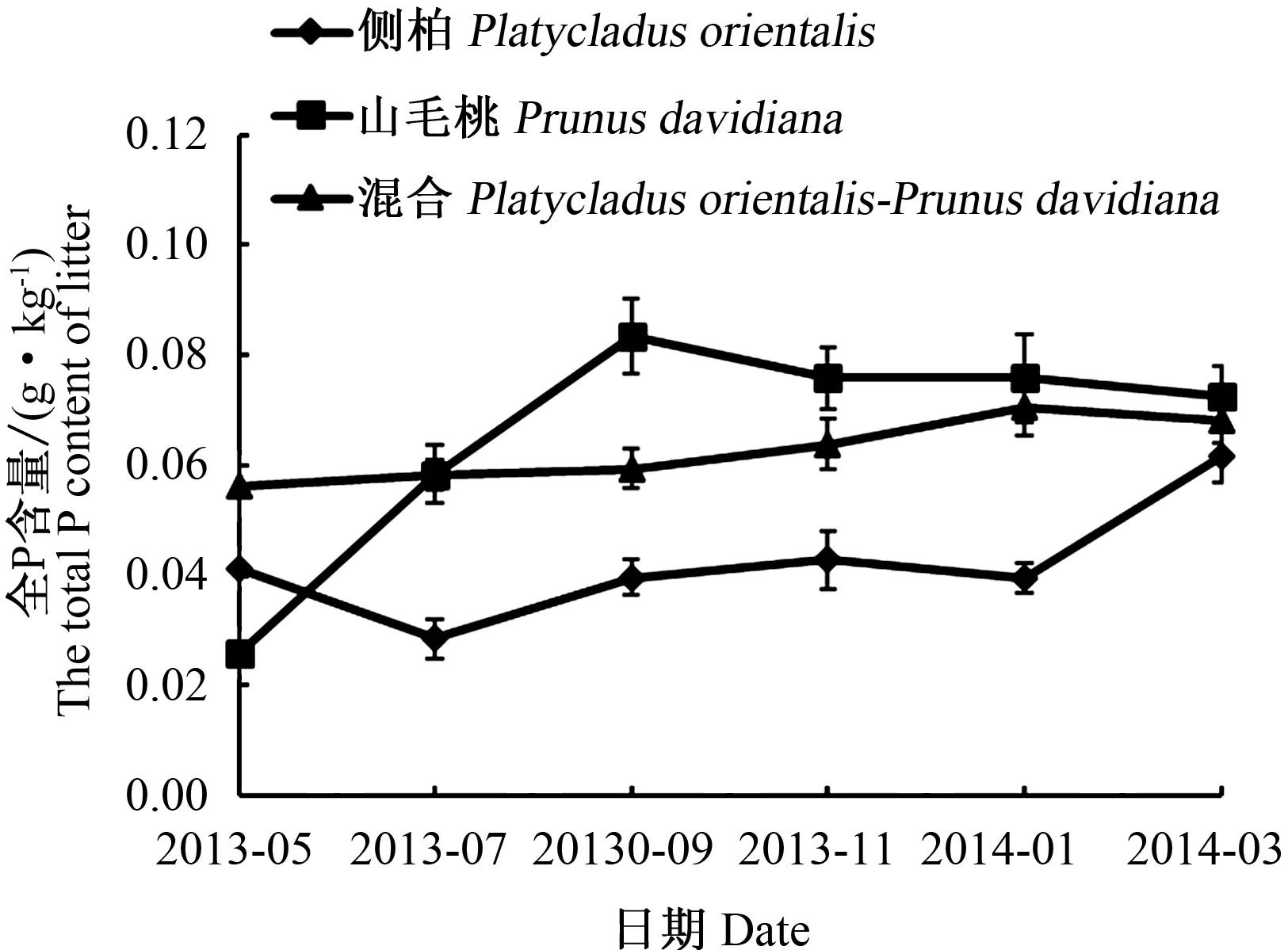
图3 3种枯落物分解过程中全P含量变化
在3种林分的枯落物分解过程中,易淋溶的K元素在分解过程中浓度不断下降,为直接释放的迁移模式,且前期比后期释放的快,山毛桃枯落物的全K变化幅度最大,侧柏枯落物的全K变化幅度最小,混合枯落物的全K含量的变化幅度基本介于两者之间(图4)。5~7月,侧柏、山毛桃及其混合枯落物中全K平均含量的变化范围分别为0.278~0.072、0.550~0.124、0.461~0.139 g·kg-1,山毛桃的变化幅度最大,侧柏的变化幅度最小。3种林分枯落物的全K含量在5月份为最大值,7月份为最小值。7~9月全K含量有明显的上升趋势,9月至次年3月3种枯落物的全K含量变化值较小。这可能是由于5~7月,K元素随着雨水的增多发生大量的淋溶现象,导致3种林分枯落物的K含量剧烈下降,变化幅度较大。

图4 3种林分枯落物分解过程中全K含量变化
3.3 枯落物分解过程中各养分的年归还量
在大多数的生态系统中,林木生长所需要的N和P有69%~87%是由枯落物分解所提供的[18]。由图5可看出,如果排除微生物的影响,各林分枯落物归还养分元素的特点明显不同。其中,P的含量及年归还量较N和K的较少,甚至在山毛桃纯林中,由于P的富集,其归还量出现了负值,为-46.40%,这与N、K的归还量增加的趋势截然不同。在整个分解过程中,3种林分的N的年归还量大小顺序依次为:侧柏-山毛桃混交林>山毛桃纯林>侧柏纯林; P的年归还量大小顺序依次为:侧柏-山毛桃混交林>侧柏纯林>山毛桃纯林;K的年归还量大小顺序依次为:山毛桃纯林>侧柏-山毛桃混交林>侧柏纯林。

图5 3种林分下枯落物分解的各养分元素归还量
4 结论与讨论
