琼东海域今生颗石藻群落研究
2015-06-01刘海娇孙军冯媛媛
刘海娇,孙军*,冯媛媛
(1.天津科技大学海洋与环境学院,天津 300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津 300457)
琼东海域今生颗石藻群落研究
刘海娇1,2,孙军1,2*,冯媛媛1,2
(1.天津科技大学海洋与环境学院,天津 300457;2.天津科技大学天津市海洋资源与化学重点实验室,天津 300457)
通过2012年7、8月在南海北部陆架琼东上升流区域的水文、化学和生物的现场综合采样调查,对采集过滤的今生颗石藻膜样在偏振光显微镜下鉴种并统计细胞丰度,分析了夏季该海域的颗石藻群落结构特征及分布规律。结果表明,夏季琼东上升流区共检出今生颗石藻34种,优势物种有赫氏艾密里藻(Emilianiahuxleyi)、大洋桥石藻(Gephyrocapsaoceanica)、纤细伞球藻(Umbellosphaeratenuis)、卡特螺旋球藻(Helicosphaeracarteri)、地中海花冠球藻(Coronosphaeramediterranea)等,颗石粒细胞丰度介于0~2 040.23×103粒/L,平均值8.10×103粒/L,颗石球丰度介于0~68.90×103cells/L,平均值5.01×103cells/L。大多数颗石藻集中分布在上升流信号强的水体中。琼东中部水域的生物多样性指数H′较雷州半岛以东水域高,颗石粒群落的均匀度指数J分布与H′呈镶嵌模式。聚类Cluster和多维定标MDS结果,将颗石粒和颗石球群落各分成4个和5个组群,经相似性分析(ANOSIM)检验证明此种划分是显著的。将生物与海区环境因子进行冗余分析(Redundancy analysis,RDA)结果表明,N/P、PO3-4、NO-2和SiO2-3是影响本次调查的今生颗石藻生物群落的主控因子。
颗石藻;群落结构;优势种;琼东上升流;聚类分析;冗余分析
1 引言
今生颗石藻是一类具有附着鞭毛的单细胞微藻,含有叶绿素a和c[1]。分类地位上属于定鞭藻门颗石藻纲(Haptophyta,Coccolithophyceae)。在生活史特定阶段通过生物矿化作用生成复杂精美的钙质鳞片(颗石粒,coccolith),其具有独特的碳酸盐反向泵机制,会释放CO2,对海洋碳源汇格局的调控起到双重作用。研究表明,由于海气界面CO2平衡时间尺度较小,停留时间短,导致陆架区成为CO2的源[2]。颗石藻独特的矿化过程在海洋酸化的背景下对海洋碳循环的影响与日俱增[3—5],它们种类丰富,生物量大,是世界海洋中最高产的钙质功能类群[6],和珊瑚礁和有孔虫等钙质生物一起影响着海水的碳酸盐和钙离子化学平衡,碳酸钙(CaCO3)的溶解导致南海次表层出现过多的钙离子(Ca2+)和较高的总碱度(TAlk)[7]。水体中今生颗石藻也可以指导古海洋沉积物的研究,以往对颗石粒研究较多,最近发现也有保存完好的颗石球细胞,比如很典型的几个今生种类(Braarudosphaera,Coccolithus,Helicosphaera,Reticulofenestra)[8];同时水体中也会出现“活化石”种类,Hagino等人在日本近岸和克罗地亚外海水体中调查发现今生种TergestiellaadriaticaKamptner,通过形态鉴定和分子系统发生分析该种是由已灭绝的化石种Cyclagelosphaerasp.演变而来[9]。
