2014年夏季南海北部超微型浮游植物分布及环境因子影响
2015-06-01魏玉秋孙军丁昌玲
魏玉秋,孙军*,丁昌玲
(1.天津科技大学海洋与环境学院,天津 300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津 300457)
2014年夏季南海北部超微型浮游植物分布及环境因子影响
魏玉秋1,2,孙军1,2*,丁昌玲1,2
(1.天津科技大学海洋与环境学院,天津 300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津 300457)
利用流式细胞仪BD Accuri C6对2014年夏季南海北部超微型浮游植物进行了现场的观测研究,发现了3类超微型光合自养浮游植物,聚球藻(Synechococcus,Syn)、原绿球藻(Prochlorococcus,Pro)和超微型真核藻类(pico-eukaryotes,Euk),并对其丰度与分布以及环境因子影响进行了研究。结果表明,Syn、Pro和Euk丰度总平均值分别为5.13×103个/mL,3.27×104个/m L和1.85×103个/mL,碳生物量均值分别为1.19μg/L,1.86μg/L和4.51μg/L。Syn、Pro和Euk的丰度表现出不同的分布特征。Syn、Pro和Euk丰度分布趋势呈现近海低而外海高,Syn和Euk丰度高值区分别出现在沿岸带与陆架和上升流影响海域,Pro丰度高值区出现在沿岸带与陆架,低值区出现在上升流影响海域。Syn、Euk丰度高值区主要分布在次表层,Pro丰度高值区主要分布在真光层底部,Euk丰度垂直变化差异相对Syn和Pro较小。超微型浮游植物与环境因子的相关性分析结果表明,Syn、Pro和Euk的碳生物量均与硝酸盐、硅酸盐浓度和深度呈现负相关关系,Pro的碳生物量与磷酸盐浓度呈现正相关关系。
超微型浮游植物;聚球藻;原绿球藻;超微型真核藻类;南海
1 引言
海洋超微型浮游植物是一类细胞粒径小于2μm的放氧光合自养浮游生物,包括聚球藻(Synechococcus,Syn)、原绿球藻(Prochlorococcus,Pro)以及超微型真核藻类(pico-eukaryotes,Euk)[1—2]。海洋聚球藻和原绿球藻作为个体微小且独立生存的类群,其光合色素独特,能量转换效率高,代谢繁殖快,特别是原绿球藻每天分裂0.72次[3],生物量循环迅速[4],因此环境适应能力较强,多分布在寡营养盐海域中,细胞丰度相当高[5]。再加上其沉降速率慢,在真光层停留时间长[6],成为海洋寡营养盐海域浮游植物群落和微食物环的重要组成部分,贡献海洋总生物量的80%及初级生产力的40%[1],因而成为海洋浮游植物生产力和生物量的主要贡献者,在生态系统中的地位和作用越来越大。
南海是我国最大的边缘海,属于热带海洋性气候,是典型的寡营养盐海域。南海特殊的地理环境受到黑潮和东亚季风的影响,产生不同尺度的涡旋,呈现多涡的特点[7—10]。南海北部受到黑潮入侵以及内部中尺度涡的影响,形成结构复杂的上层环流,在季风和反气旋影响下的上升流和下降流作用明显,进一步影响着营养盐的输送以及超微型浮游植物的分布[11—14,10]。南海北部复杂的生态环境还表现在该海域营养盐丰富、盐度低;陆架中营养和中盐度及陆坡与外海的寡营养盐和高盐度;光照、温度、盐度、营养盐存在垂直分布的差异;水体层化和跃层明显等特点,造成南海北部超微型浮游植物在丰度分布范围上的差异[15]。
流式细胞仪检测技术(flow cytometry,FCM)可以根据浮游植物自发荧光的特性,反映出细胞大小、形状结构、色素类型以及内部结构,在现场能够迅速完成对单细胞浮游植物定量和定性的多参数分析研究,甚至能完成对细胞颗粒的分选[16]。杨燕辉和焦念志[17]利用流式细胞仪对胶州湾超微型浮游植物的群落分布和动态变化进行连续近一年的大面调查,发现胶州湾的超微型浮游植物主要由Syn和Euk组成,其叶绿素生物量占总叶绿素生物量的20%以上;乐凤凤等[18]采用流式细胞术测定了长江口附近海域超微型浮游生物丰度和碳生物量,并分析了其与环境因子之间的关系,发现Syn和Euk丰度呈现近海低外海高的分布趋势,Syn丰度最大值出现在真光层底部,而Euk丰度最大值主要出现在次表层,Syn和Euk的碳生物量与营养盐之间存在负相关关系;屈佩等[19]利用流式细胞术获取了夏季南黄海超微型浮游生物丰度数据,并分析了其组成和分布规律以及主要的环境影响因子,发现Syn和Euk受温度和光照的限制明显,调查海域营养盐浓度较高,不是其限制因子,但是能够促进超微型浮游植物的生长。本文通过对南海北部超微型浮游植物群落结构和分布调查分析研究,以期为该陆架生态系统积累生物背景资料,并结合其对环境因子的响应特征,为构建海洋生态模型提供基础参数。
2 材料与方法
2.1 研究海域和采样站位