1)在气温高、降水多的5到11月,3种枯落物的平均月失重率较大;在气温低、降水较少的11月到次年3月,3种枯落物的平均月失重率变化较为平缓。以往的研究表明,高温可以促进枯落物的分解,低温则抑制枯落物的分解[19,20]。温度和水分在时间尺度上的变化导致的土壤的干湿交替和冻融现象往往会促进枯落物分解,一方面,这2种现象可能加速枯落物的裂解程度[21,22],另一方面,土壤水分和温度状况的交替转变可能刺激微生物的活性[23]。
2)3种林分枯落物的分解速率总体上呈现山毛桃纯林枯落物>侧柏-山毛桃混交林枯落物>侧柏纯林枯落物。侧柏-山毛桃混交林枯落物类型为针、阔混合枯落物,相对于单一类型的枯落物,其物理结构和化学性质发生改变,可能会增加微生物种类的多样性,导致枯落物周围的分解者群落组成的丰富度增多和活性的增强。本次研究中侧柏-山毛桃混交林枯落物的分解速率处于山毛桃纯林和侧柏纯林之间,与以往关于混交林枯落物分解速率方面的研究结果一致[8,24,25]。
3)森林枯落物的分解可以分为3个阶段:粉碎、物理淋溶和有机物的分解代谢过程[26]。在本次研究中,枯落物养分含量的变化趋势呈现出:N含量先升后降,为富集-释放的模式;K含量不断下降,为直接释放的模式;P含量持续升高,出现富集现象,这与其他的研究结论相同[27-31]。
4)N、P、K 3种养分的年归还量大小顺序依次为:侧柏-山毛桃混交林>山毛桃纯林>侧柏纯林;侧柏-山毛桃混交林>侧柏纯林>山毛桃纯林;山毛桃纯林>侧柏-山毛桃混交林>侧柏纯林。混交林枯落物中N和P的年归还量较山毛桃纯林明显增加,而K的年归还量较侧柏纯林明显增加。
[1] 于恩娜,王金贵,初宝顺. 凋落物及其在森林生态中的作用[J]. 现代农业科技,2009(2):286-288.