中尺度涡(mesoscale eddy)是南海最重要的物理现象之一,对海洋循环发挥重要作用,从卫星海表高度(sea surface height,SSH)图来看,南海和西北太平洋是强的涡旋活动区[10],琼东附近充满温度低且富营养的冷涡中心,气旋涡存在的地方其叶绿素和初级生产为比非涡旋的地方高,是控制生物生产的关键过程[11]。因其涉及渔业捕捞量等实际生产问题,上升流是世界海洋中最为关注的现象之一。受东亚西南季风影响,南海北部形成粤东、琼东以及台湾海峡等沿岸上升流[12—15]。其中琼东上升流是夏季出现在海南岛以东的季节性沿岸上升流,此前研究并不多,自2006年以来才开始较为系统的调查[16]。相比冲淡水区域,南海近岸上升流区的溶解无机碳(DIC)和总碱度(TAlk)较高,这将影响此区域颗石藻的生长和分布[17]。
本文通过对南海琼东上升流区今生颗石藻群落结构及空间分布的研究,并结合同步环境资料进行综合分析,以期为琼东海区开展今生颗石藻在酸化等气候背景下的生理变化和生态机制研究累积基础数据和经验。
2 材料方法
2.1 站位
2012年7月30日到8月17日搭载“东方红2”号科考船于南海北部(18°~22°N,110°~116°E)设置如图1所示的现场调查站位,分别进行水文、化学和生物的现场综合调查。
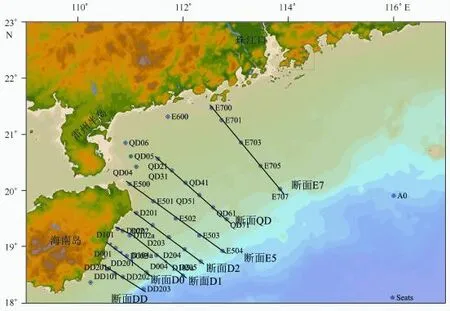
图1 2012年夏季琼东上升流海域调查站位Fig.1 Sampling sites in Qiongdong upwelling water during summer 2012
2.2 样品采集和处理
调查现场使用Seabird 917Plus CTD莲花型采水器,同期进行了水文、化学参数调查,温度、盐度和深度数据来自CTD,采样标准层次为5 m、25 m(或15 m)、50 m、75 m、100 m(或125 m)、150 m和200 m,近岸区域不足200 m的站位取底深以上5 m层次。今生颗石藻采样方法:从CTD采水器各标准层取500 m L海水装入PE采样瓶,立刻加入弱碱性甲醛固定,终浓度1%~2%,置于暗处常温保存。待回到室内实验室后,过滤400~500 mL亚样品到25 mm×0.22μm的混合纤维素膜上,真空泵压力小于100 mm Hg。过滤完毕后,将滤膜放入事先编号的表面皿中风干,剪取一定面积的中央滤膜,采用中性树胶将其固定在载玻片上并封片,待其自然阴干后放入样品盒保存[18—19]。今生颗石藻的镜检工作采用Motic(BA300)偏振光显微镜在1 000×倍率下,观看200~400个视野数不等,依据每个样品的细胞丰度而定,镜检事项参照Bollmann等[20],分类及命名法则参照[21—23]。颗石粒和颗石藻细胞丰度计算方法参见孙军等[19]。
2.3 数据统计分析
今生颗石藻群落指数统计采用香农威纳多样性指数(H′,Shannon-Wiener index),物种均匀度指数(J,Pielou index)和优势度Y,具体计算公式参见孙军
等[24]。
聚类、标序分析采用的颗石藻物种满足出现频率大于30%,单种相对丰度至少有一个站位是大于5%,这样就排除了机会种对群落结构的干扰作用。对筛选出的物种在PRIMER 6.0软件包中计算Bray-Curtis相似性系数,画出相似性矩阵聚类(Cluster)和非参数多维尺度转换标序图(Non-metric multidimensional scaling,MDS),用协强系数(Stress)来检验MDS结果。此后,应用相似性分析(ANOSIM)对各组群之间的结构差异进行统计检验,并根据SIMPER程序分析各物种的贡献率。
解释变量(颗石藻生物群落)与响应变量(环境因子)之间的对应关系在CANOCO 4.5软件中应用约束型排序方法展开分析。生物数据首先经过平方根转化以平衡稀有种权重,然后进行降趋分析(Detrended correspondence analysis,DCA),结果表明最大梯度值小于3,符合线性模型分析标准,合适采用冗余分析(Redundancy analysis,RDA),在开始分析前,应用蒙特卡罗检验(Monte Carlo permutation test)筛选出显著性水平p<0.05的环境因子,最后做出可视化的三序图(站位,物种和环境),分析各因素之间的相关关系。