2014年8月12日至9月4日搭乘“实验一号”考察船在南海北部及其邻近水域(18°~22°N、114°~116°E)进行了现场采样及样品处理。设1条调查断面,共8个采样站位(图1)。水样由CTD的Niskin采水器采集,各站位根据不同水深从表层、5 m、25 m、50 m、75 m、100 m、150 m、200 m水深进行采样。每层水样采取2 mL,现场应用流式细胞仪(BD Accuri C6)直接进行测定。同时,获取温度、盐度等数据。
图1 采样站位Fig.1 Sampling stations
2.2 海洋超微型浮游植物类群分析
海洋超微型浮游植物中含有自发荧光的蛋白,不需要加荧光素可以直接用流式细胞仪检测。通过BD Accuri C6软件记录波长(585±40)nm(FL2)的橙色荧光和大于波长670 nm(FL3)的红色荧光,用以表征藻红蛋白荧光及叶绿素荧光等数据[20]。应用流式细胞仪的FL2、FL3荧光检测器检测到这两种荧光信号,用于区分仅发射红色荧光的Syn[21],再利用细胞大小、叶绿素相对含量,区分Pro与Euk类群,细胞颗粒较小、叶绿素信号较弱的是Pro,细胞颗粒大、叶绿素含量相对较高的是Euk[15]。本次在南海北部现场应用流式细胞仪调查中,共检测到3类超微型浮游植物:Syn、Pro和Euk,它们在所有调查站位都有分布。
2.3 各类群生物量估算
各类超微型浮游植物的细胞丰度转化为碳生物量,由于各类超微型浮游植物细胞碳转换系数因不同海域而存在不同差异。本文选择国际上普遍使用的超微型浮游植物细胞碳转换系数:Syn的碳转换系数为250 fg/个;Pro的碳转换系数为53 fg/个;Euk的碳转换系数为2 100 fg/个[21—23]。
3 结果
3.1 水文环境特征
3.1.1 温盐参数
夏季南海北部及其邻近海域的水团主要受到近岸珠江口冲淡水和流场内形成的多个中尺度涡的影响。图2为夏季调查海域温盐分布情况。由图2可见,调查海域水温变化范围在14.3℃(I1站位的200 m)至30.0℃(K4站位的25 m)之间,次高温值29.9℃出现在K3站位的3 m水层,温度总平均值为22.1℃。水温从近岸到I1站位呈现递增趋势,高温主要出现在I1和K3站位,由K3站位到外海海域呈现递减趋势,低温出现在K2站位,变化趋势较小,同一平面温度差异较小。表层温度高,虽水深的增加而降低,温度跃层明显,水温具有显著的层化现象(图2a)。
盐度的变化范围在33.1(J3站位的2 m)至34.6(K2站位的140 m)之间,盐度总平均值为34.2,同一水层低盐度区位于调查海域靠近珠江口区域。其盐度变化趋势与温度基本一致,同一平面分布差异较小;表层盐度较低,随水深的增加而升高,同样具有明显的层化现象(图2b)。

图2 夏季调查海域温度(a)和盐度(b)断面分布Fig.2 The section distributions of temperature(a)and salinity(b)in the study area in summer.
3.1.2 营养盐参数
夏季南海北部营养盐分布受水文状况影响显著,该调查海域营养盐的分布主要受到近岸径流营养盐输入和沿岸反气旋型涡旋中心区域的影响。该调查断面硝酸盐浓度的分布变化范围在0.038μmol/L(K3站位的25 m)至18.769μmol/L(SEATS站位的200 m)之间,硝酸盐浓度总平均值为5.662μmol/L。硝酸盐的浓度分布在近岸较高,由近岸向外海呈现明显的降低趋势。硝酸盐浓度跃层大多在50~75 m水深,表层硝酸盐浓度分布较低,随水深的增加而增加,层化现象明显。
亚硝酸盐浓度的分布变化范围小于检测限至2.303μmol/L(J3站位的5 m)之间,亚硝酸盐浓度总平均值为0.182μmol/L。亚硝酸盐浓度高值区位于陆架中部的J3站位,外海的浓度分布偏低,其浓度大多低于检测限而检测不出。
硅酸盐浓度的分布变化范围在0.248μmol/L(J5站位的5 m)至22.202μmol/L(SEATS站位的200 m)之间,硅酸盐浓度总平均值为6.629μmol/L。硅酸盐浓度总的分布趋势是由近岸向外海逐渐减少,但是在I1站位硅酸盐浓度比周围海域明显高。表层硅酸盐浓度较低,其浓度随水深而增加,该调查海域具有明显的垂直层化现象,跃层大多分布在75 m水深,在I1站位50 m水深出现明显的跃层。
磷酸盐浓度分布范围在0.173μmol/L(J4站位的5m)至6.95μmol/L(I1站位的5 m)之间,次高值6.757μmol/L出现在K2站位的5 m,磷酸盐浓度总平均值为1.211μmol/L。整个调查海域磷酸盐浓度较高,外海相对较低。磷酸盐浓度高值区主要出现在I1、K2和K3站位。磷酸盐主要分布在75 m以浅水层,其高浓度区域主要成块状分布在5~50 m水深。
3.2 超微型浮游植物分布特征