[2] 覃世赢. 厚荚相思人工幼林生物量与生产力和养分循环的研究[D]. 南宁:广西大学林学院,2006:22-27.
[3] 官丽莉,周国逸,张德强. 鼎湖山南亚热带常绿阔叶林凋落物量20年动态研究[J]. 植物生态学报,2004,28(4):449-456.
[4] LORANGER G, PONGE J F,IMBERT D,et al. Leaf decomposition in two semi evergreen tropical forests:influence of litter quality [J]. Biology and Fertility Soils,2002,35:247-252.
[5] 郭艳娜,霍沁建,袁 玲. 森林土壤肥力概述[J]. 中国农学通报,2004,20(3):143-148.
[6] 张得强,叶万辉,余清发,等. 鼎湖山演替系列中代表性森林凋落物的研究[J].生态学报,2000,20(6):938-944.
[7] 徐振邦,代力民,张扬建,等. 长白山森林生态系统椴树叶分解进程的研究[J].应用生态学报,2001,12(4):501-504.
[8] 张鼎华,翟明普,林 平,等. 杨树刺槐混交林枯落物分解速率的研究[J].中国生态农业学报,2004,12(3):24-26.
[9] 李雪峰,张 岩,牛丽君,等. 长白山白桦(Brtulaplatyphlla)纯林和白桦山杨(Populusdavidina)混交林凋落物的分解[J]. 生态学报,2007,27(5):1782-1789.
[10]罗修宝. 杉木与南酸枣混交林生物量及土壤肥力研究[J].浙江林业科技,2013,33(2):43-46.
[11]BLAIR J M. DECAY R,nitrogen fluxes and decomposer communities of single and mixed species foliar litter[J]. Ecology,1990,71(5):1976-1985.
[12]BRIONES M I,INESON P. Decomposition of eucalyptus leaves in litter mixtures [J]. Soil Biochem,1996,28:1381-1388.
[13]CARLYLE J C ,MALCOLM D C. Nitrogen availability beneath spruce and mixed larch and spruce stands growing on deep peat:I.Net mineralization measured by field and laboratory in cubations [J]. Plant Soil,1986,93:95-113.
[14]杨喜田,张盼盼,耿建伟,等. 春秋季节侧柏-山毛桃群落演替初期的小气候特征[J]. 气象与环境科学,2014,37(2):23-24.
[15]OLSON J S.Energy storage and the balance of producers and decomposers in ecological systems [J].Ecology,1963,44(2):322-331.
[16]鲍士旦. 土壤农化分析 [M]. 北京:中国农业出版社. 2000:17-18.
[17]李淑兰,陈永亮. 不同落叶林林下凋落物的分解与养分归还[J]. 南京林业大学学报,2004,2(5):61-62.
[18]郭剑芬,杨玉盛,陈光水,等. 森林凋落物分解研究进展[J]. 林业科学, 2006,42(4):93-100.
[19]FACELLI J M,PICKETT S T A. Plant litter: its dynamics and effects on plant community structure [J]. Botanical Review,1991,57(1):1-32.
[20]VOGT K A,GRIER C C,VOGT D J. Production,turnover,and nutrient dynamics of above and belowground detritus of world forests [J]. Advances in Ecology Research,1986,15:303-377.
[21]CHAPIN F S,MATSON P A,MOONEY H A. Terrestrial decomposition in principles of terrestrial ecosystem ecology [M]. New York:Business Media Press,2002:151-175.
[22]POLUNIN N V C. The decomposition of emergent macrophytes in fresh water [J]. Advances in Ecological Research,1984,14:115-166.