3 结果
3.1 颗石藻物种组成
调查期间发现今生颗石藻物种34种(表1),优势物种有赫氏艾密里藻(Emilianiahuxleyi)、大洋桥石藻(Gephyrocapsaoceanica)、纤细伞球藻(Umbellosphaeratenuis)、卡特螺旋球藻(Helicosphaeracart-eri)、地中海花冠球藻(Coronosphaeramediterranea)、粗壮环翼球藻(Algirosphaerarobusta)、深水花球藻(Florisphaeraprofunda)和喇叭盘球藻(Discosphaera tubifera)(表2),其中赫氏艾密里藻和大洋桥石藻除了拥有最高的出现频率外,颗石粒相对丰度几乎各占据整体群落的一半,在调查区今生颗石藻群落中占据绝对优势。其他优势物种虽然出现频率也较高,但相对丰度低。

表1 夏季琼东上升流海域今生颗石藻物种种名录Tab.1 Species list of modern coccolithophores in the summer Qiongdong waters
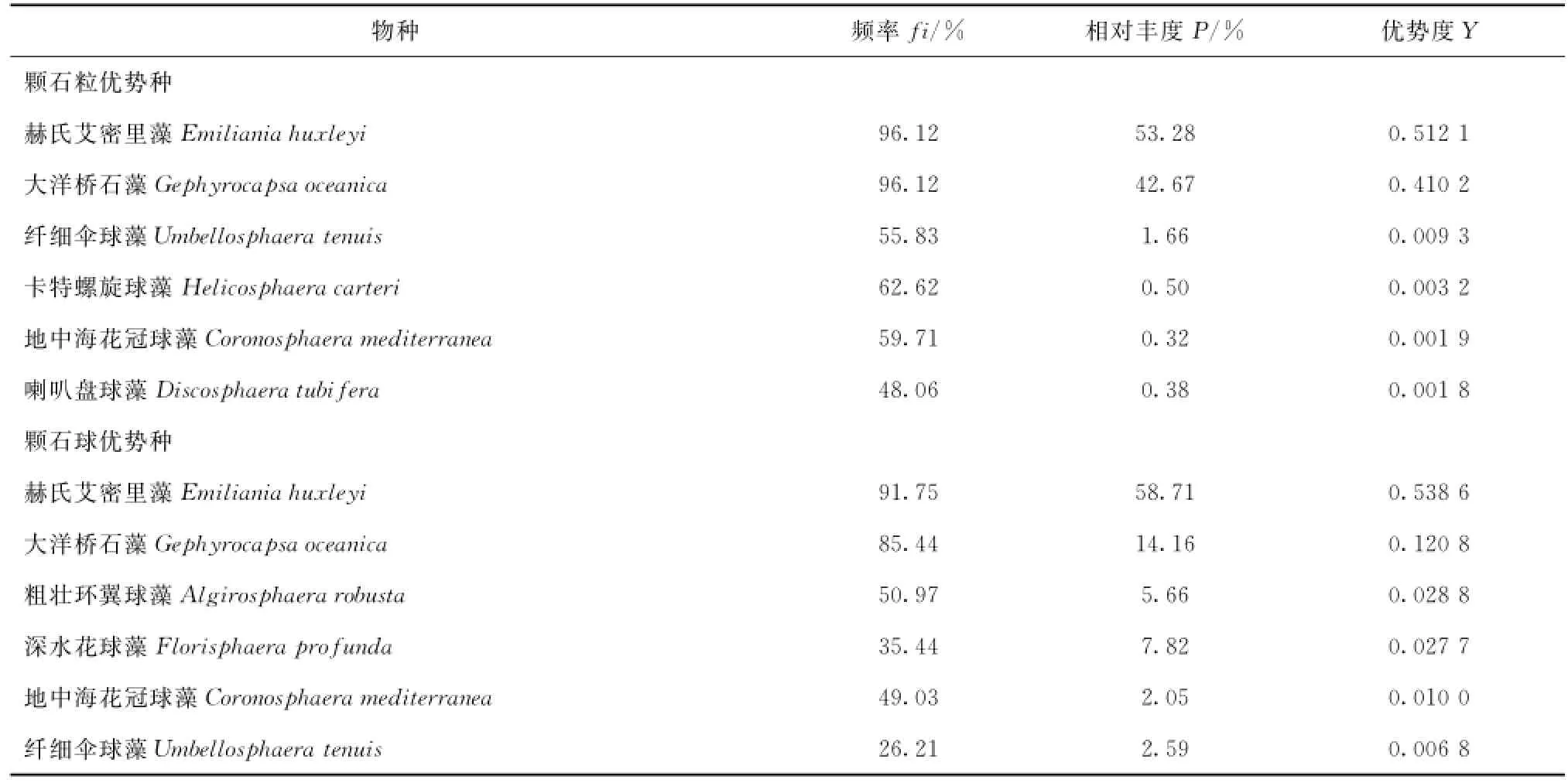
表2 夏季琼东上升流海域今生颗石藻优势物种Tab.2 Common modern coccolithophores in the summer Qiongdong waters
3.2 颗石藻丰度及空间分布
本次调查中颗石粒丰度介于0~2 040.23(平均值:159.27)×103cells/L,颗石球丰度介于0~68.90(平均值:8.10)×103粒/L;表层颗石粒丰度介于0~191.33(平均值:40.29)×103粒/L,颗石球丰度介于0~46.89(平均值,5.01)×103cells/L,与历史资料对比见表2。由数据分析可知颗石粒多集中分布在5 m和75 m水层,因此图2展示了优势种的颗石粒在5 m和75 m层的平面分布(同一物种在两个层次的图例一致),而颗石球在5 m和50 m丰度较高,故图3描绘了颗石球在这两个水层的平面分布。由图2可知,不同优势物种不同层次分布趋势不同,赫氏艾密里藻和大洋桥石藻主要分布在75 m,纤细伞球藻高值出现在SEATS站位表层,地中海花冠球藻分布较为均匀。图3的颗石球高值多集中在50 m,除了纤细伞球藻。
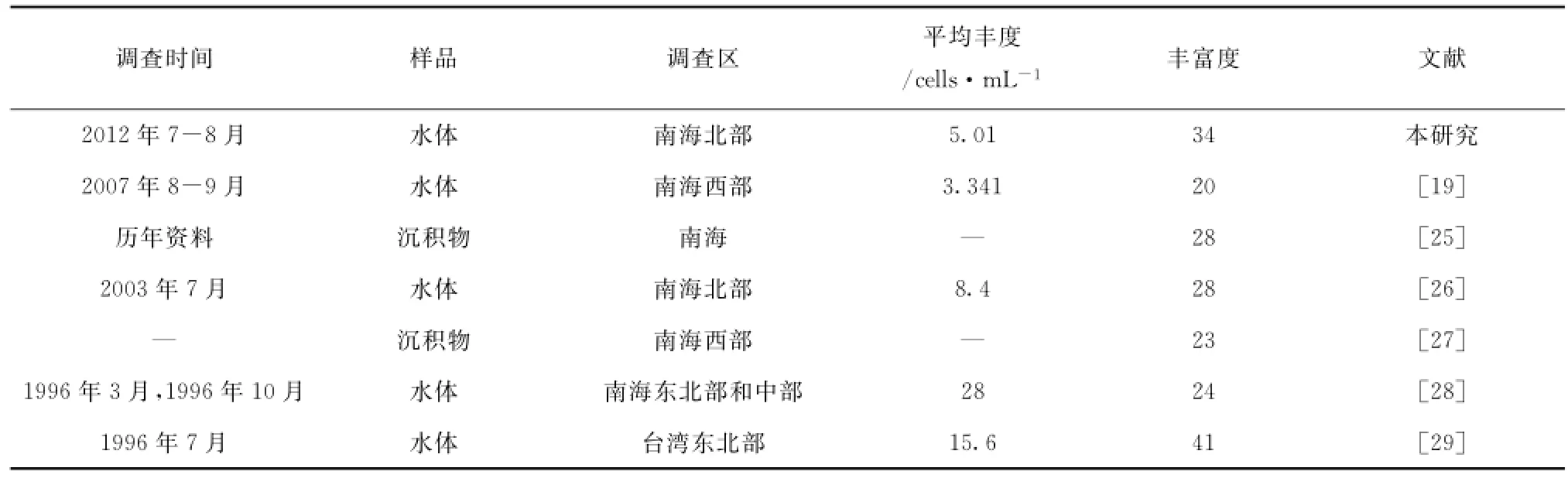
表2 南海海域今生颗石藻历史资料对比Tab.2 Historical data of coccolithophores in the summer Qiongdong waters
图4列举了前6种颗石粒物种在温度-盐度下的点聚图分布,赫氏艾密里藻、大洋桥石藻、卡特螺旋球藻和喇叭盘球藻的丰度高值集中在高盐、低温的上升流水体中(图4a,4b,4d,4f);地中海花冠球藻在整个水体分布较为一致(图4e);纤细伞球藻高值出现在盐度32~34之间的次表层水体(图4f)。从颗石球的空间分布来看(图5),除了纤细伞球藻分布在中部海域的次表层外(图5f),其他优势种在高盐、低温水体中均有高丰度值(图5a~5e)。
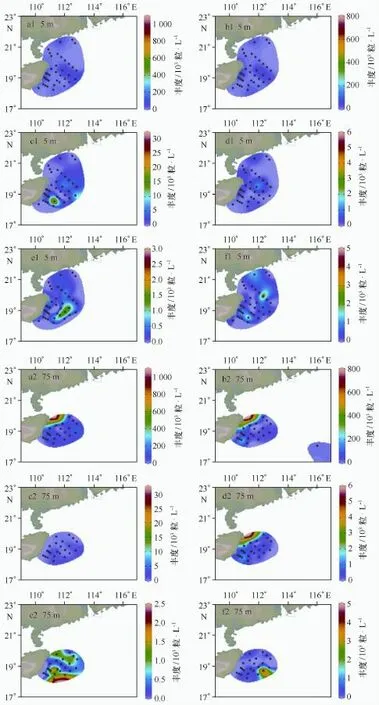
图2 颗石粒丰度在5 m和75 m层的平面分布Fig.2 Distribution of coccolith density at 5 m and 75 m
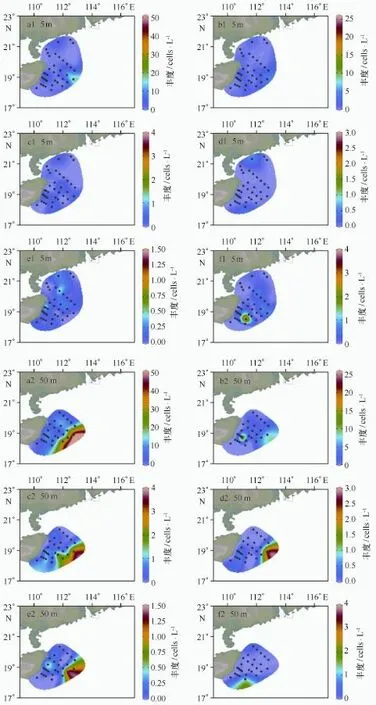