夏季南海北部Syn细胞丰度变化范围在0.82× 102个/m L(K2站位的150 m)至4.54×104个/m L(I1站位的50 m),平均细胞丰度为5.13×103个/m L;水柱积分平均丰度变化范围在6.96×102~1.45×104个/m L,平均值为4.76×103个/mL,碳生物量均值则为1.19μg/L。调查海区的Syn细胞丰度数量级基本在1×103个/m L。近岸J3站位细胞丰度为1.45× 104个/mL,出现明显的高值区;次高值区在I1站位,细胞丰度为8.03×103个/m L;低值区出现在外海SEATS站位,细胞丰度为6.96×102个/m L;高值区比低值区高2个数量级,Syn细胞丰度呈现由近岸向外海逐渐降低的分布趋势(图3,表1)。
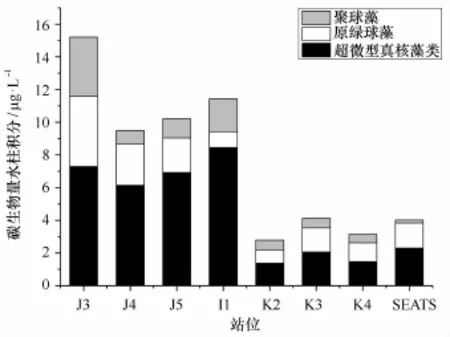
图3 三类超微型浮游植物碳生物量水柱积分分布Fig.3 The carbon biomass distributions of water column integral of three picoplankton groups
Pro细胞丰度变化范围在2.30×102个/m L(K2站位的200 m)至1.14×105个/mL(K3站位的50 m),总平均细胞丰度为3.27×104个/m L;水柱积分平均丰度变化范围1.50×104~8.11×104个/m L,平均值为3.50×104个/m L,碳生物量均值则为1.86 μg/L。 Pro水柱积分细胞丰度数量级均在1×104个/m L,在外海Pro细胞丰度比Syn高出1个数量级,比Euk高出2个数量,与Jiao和Yang等[3]对中国外海Pro、Syn、Euk数量研究一致。该调查海域Pro细胞丰度分布趋势呈现近岸高外海低,高值区出现在近岸J3站位,细胞丰度为8.11×104个/mL;低值区出现在大陆坡的I1和K2站位,细胞丰度分别为1.82×104个/m L、1.50×104个/m L。外海海域K3、K4和SEATS站位的细胞丰度大体相当,细胞丰度分布范围在2.15 ×104~2.91×104个/m L(图3,表1)。
Euk细胞丰度变化范围在0.15×102个/mL(SEATS站位的200 m)至2.78×104个/m L(I1站位的50 m),总平均细胞丰度为1.85×103个/m L;水柱积分平均丰度变化范围在6.56×102~4.03×103个/mL,平均值为2.15×103个/mL,碳生物量均值则为4.51μg/L。Euk细胞丰度分布的变化趋势是近岸高外海低,Euk高值区主要分布在近岸,细胞丰度在2.93×103~4.03×103个/mL;外海Euk分布相对较少,细胞丰度为1×102个/mL,近岸比外海高1个数量级(图3,表1)。

表1 三类超微型浮游植物细胞丰度水柱积分分布Tab.1 The cell abundance distributions of water column integral of three picoplankton groups
为便于阐明观测结果,将夏季南海北部研究断面按照自然地理区划分为小于200 m水深的沿岸带与陆架(J3、J4、J5、I1)和大于200 m水深的陆坡与开阔大洋(K2、K3、K4、SEATS)两部分[15]。Syn细胞丰度主要分布范围在0~75 m水深,各站位高值区主要出现在50 m水层,平均值为1.78×104个/mL,最大值出现在I1站位50 m水层,细胞丰度为4.54×104个/m L。该调查海域Syn细胞丰度随水深的增加,呈现先增加后降低的趋势(图4)。在整个调查海域Syn细胞丰度主要分布在100 m以浅水层,在沿岸带与陆架各水层Syn细胞丰度高,在50 m水层的平均细胞丰度为2.30×104个/m L,分布深度可达100 m;陆坡与开阔大洋各水层Syn细胞丰度低,在50 m水层的平均细胞丰度为1.27×104个/m L,分布深度多在75 m以浅水层。Syn丰度分布特点是沿岸带与陆架细胞丰度高,陆坡与开阔大洋细胞丰度低(图5a)。
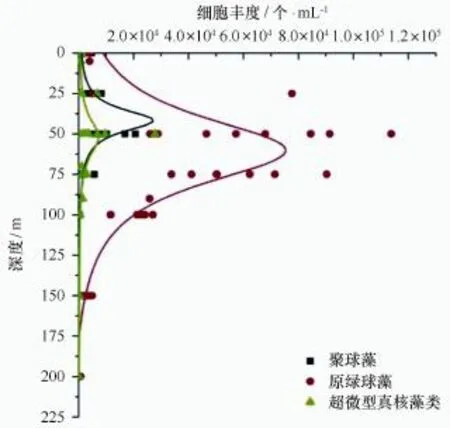
图4 三类超微型浮游植物细胞丰度垂直分布(所有采样站位汇总)Fig.4 The vertical distributions of cell abundance of three picoplankton groups(all sampling stations summary)
Pro细胞丰度主要分布范围在0~150 m水深,高值区出现在50 m水层,平均值为6.46×104个/m L,次高值区出现在75 m水层,平均值为5.71×104个/m L。该调查海域Pro细胞丰度随水深的增加,呈现先增加后降低的趋势(图4)。在整个调查海域Pro成块状分布,主要分布在150 m以浅水层,甚至在外海SEATS站位200 m水层可检测到Pro,其细胞丰度为6.74×102个/m L,高值区主要分布在沿岸带与陆架和外海的SEATS站位。Pro在沿岸带与陆架50 m层的平均细胞丰度为6.99×104个/m L;在陆坡与开阔大洋50 m层的平均细胞丰度为5.92×104个/m L。夏季该研究海域Pro丰度分布特点是沿岸带与陆架细胞丰度高,陆坡与开阔大洋细胞丰度低,与Syn和Euk相比,其在整个海域分布范围广,细胞丰度高(图5b)。