[23]李 强,周道玮,陈笑莹. 地上枯落物的累积、分解及其在陆地生态系统中的作用[J]. 生态学报,2014,34(14):3812-3813.
[24]肖慈英,黄青春,阮宏华.松、栎纯林及混交林凋落物分解特性研究[J].土壤学报,2002,39(5):763-767.
[25]陈永亮,李淑兰.胡桃楸、落叶松纯林及其混交林下叶凋落物分解与养分归还的比较研究[J].林业科技,2004,29(5):9-12.
[26]WOOD T G. Field investigation on the decomposition of leave of Eucalyptus delegatensis in relation to environmental factors [J]. Pedobiologia,1991,14:343-371.
[27]卜 涛,张水奎,宋新章,等.几个环境因子对凋落物分解的影响[J]. 浙江农林大学学报,2013,30(5):740-742.
[28]莫江明,布 朗,孔国辉,等. 鼎湖山生物圈保护区马尾松林凋落物的分解及其营养动态研究[J].植物生态学报,1996,20(6):534-542.
[29]魏玉莲. 木材腐朽菌在森林生态系统中的功能[J]. 应用生态学报,2004,15(10):1935-1938.
[30]LEE K,KOO C.Enhancement of growth and survival ofPopulusalbaglandulosa cutting inoculated with ectomycorrhizal fungus,Pisolithustintoriusunder fumigated nursery condition [J]. Journal of Korean Forestry Society,1985,70:72-76.
[31]王 瑾,黄建辉.暖温带地区主要树种叶片凋落物分解过程中主要元素释放的比较[J]. 植物生态学报,2001,25(3):375-380.
(责任编辑:蒋国良)
Study on litter decomposition dynamics and nutrient return in the mixed forest ofPlatycladusorientalisandPrunusdavidiana
WU Ming, ZHAO Jiabao, KONG Yuhua, ZHANG Panpan, YANG Xitian, CHANG Hairong, WANG Yibo
(College of Forestry, Henan Agricultural University, Zhengzhou 450002, China)
10-year old ofPlatycladusorientalisandPrunusdavidianamixed forest was selected as the research subject in the Yellow River Xiaolangdi reservoir. And the litter bag method was used to analyze the seasonal dynamics of litter decomposition and nutrient return. The results showed that: (1) the dynamics of the weightlessness rate for all three kinds of litter types were sharp during May to November because of higher temperature and large amount of precipitation. On the contrary, within the lower temperature and precipitation amount during November to March, its dynamic trend was gentle. Further more, the decomposition rates in different litter types showed the following order:Prunusdavidianalitter>Platycladusorientalis-Prunusdavidianamixed litters>PlatycladusOrientalislitter; (2) the litter nutrient content presented that: with the litter decomposition, the litter N content tend to increase and then decrease, showing the pattern of enrichment-release; while the P concentration in litter rose continually with the enrichment pattern; the K content showed the decrease trends from the beginning to the end under the release pattern; (3) the yearly nutrient return amounts in three kinds of forests showed the following order:PlatycladusOrientalis>Prunusdavidiana>Platycladusorientalis-Prunusdavidianamixed forest for the N return amount;Platycladusorientalis-Prunusdavidianamixed forest >PlatycladusOrientalis>Prunusdavidianafor the P return amount;Prunusdavidiana>Platycladusorientalis-Prunusdavidianamixed forest >PlatycladusOrientalisfor the K return amount. The mixed forest type could significantly increase the litter decomposition rates and N/P return amounts compared with the pure forest type (P<0.05).
Platycladusorientalis;Prunusdavidiana; litter decomposition; nutrient return
2014-12-10
国家自然科学基金项目(31170580)
吴 明(1989-),女,河南安阳人,硕士研究生,从事植被恢复的理论与技术研究。
杨喜田(1965-),男,河南长垣人,教授,博士生导师。
1000-2340(2015)03-0320-05
S718
A