图3 颗石球丰度在5 m和50 m层的平面分布Fig.3 Distribution of coccosphere density at 5 m and 50 m
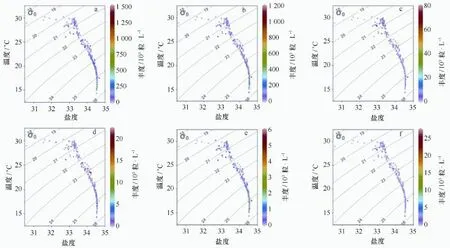
图4 夏季琼东海域颗石粒在温度-盐度(T-S)背景下的分布Fig.4 Coccolith distribution under the temperature-salinity(T-S)properties in the summer Qiongdong water
3.3 多样性指数
对琼东海区今生颗石藻的群落多样性指数研究结果显示,香农威纳多样性指数H′介于0.65~2.85之间,平均值为1.67,物种均匀度指数J介于0.43~1之间,均值为0.76。由图6分布趋势来看,二者呈镶嵌式分布,颗石粒的H′近岸低于外海,H′高值位于海南岛东北部和SEATS站位,J高值出现在湛江附近和海南岛中部水域;颗石球的H′高值则位于海南岛中部和南部水域,J分布与颗石粒J分布趋势类似。
3.4 群落结构描述
基于相似性系数的聚类分析结果如图7所示。从图中可知颗石粒类群在75%的水平上被分为4个组群(GroupⅠ、Ⅱ、Ⅲ、Ⅳ),MDS标序图中协强系数Stress<0.2,认为有一定解释意义[30]。第Ⅰ组包括D002a和DD201a站,位于海南岛南部近岸;第Ⅱ组包含调查区大部分站位,分布比较分散;第Ⅲ组包括QD41、E703、E501、D102a、DD202、D001、D003a等站位;第Ⅳ组只有SEATS站。接下来采用相似性分析检验(ANOSIM)对上述聚类结果进行显著性检验,结果显示GlobalR值等于0.751(p=0.001),大于0.5,说明差异极显著[31],4个组群分离完全。进一步利用相似性百分比分析程序(SIMPER)分析每个组群中各优势种的贡献百分比(由于PRIMER6.0软件对于仅有一个站位的组群无法计算贡献率,因此对GroupⅣ不予讨论),进而分析影响分组的关键种。SIMPER结果表明,颗石粒聚类的GroupⅠ与Ⅲ之间的相异性(average dissimilarity)最高,达到59.35,其他组群之间均小于50%。由表3的物种贡献率(取大于6%为优势)可知,GroupⅠ与Ⅱ的出现共同颗石粒优势种有4个,且赫氏艾密里藻和大洋桥石藻占25%以上,GroupⅡ优势种最多,有8个,GroupⅠ与Ⅲ也有4种共同优势种,且最优势物种是大洋桥石藻,GroupⅢ中优势物种贡献率都偏高,最低的细孔钙盘藻贡献率也高达6.72%。
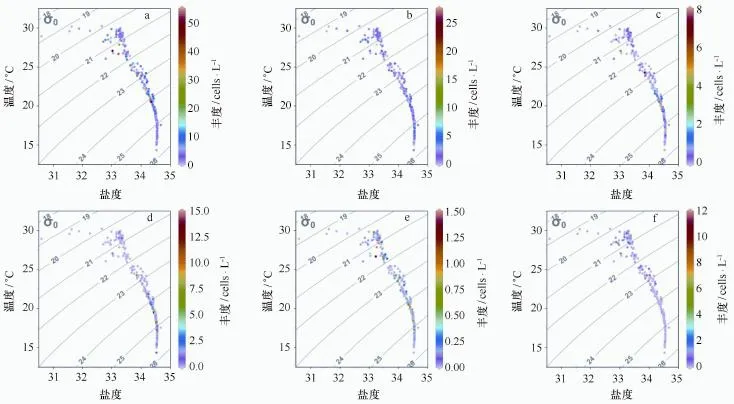
图5 夏季琼东海域颗石球在温度-盐度(T-S)背景下的分布Fig.5 Coccosphere distribution under the temperature-salinity(T-S)properties in the summer Qiongdong water
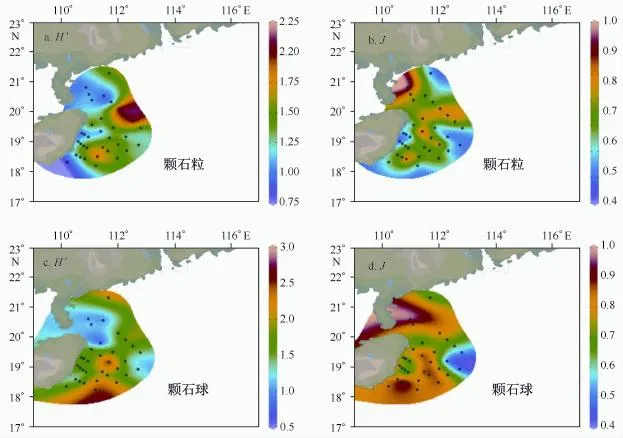
图6 颗石藻表层生物多样性指数(H′)和物种均匀度指数(J)平面分布Fig.6 Surface distribution of Shannon-Wiener index(H′)and Pielou index(J)in summer Qiongdong upwelled water
颗石球在40%的相似水平上可分为5个组群(GroupⅠ、Ⅱ、Ⅲ、Ⅳ、Ⅴ),MDS标序图中协强系数Stress<0.05,说明具有很好的代表性[30]。GroupⅣ和Ⅴ仅由一个站位组成,因此不进行贡献率统计,ANOSIM的结果显示GlobalR值等于0.847(p=0.001),说明差异极显著。从SIMPER程序显示的结果看(表3,颗石球),颗石球优势种较单一,且由赫氏艾密里藻占据主导作用,其他种类贡献率都很低。