Euk细胞丰度主要分布范围在0~100 m水深,高值区出现在50 m水层,平均值为7.76×103个/m L,最大值出现在I1站位50 m水层,细胞丰度为2.78×104个/mL。该调查海域Euk细胞丰度随水深增加,呈现先增加后降低的趋势,但是相比Syn、Pro的变化趋势,Euk细胞丰度分布比较均匀,峰值不是很突出,除在沿岸带与陆架的I1站位峰值较高,在陆坡与开阔大洋没有见到很明显的高峰层(图4)。该调查海域Euk细胞丰度主要分布在100 m以浅,沿岸带与陆架各水层分布较高,在50 m水层,Euk平均细胞丰度为1.47×104个/mL;陆坡与开阔大洋各水层分布较低,在50 m水层,平均细胞丰度为2.52×103个/m L。Euk丰度分布特点是沿岸带与陆架细胞丰度高,陆坡与开阔大洋细胞丰度低,两个区域在50 m层平均细胞丰度相差1个数量级(图5c)。
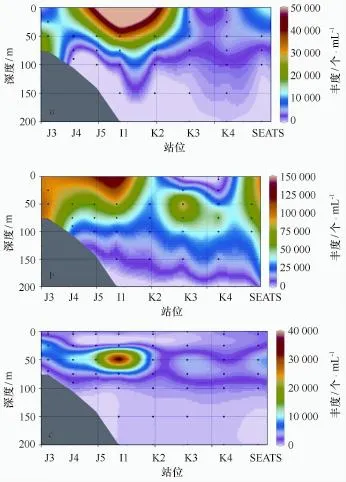
图5 聚球藻(a)、原绿球藻(b)和超微型真核藻类(c)细胞丰度断面分布Fig.5 The section distributions of cell abundance of three picoplankton groups(a.Syn,b.Pro,c.Euk)
3.3 超微型浮游植物碳生物量与环境因子的关系
表2为夏季南海北部调查海域超微型浮游植物碳生物量与环境因子的相关性。该研究的环境因子主要包括:温度、盐度、深度以及营养盐(硝酸盐、亚硝酸盐、硅酸盐和磷酸盐)浓度。由表2可知,Syn、Pro碳生物量与硝酸盐、硅酸盐、深度有显著的负相关关系,Syn与温度存在正相关关系;Pro与亚硝酸盐、磷酸盐存在正相关关系;Euk碳生物量与硝酸盐有显著的负相关关系。
4 讨论
南海北部位于119°E以西,由黑潮入侵所带来的温度、盐度和营养盐等不同的变化对该研究海域影响较小[25—26]。该研究海域营养盐浓度在水平分布上呈明显的近岸高外海低的分布趋势,主要是珠江河口冲淡水对营养盐的输入;受反气旋型涡旋中心区域的影响,高温、低盐以及低营养盐的表层水向下扩张,营养盐等深线出现明显的下移现象,50 m以浅水层营养盐较低,其含量常低于常规分光光度法检测限[27—28]。
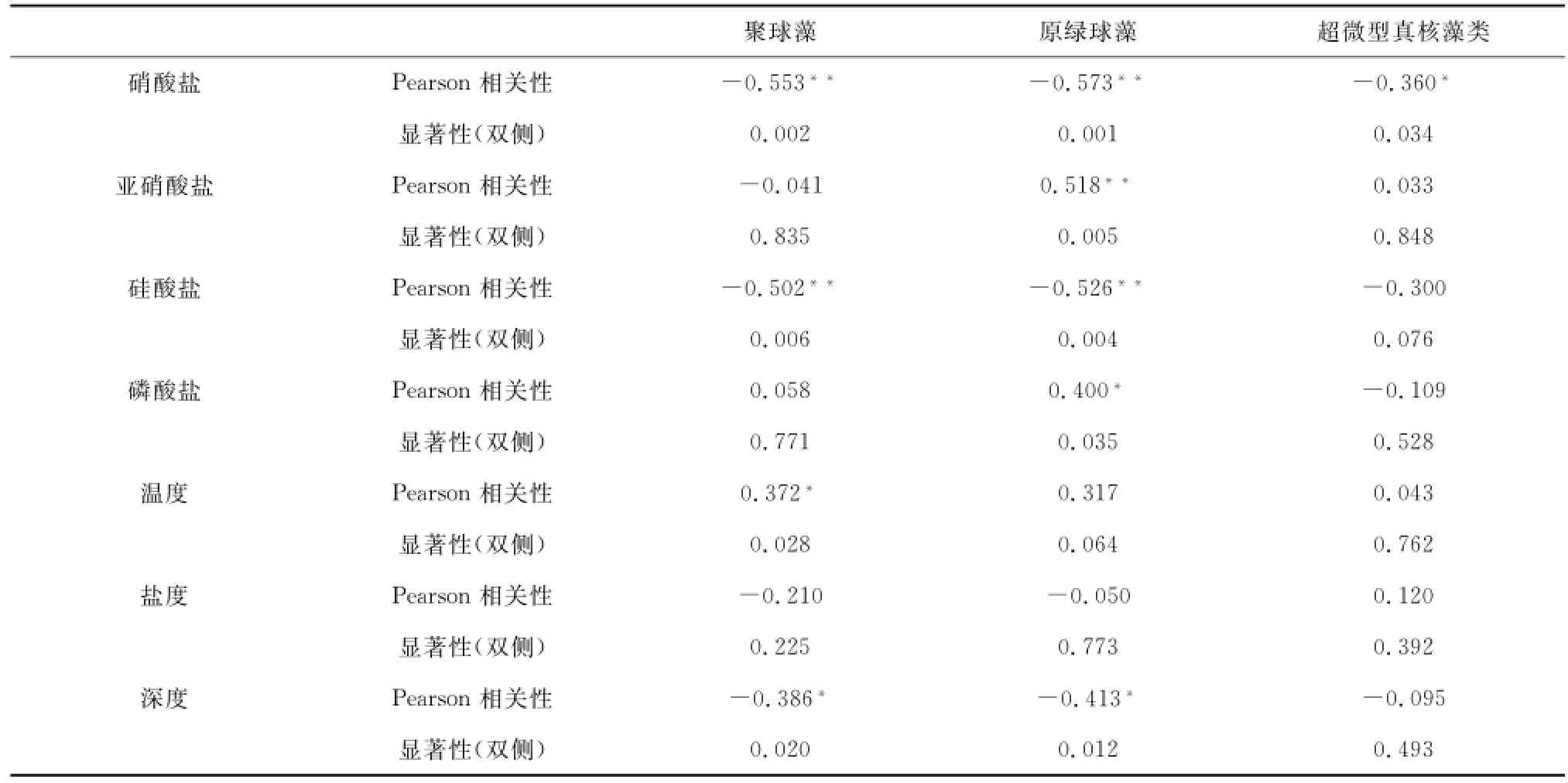
表2 超微型浮游植物碳生物量与环境因子的Pearson相关性Tab.2 Pearson correlation analysis between picoplankton carbon biomass and environmental factors
由图2温盐分布图可知,在K2站位从25 m到50 m为高温低盐区域,可能与反气旋型涡旋中心区域的影响有关。但是其向下扩张的变化趋势较小,是因为该海域温盐分布同时受到由流场内多个中尺度涡作用下产生不同水团(近岸混合水团,南海表层、次表层、中层、深层水团)的影响,其表层水团具有分布范围广,变性强烈等特点,影响了调查海区大部分海域[26],同时夏季南海北部海面上始终存在着季节性上升流的影响[29],因此同平面温盐分布差异较小。该海域营养盐跃层多分布50~75 m水深,亚硝酸盐浓度甚至低于检测限。上升流具有显著的低温、高盐、高磷、低溶氧等特征,而该研究海域I1、K2站位表层磷酸盐浓度普遍较高,硅酸盐在I1站位跃层呈现明显的上升趋势,具有部分上升流特征,主要原因是该研究海域受反气旋型涡旋和上升流共同影响,且上升流的影响大于反气旋型涡旋该的影响[28—30]。总之,该研究海域复杂的环境变化,造成了三类超微型浮游植物在丰度分布范围上的差异。
Syn、Pro在南海北部调查区域的分布特征,主要是受环境因子的影响。Syn的最适生长温度在20~26℃;Pro的适合生长温度在10~30℃,其自然地理分布受到海水温度的调控,通常当水温低于下限时,极少有原绿球藻存在[22]。夏季该研究海域温度变化范围在14.3~30.0℃之间,同一平面温度分布差异较小(图2a),因此温度并不是影响其水平分布变化的限制因子。前期基于分子系统学的研究将Syn分为MC-A(marine Cluster A)、MC-B(marine Cluster B)和MC-C(marine Cluster C)三类,其中MC-A适应高盐环境,MC-B耐盐但生长适应低盐环境;对MC-C的研究不多[31],Syn不同的进化类型可适应不同盐度的环境;Pro虽然主要分布在大洋的寡营养区域,但是在东海冲淡水稀释区域也有发现,因此其生长也对盐度有着较广范围的适应性[32]。该调查海域盐度的变化范围在33.1~34.6之间,变化范围较小,盐度同样不是影响其水平分布变化的限制因子。
营养盐是Syn、Pro生长繁殖的必需条件[33],通过Syn、Pro碳生物量与营养盐相关性分析发现,Syn、Pro与硝酸盐、硅酸盐有显著的负相关关系(表2)。Pro与硝酸盐浓度负相关,其可能原因是Pro缺少编码硝酸盐还原酶基因,不能利用硝酸盐[32],而Syn的氮源主要是铵和硝酸盐,虽然在某些条件下相关性可能只是表象,但这些相关性却能反映出该调查海域并非受到硝酸盐和硅酸盐限制[34]。
磷酸盐作为Pro另一个限制性营养元素,会直接影响到其生长繁殖,研究表明磷元素对Pro生长的作用并不明显,Pro对磷元素的需求较小,只有磷酸盐浓度低于0.03μmol/L时,磷才会成为Pro的限制因子[26,35—36]。本研究通过相关性分析,发现Pro碳生物量与磷酸盐存在正相关关系,该调查海域由于受到珠江口冲淡水和上升流的影响,在其影响范围内磷酸盐的浓度相对较高,说明磷酸盐不再成为限制性营养盐。该调查海域受上升流影响最明显的I1、K2站位,其磷酸盐浓度最高,平均值为2.87μmol/L、2.61 μmol/L,而K3站位受上升流的影响较小,磷酸盐浓度平均值1.46μmol/L。研究证据表明,适宜Pro生长繁殖的磷酸盐浓度范围在0~0.1μmol/L,高浓度的磷酸盐会抑制Pro的生长繁殖[36],在东海和南海海域,Pro的丰度分布和生长率与磷酸盐存在负相关关系[37],因此就不难解释在I1、K2站位,Pro细胞丰度最低,而K3站位的Pro细胞丰度稍高的现象。
夏季珠江河口径流量增大,大量的营养盐随其输入,该海域并不是典型的寡营养盐水域[22],近岸水体含丰富的营养盐,为Syn、Pro的生长繁殖提供了良好的生存环境,其分布沿着营养盐浓度梯度,呈现近岸高外海低分布趋势。有研究发现在近岸河口营养盐丰富的区域,Syn、Pro细胞丰度却很低,这与河口区域悬浮颗粒物对光的可得性的限制有关[38],但是该调查海域J3站位位于陆架中部,该区受悬浮颗粒物对光的可得性的限制的影响较小,水体透明度高,营养盐丰富,Syn、Pro在该站位细胞丰度最高,分别为1.45×104个/m L和8.11×104个/m L。SEATS站位Syn细胞丰度低,为6.96×102个/m L。在图5 I1站位Syn细胞丰度出现明显的高值,为8.03×103个/m L,原因可能是I1站位处在上升流影响区域,由上升流所携带丰富的营养盐,为Syn的生长繁殖提供了有利条件。
光照是控制Syn、Pro丰度分布的关键因子,主要是影响Syn、Pro的垂直分布。Syn对光的不同适应,演化出有利于光能吸收的藻红蛋白(phyeoerythrin,PE)和藻蓝蛋白(phyeoeyanin,PC)。Pro具有较低的光饱和强度,对强光极为敏感,演化出高光型和低光型两种生态类型[22]。夏季南海北部光照较强,强光对Syn、Pro具有抑制作用[38],因此该研究海域表层的Syn、Pro细胞丰度低,在加上50 m水深存在营养盐跃层,Syn、Pro细胞丰度多分布在50 m水深。Pro可捕捉真光层底部极微弱的光进行高效的光合作用,Pro比Syn在真光层底部具有更强的适应性和竞争优势[22,39],因此Pro在75 m等深线同样存在较高的细胞丰度。这与焦念志等[22]在中国南海海域研究Syn、Pro细胞丰度垂直分布类型一致,该类型常出现在热带、亚热带寡营养盐海域。
通过Syn、Pro碳生物量与深度相关性分析发现,与深度有显著的负相关关系,随水深的增加,温度减小,光照强度减弱。Syn随水深的变化,受温度、光照以及营养盐的影响,在沿岸带与陆架富营养区域分布深度可达100 m,陆坡与开阔大洋寡营养区域主要位于75 m以浅水层;Pro随水深的变化,受到温度和光照的影响,多分布在150 m以浅水层。Syn、Pro在不同的自然地理区对深度存在不同适应性和分布格局,说明Syn、Pro的垂直分布变化趋势同时受到温度、光照和营养盐共同影响。
Euk具有较强的环境适应能力,即使在寡营养盐海域都可检测到Euk的存在,夏季温度不再是其限制因子,但是前期研究表明其生长对营养盐的依赖性比较大[27,39]。Euk在沿岸带与陆架区域有着较高的丰度,该海区珠江口冲淡水输入大量的营养盐,可见营养盐对该区域Euk的生长是具有促进作用。整个调查海区Euk的分布受到温度和光照影响相对较大。由于温度和光照的限制,Euk主要分布在100 m以浅水层,丰度高值区在50 m真光层以上。需要指出的是I1站位Euk的细胞丰度都比较高,可能是受到该处上升流的影响。
5 结论
(1)本次2014年夏季南海北部现场应用流式细胞仪调查中,共检测到3类超微型浮游植物:Syn、Pro和Euk,且它们在所有调查站位都有分布。Syn、Pro和Euk丰度分布呈现出近海低而外海高的分布趋势,Syn和Euk丰度高值区分别出现在沿岸带与陆架和上升流影响海域,Pro丰度高值区出现在沿岸带与陆架,低值区出现在上升流影响海域;Syn、Euk丰度高值区主要分布在次表层,Pro丰度高值区主要出现在真光层底部。
(2)3类超微型浮游植物丰度分布变化由温度、光照以及珠江口冲淡水和上升流所引起营养盐的变化共同影响。
致谢:同济大学张传伦教授提供营养盐数据,“实验1”号科研调查船为本次调查提供了宝贵的采样平台,谨致谢忱!