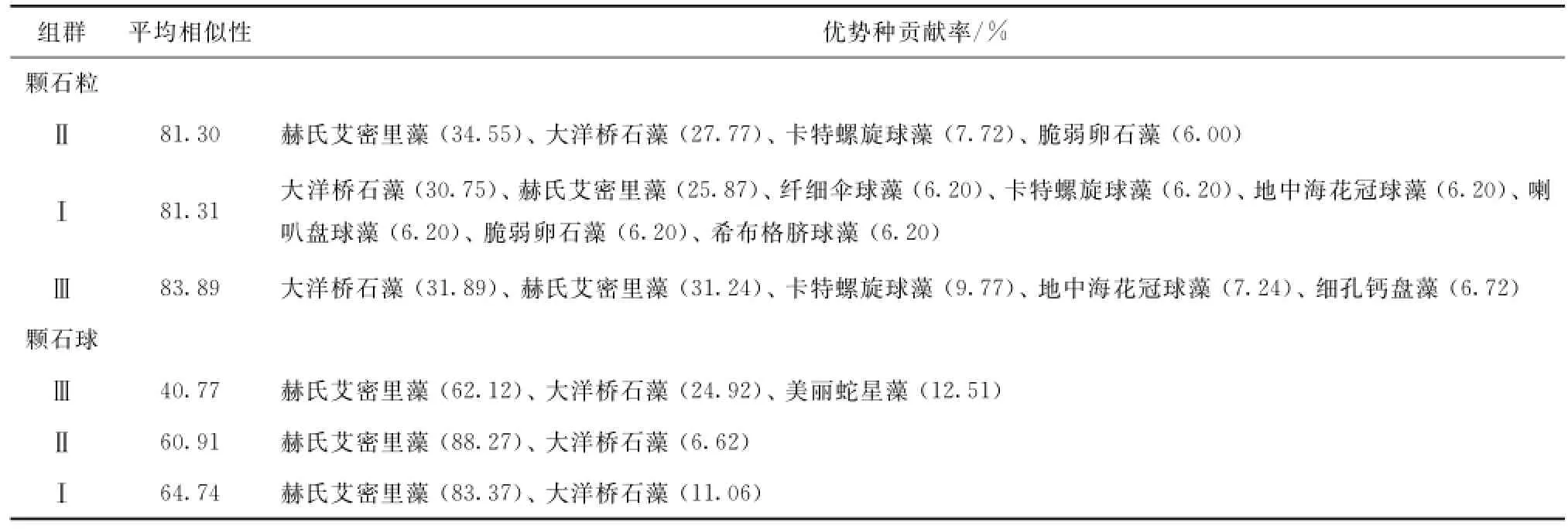
表3 颗石粒优势物种在各个聚类组群中的贡献率Tab.3 Coccolith dominant species percentage contributions to each group
3.5 颗石藻生物群落结构对环境因子的响应
综合应用主成分分析(principle component analysis,PCA)和冗余分析(redundancy analysis,RDA)对影响琼东海区颗石藻群落结构的多重上行效应的环境因子进行排序分析。对颗石藻丰度进行标准化后获得如图8所示的PCA排序结果,颗石粒的前两个主成分加和解释量为48.9%,其中第一轴(PC1)占34.9%,第二轴(PC2)占14%,由图8a颗石粒的PCA二维图可以看出对群落贡献较大的优势物种,贡献极小的非主要成分有不规则伞球藻(U.irregu)、Scyphosphaeraapsteinii(S.apstei)、海洋颗石藻海洋变种(C.pelagi)、贺氏脐球藻(U.hulbur)、翎状角石藻(C.crista)、双片花冠球藻(C.binoda)、贝氏布拉藻(B.bigelo)、Coccolithusbraarudii(C.braaru)和Pontosphaeramultipora(P.multip),大部分主成分多分布在PC1,如赫氏艾密里藻、希布格脐球藻和地中海花冠球藻等,卡特螺旋球藻和纤细伞球藻对PC2相对依赖性大。颗石球的两个主成分轴解释度较高,加和值为72.3%,从图8b可知,贡献较大的物种有扇形剑石藻(G.flabel)、深水花球藻(F.profun)和粗壮环翼球藻(A.robust),与PC2有正相关关系,赫氏艾密里藻(E.huxley)和大洋桥石藻(G.oceani)与PC1也是正相关性,纤细伞球藻(U.tenuis)与PC1呈负相关性。
从PCA排序图中筛选出11个主要颗石粒优势种进行RDA约束型排序分析(图9a),最终筛选出5个解释变量(p<0.05),第一轴(典范轴)对响应变量的解释度为17.2%,能被第一轴很好解释的有赫氏艾密里藻、细孔钙盘藻、玻璃质螺旋球藻(H.hyalina)、大洋桥石藻和卡特螺旋球藻,与PO3-4和N/P呈正相关,并且大部分站位(图中空心圆)的群落结构主要由这些物种构成;溶解氧(DO)与第一轴呈显著正相关,水深(Depth)与第二轴呈显著正相关,亚硝酸盐(NO-2)则与第二轴呈微弱负相关,对物种影响较小,纤细伞球藻与DO和Depth呈正相关性,脆弱卵石藻(O.fragil)和美丽条结藻(S.pulchr)与Depth呈正相关,图中不同物种箭头长度差异大,表明各站位之间的群落结构差异。筛选出9个主要成分的颗石球物种以及满足条件的4种解释变量(p<0.05)进行RDA分析,环境变量之间的角度很小,全部集中在第一轴附近,说明之间的相关性很大,深水花球藻和扇形剑石藻分布较近,与各项营养盐呈正相关关系,美丽条结藻(S.pulchr)和喇叭盘球藻(D.tubife)与各环境因子呈负相关性,赫氏艾密里藻(E.huxleyi)和大洋桥石藻(G.oceani)与第一轴关系较大,与环境因子无明显相关性,地中海花冠球藻(C.medite)、美丽蛇星藻(O.formos)和粗壮环翼球藻(A.robust)与N/P的正相关关系最大。

图7 琼东海域颗石藻群落结构的聚类分析(a1,a2)和非参数多维尺度标序(b1,b2)Fig.7 Cluster analysis(a1,a2)and MDS ordination(b1,b2)of coccolith and coccosphere community structure in Qiongdong upwelling area