[1] Olson R J,Chisholm S W,Zettler E R,et al.Spatial and temporal distributions of prochlorophyte picoplankton in the North Atlantic Ocean[J].Deep-Sea Research Part A.Oceanographic Research Papers,1990,37(6):1033-1051.
[2] Zubkov M V,Sleigh M A,Tarran G A,et al.Picoplanktonic community structure on an Atlantic transect from 50°N to 50°S[J].Deep-Sea Research PartⅠ:Oceanographic Research Papers,1998,45(8):1339-1355.
[3] Jiao N Z,Yang Y H.Ecological studies onProchlorococcusin China seas[J].Chinese Science Bulletin,2002,47(15):1243-1250.
[4] 宁修仁,沃洛D.长江口及其毗连东海水域蓝细菌的分布和细胞特性及其环境调节[J].海洋学报,1991,13(4):552-559.
Ning Xiuren,Vogel D.The distribution and environmentally regulating mechanism of the Cyanobacteria with its cell characteristic in Changjiang Estuary and its adjacent waters[J].Haiyang Xuebao,1991,13(4):552-559.
[5] McManus G B,Dawson R.Phytoplankton pigments in the deep chlorophyll maximum of the Caribbean Sea and the western tropical Atlantic Ocean[J].Marine Ecology Progress Series,1994,113(1):199-206.
[6] 李超伦,栾凤鹤.东海春季真光层分级叶绿素a分布特点的初步研究[J].海洋科学,1998(4):59-62.
Li Chaolun,Luan Fenghe.A proliminary study on the distribution of size-fractionated chlorophyllain the eu-photic zone of the East China Sea in spring[J].Marine Science,1998(4):59-62.
[7] 蔡树群,苏纪兰,甘子钧,等.冬季南海上层环流动力机制的数值研究[J].海洋学报,2001,23(5):14-23.Cai Shugun,Su Jilan,Gan Zijun,et al.The numerical study on the dynamic mechanism of the South China Sea upper circulation in winter[J].Haiyang Xuebao,2001,23(5):14-23.
[8] Wang G H,Su J L,Qi Y Q.Advances in studying mesoscale eddies in South China Sea[J].Advances in Earth Science,2005,20(8):882-886.
[9] 乐凤凤,宁修仁,刘诚刚,等.2006年冬季南海北部浮游植物生物量和初级生产力及其环境调控[J].生态学报,2008,28(11):5775-5784.
Le Fengfeng,Ning Xiuren,Liu Chenggang,et al.Standing stock and production of phytoplankton in the northern South China Sea during winter of 2006[J].Acta Ecologica Sinica,2008,28(11):5775-5784.
[10] 孙军,宋书群,乐凤凤,等.2004年冬季南海北部浮游植物[J].海洋学报,2007,29(5):132-145.
Sun Jun,Song Shuqun,Le Fengfeng,et al.Phytoplankton in northern South China Sea in the winter of 2004[J].Haiyang Xuebao,2007,29(5):132-145.
[11] 高劲松.南海北部中尺度涡及北部湾环流结构与生成机制研究[D].青岛:中国海洋大学,2013.
Gao Jinsong.The research on the eddy in the South China Sea and circulation in the Beibu Gulf[D].Qingdao:Ocean University of China,2013.
[12] Xue H J,Chai F,Pettigrew N,et al.Kuroshio intrusion and the circulation in the South China Sea[J].Journal of Geophysical Research,2004,109(C2),doi:10.1029/2002JC001724.
[13] Wu C R,Chiang T L.Mesoscale eddies in the northern South China Sea[J].Deep-Sea Research PartⅡ:Topical Studies in Oceanography,2007,54(4/5):1575-1588.
[14] Liu Q Y,Kaneko A,Su J L.Recent progress in studies of the South China Sea circulation[J].Journal of Oceanography,2008,64(5),753-762.
[15] 宁修仁,蔡昱明,李国为,等.南海北部微微型光合浮游生物的丰度及环境调控[J].海洋学报,2003,25(3):83-97.
Ning Xiuren,Cai Yuming,Li Guowei,et al.Photosynthetic Picoplankton in the northern South China Sea[J].Haiyang Xuebao,2003,25(3):83-97.
[16] 刘昕,张俊彬,黄良民.流式细胞仪在海洋生物学研究中的应用[J].海洋科学,2007,31(1):92-96.
Liu Xin,Zhang Junbin,Huang Liangmin.Applications of flow cytometry(FCM)in researches of marine biology[J].Marine Sciences,2007,31(1):92-96.
[17] 杨燕辉,焦念志.流式细胞仪检测的胶州湾超微型浮游植物的分布和动态变化[C]//中国海洋与湖沼学会甲壳动物学分会、中国动物学会、中国海洋与湖沼学会生态学分会2000年学术研讨会论文摘要集.青岛:中国海洋湖沼学会,2000.
Yang Yanhui,Jiao Nianzhi.The distribution and dynamic change of picophytoplankton in Jiaozhou Gulf by flow cytometry(FCM)[C]//China ocean and limnetic society zoology section of the shell,China Zoological Society,Ecology branch of China Marine and lacustrine bog institute in 2000 attended the academic seminar on abstract set.Qingdao:Chinese Society for Oceanology and Limnology,2000.
[18] 乐凤凤,刘诚刚,郝锵,等.2006年秋季长江口及邻近海域微微型浮游生物的分布特征及环境影响因素[J].海洋学研究,2011,29(3):108-117.
Le Fengfeng,Liu Chenggang,Hao Qiang,et al.Distribution and environmental impact factors of picoplankton in Changjiang River Estuary and its adjacent sea areas in autumn of 2006[J].Journal of Marine Sciences,2011,29(3):108-117.
[19] 屈佩,王宗灵,庞敏,等.夏季南黄海主要环境因子对微微型浮游生物分布影响[J].海洋学报,2013,35(4):176-187.
Qu Pei,Wang Zongling,Pang Min,et al.Influence of environmental factors on picoplankton in the southern Yellow Sea in summer[J].Haiyang Xuebao,2013,35(4):176-187.
[20] Marie D,Partensky F,Vaulot D,et al.Enumeration of phytoplankton,bacteria,and viruses in marine samples[M]//Robinson J P,Darzynkiewicz Z,Dean P N,et al.Current protocols in cytometry.New York:John Wiley&Sons,Inc,1999.
[21] Phinney D A,Cucci T L.Flow cytometry and phytoplankton[J].Cytometry,1989,10(5):511-521.
[22] 焦念志.海洋微型生物生态学[M].北京:科学出版社,2006.
Jiao Nianzhi.Marine Picoplankton Ecology[M].Beijing:China Science Press,2006.
[23] Buck K R,Chavez F P,Campbell L.Basin-wide distributions of living carbon components and the inverted trophic pyramid of the central gyre of the North Atlantic Ocean,summer 1993[J].Aquatic Microbial Ecology,1996,10(3):283-298.
[24] Campbell L,Nolla H A,Vaulot D.The importance ofProchlorococcusto community structure in the central North Pacific Ocean[J].Limnology and Oceanography,1994,39(4):954-961.
[25] 程国胜,孙佳东,俎婷婷,等.2011年夏季南海北部海区水团分析[J].热带海洋学报,2014,33(3):10-16.
Cheng Guosheng,Sun Jiadong,Zu Tingting,et al.Analysis of water masses in the northern South China Sea in summer 2011[J].Journal of Tropical Oceanography,2014,33(3):10-16.
[26] Jiao N Z,Yang Y H,Hong N,et al.Dynamics of autotrophic picoplankton and heterotrophic bacteria in the East China Sea[J].Continental Shelf Research,2005,25(10):1265-1279.
[27] 袁梁英.南海北部营养盐结构特征[D].厦门:厦门大学,2005.
Yuan Liangying.Distribution and characteristics of nutrients in the Northern South China Sea[D].Xiamen:Xiamen University,2005.
[28] 石晓勇,李鸿妹,韩秀荣,等.夏季南海北部典型中尺度物理过程对营养盐及溶解氧分布特征的影响[J].环境科学学报,2014,34(3):695-703.
Shi Xiaoyong,Li Hongmei,Han Xiurong,et al.Influence of typical mesoscale oceanographical process on the distribution of nutrients and dissolved oxygen in the Northern part of South China Sea in summer[J].Acta Scientiae Circumstantiae,2014,34(3):695-703.
[29] 于文泉.南海北部上升流的初步探讨[J].海洋科学,1987,6(11):7-10.
Yu Wenquan.A preliminary approach of the upwelling for the northern South China Sea[J].Marine Science,1987,6(11):7-10.