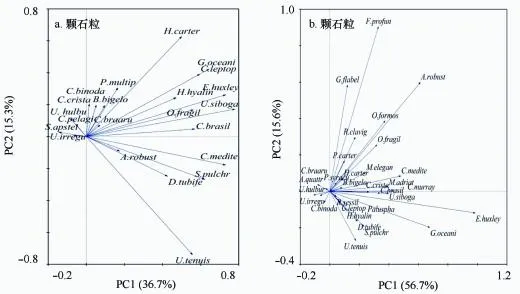
图8 琼东上升流海域颗石藻生物群落组成的主成分分析(PCA)Fig.8 Results of the principal components analysis(PCA)of the coccolithophore community in the upwelling area of Qiongdong
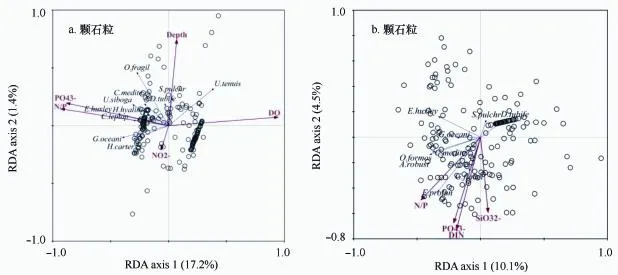
图9 采样站位颗石藻群落组成与环境因子的冗余分析(RDA)图(空心圆代表各站位)Fig.9 Ordination triplot of the redundancy analysis(RDA)of modern coccolithophore data and environmental variables of the studied Qiongdong upwelling area(empty circles represent sampling sites)
4 讨论和结论
本次研究中夏季琼东上升流地区今生颗石藻丰度较近年比差异不大,物种丰富度有所增高(表2),最优势物种是世界性种赫氏艾密里藻和大洋桥石藻,受局地海区环境和气候影响,其他优势种排名有差异,颗石粒和颗石球拥有4个共同优势种,赫氏艾密里藻、大洋桥石藻、纤细伞球藻和地中海花冠球藻。
沿岸上升流为光合自养生物提供丰富的营养物质,大大提高了局部海区的生产力,强上升流区形成了世界著名渔场[12,15]。夏季南海北部陆架上升流区有3个,分别是琼东沿岸上升流、雷州半岛以东的粤西上升流,以及广州湾以南的上升流,其强弱主要受控于风场[13—16,32]。仅选取调查区中部琼东断面E5(E500~E504站位)以及粤西断面E7(E700~E707站位)温度、盐度断面分布来追踪上升流信号(图10),从E5断面可以发现温度、盐度等值线有离岸下倾的趋势,是典型的上升流信号特征,表层受河流、降雨等影响导致上升流信号较弱[32],另外粤西的E7断面上升流信号较弱,一方面是由于粤西沿岸上升流强度一直较琼东弱[32],另一方面现场调查时间在8月9日前后,而上升流最强出现在6、7月[16]。颗石藻作为微型浮游植物的一类,在上升流水体中更易生长繁殖[33],然而不同物种对上升流信号响应结果不同,在图4、图5的优势种的点聚图中可知,大部分颗石藻分布在明显的低温、高盐、高密的上升流信号较强的水体,而图4c,5f中的纤细伞球藻高值分布在高温低盐的非上升流水体中。从大多数物种的平面分布(图2,图3)看,海南岛以东和雷州半岛东部出现物种丰度高值区。群落多样性指数和均匀度指数分布趋势在雷州半岛东部的上升流域呈镶嵌特征(图6),原因在于物种丰富度(species richness)的影响,此处H′并不高,因其物种丰富度低,故均匀度J很高,琼东上升流域态势与之相反,H′和J的数值均很高。这说明上升流除了有季节性外,还有一定的局地性,生态系统特征及生物地球化学过程不尽相同,比如琼东水域地形及流态较复杂,备受黑潮等水团入侵的影响[13]。
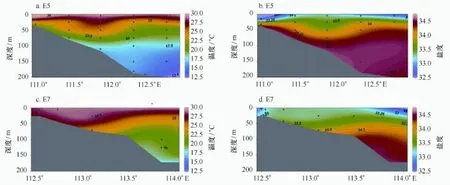
图10 E5和E7断面的温度、盐度垂直剖面分布Fig.10 Transectional pattern of temperature and salinity of E5 and E7 section