[30] 韩舞鹰,马克美.粤东沿岸上升流的研究[J].海洋学报,1988,10(1):53-59.
Han Wuying,Ma Kemei.The study of the coastal upwelling of Guangdong[J].Haiyang Xuebao,1988,10(1):53-59.
[31] 陈敏艺,袁洁,陈月琴,等.海洋超微型浮游植物遗传多样性的分子系统学研究进展[J].自然科学进展,2005,15(9):1032-1041.
Chen Minyi,Yuan Jie,Chen Yueqin,et al.Genetic diversity research progress of marine picophytoplankton molecular systematics[J].Advances in Natural Science,2005,15(9):1032-1041.
[32] 王丽,焦念志.海洋原绿球藻对环境的适应机制[J].自然科学进展,2009,19(6):591-598.
Wang Li,Jiao Nianzhi.The adaptation mechanism of Prochlorococcus to the environment[J].Advances in Natural Science,2009,19(6):591-598.
[33] Chiang K P,Kuo M C,Chang J,et al.Spatial and temporal variation of theSynechococcuspopulation in the East China Sea and its contribution to phytoplankton biomass[J].Continental Shelf Research,2002,22(1):3-13.
[34] Lee Chen Y L,Chen H Y,Lee W H,et al.New production in the East China Sea,comparison between well-mixed winter and stratified summer conditions[J].Continental Shelf Research,2001,21(6/7):751-764.
[35] 焦念志,杨燕辉.中国海原绿球藻研究[J].科学通报,2002,47(7):485-491.
Jiao Nianzhi,Yan Yanhui.Ecological studies onProchlorococcusin China seas[J].Chinese Science Bulletin,2002,47(15):1243-1250.
[36] 蒋霞敏.原绿球藻的生态条件研究[C]//中国生态学会2006学术年会论文荟萃.北京:中国生态学学会,2006.
Jiang Xiamin.The study on ecology condition ofProchlorococcus[C]//China's Ecological Society 2006 Annual Meeting of the Academic Paper.Beijing:Ecological Society of China,2006.
[37] Liu A K,Chang Y S,Hsu M K,et al.Evolution of nonlinear internal waves in the East and South China Seas[J].Journal of Geophysical Research,1998,103(C4):7995-8008.
[38] 杨琳,汪岷,刘冠群,等.胶州湾微微型浮游植物丰度及其与环境因子的相关性分析[J].海洋与湖沼,2012,43(5):967-975.
Yang Lin,Wang Min,Liu Guanqun,et al.The abundance of picophytoplankton and correlation analysis with environmental factors in Jiaozhou Bay[J].Oceanologia et Limnologia Sinica,2012,43(5):967-975.
[39] 汪岷,白晓歌,梁彦韬,等.北黄海夏季微微型浮游植物的分布[J].植物生态学报,2008,32(5):1184-1193.
Wang Min,Bai Xiaoge,Liang Yantao,et al.Summer distribution of picophytoplankton in the north Yellow Sea[J].Journal of Plant Ecology(Chinese Version),2008,32(5):1184-1193.
Distribution and environmental impact factors of picoplankton in the northern South China Sea in summer 2014
Wei Yuqiu1,2,Sun Jun1,2,Ding Changling1,2
(1.College of Marine and Environmental Sciences,Tianjin University of Science and Technology,Tianjin 300457,China;2.Tianjin Key Laboratory of Marine Resources and Chemistry,Tianjin University of Science and Technology,Tianjin 300457,China)
During summer of 2014,an investigation on the abundance and distribution of the three photosynthetic picoplankton groups ofSynechococcus(Syn),Prochlorococcus(Pro)and pico-eukaryotes(Euk)in the northern South China Sea and their environmentally regulating mechanisms was carried out using flow cytometry BD Accuri C6.The results show that the average abundances of Syn,Pro and Euk were 5.13×103cells/m L,3.27×104cells/m L and 1.85×103cells/m L,and the average abundances of carbon biomass were 1.19μg/L,1.86μg/L and 4.51 μg/L,respectively.The cell abundance of each picoplankton group showed different distribution characteristics.The abundance of Syn,Pro and Euk was high nearshore and low off-shore.The highest abundances of Syn and Euk population both appeared in coast and continental shelf and upwelling waters.The highest abundance of Pro was observed along coast and continental shelf,while the lowest Pro population abundance was observed in upwelling waters.High values of Syn and Euk were mainly found at the surface,while high values of Pro mainly occurred at the bottom of euphotic layer.The variation in Euk distribution was less than those of Syn and Pro.The results of environmental factor correlation analysis indicated close negative correlations between picoplankton carbon biomass and nitrate concentration,silicate concentration and depth,and a significant positive correlation between picoplankton carbon biomass and phosphate concentration.
picoplankon;Synechococcus;Prochlorococcus;pico-eukaryotes;South China Sea
Q948.8
A
0253-4193(2015)12-0056-10
魏玉秋,孙军,丁昌玲.2014年夏季南海北部超微型浮游植物分布及环境因子影响[J].海洋学报,2015,37(12):56—65,
10.3969/j.issn.0253-4193.2015.12.006
Wei Yuqiu,Sun Jun,Ding Changling.Distribution and environmental impact factors of picoplankton in the northern South China Sea in summer 2014[J].Haiyang Xuebao,2015,37(12):56—65,doi:10.3969/j.issn.0253-4193.2015.12.006
2015-05-18;
2015-10-08。
国家海洋局公益性项目(201105021-03);国家重点基础研究发展计划(2015CB954002);教育部新世纪优秀人才计划(NCET-12-1065);国家自然科学基金(41176136,41276124)。
魏玉秋(1991—),男,山东省五莲县人,从事浮游植物生态学研究。
*通信作者:孙军,教授,从事海洋生态学研究。E-mail:phytoplankton@163.com