颗石藻等浮游植物生物群落结构受控于海区物理、化学、水文和其他生物因素,在这些因素的综合影响下,生物分布呈一定的斑块状,探讨生物因子对物理环境因子的响应特征、规律很有必要。首先将颗石藻生物数据典型水层进行聚类和多维定标分析,两种组合图形更好地展现颗石藻生物种群空间结构。各个站位的颗石粒和颗石球分布特征及组成不同,因此分别分成了四类和五类组群,相似性分析(ANOSIM)的统计检验结果GlobalR值均高于0.5,表明有显著差异。进一步通过PCA筛选出主要成分的生物因子,结合海区物理化学环境因子进行整体综合分析,RDA结果显示N/P、PO3-4、NO-2、DO和Depth决定了颗石粒群落组成与分布,而影响颗石球群落结构的主要影响因素有N/P、PO3-4、SiO2-3和DIN(NO-2和NO-3),RDA分析结果的典范轴解释度不高,一方面由于解释变量经过显著性的筛选,符合条件的因子少,另一方面也说明颗石藻生物群落结构的复杂性,时空调控机理复杂,影响因素多元化,除了上行主导作用外,下行效应因子也有一定影响,但仍可以解释一定的分布规律。为进一步探明琼东上升流区今生颗石藻的生物多样性规律及生态分布特征既有其独立性,也与其他上升流有关联性,因此未来研究亟需加大南海更大的时空尺度调查,综合对比分析其他地区上升流及生物耦合特征,发现琼东上升流区今生颗石藻群落的演替规律及对未来环境的响应趋势。
致谢:感谢厦门大学戴民汉和胡建宇教授提供营养盐及各项水文参数,感谢郭术津博士协助现场采样工作,“东方红2”号科考船提供作业平台,谨致谢忱!
[1] Moheimani N R,Webb J P,Borowitzka M A.Bioremediation and other potential applications of coccolithophorid algae:a review[J].Algal Re-search,2012,1(2):120-133.
[2] Bauer JE,Cai W J,Raymond P A,et al.The changing carbon cycle of the coastal ocean[J].Nature,2013,504(7478):61-70.
[3] 孙军.今生颗石藻的有机碳泵和碳酸盐反向泵[J].地球科学进展,2007,22(12):1231-1239.
Sun Jun.Organic carbon pump and carbonate counter pump of living coccolithophorid[J].Advances in Earth Science,2007,22(12):1231-1239.
[4] Alvain S,Moulin C,Dandonneau Y,et al.Seasonal distribution and succession of dominant phytoplankton groups in the global ocean:a satellite view[J].Global Biogeochemical Cycles,2008,22(3),doi:10.1029/2007GB003154.
[5] Sun J,Gu X Y,Feng Y Y,et al.Summer and winter living coccolithophores in the Yellow Sea and the East China Sea[J].Biogeosciences,2014,11:779-806.
[6] Richier S,Kerros M E,de Vargas C,et al.Light-dependent transcriptional regulation of genes of biogeochemical interest in the diploid and haploid life cycle stages ofEmilianiahuxleyi[J].Applied and Environmental Microbiology,2009,75(10):3366-3369.
[7] Cao Z M,Dai M H.Shallow-depth CaCO3dissolution:evidence from excess calcium in the South China Sea and its export to the Pacific Ocean[J].Global Biogeochemical Cycles,2011,25(2),doi:10.1029/2009GB003690.
[8] Bown P R,Gibbs SJ,Sheward R,et al.Searching for cells:the potential of fossil coccospheres in coccolithophore research[J].Journal of Nannoplankton Research,2014,34:5-21.
[9] Hagino K,Young J R,Bown P R,et al.Re-discovery of a“living fossil”coccolithophore from the coastal waters of Japan and Croatia[J].Marine Micropaleontology,2015,116:28-37.
[10] Zhang C H,Xi X L,Liu S T,et al.A mesoscale eddy detection method of specific intensity and scale from SSH image in the South China Sea and the Northwest Pacific[J].Science China Earth Sciences,2014,57(8):1897-1906.
[11] Hu Z F,Tan Y H,Song X Y,et al.Influenceof mesoscaleeddies on primary production in the South China Sea during spring inter-monsoon period[J].Acta Oceanologica Sinica,2014,33(3):118-128.
[12] Jing Z Y,Qi Y Q,Du Y.Upwellingin the continental shelf of northern South China Sea associated with 1997-1998 El Niño[J].Journal of Geophysical Research,2011,116(C2),doi:10.1029/2010JC006598.
[13] 经志友,齐义泉,华祖林.南海北部陆架区夏季上升流数值研究[J].热带海洋学报,2008,27(3):1-8.
Jing Zhiyou,Qi Yiquan,Hua Zulin.Numerical study on summer upwelling over northern continental shelf of South China Sea[J].Journal of Tropical Oceanography,2008,27(3):1-8.
[14] 颜廷壮.浙江和琼东沿岸上升流的成因分析[J].海洋学报,1992,14(3):12-18.
Yan Tingzhuang.A diagnostic analysis on Zhejiang and Qiongdong coastal upwelling[J].Haiyang Xuebao,1992,14(3):12-18.
[15] 韩舞鹰,王明彪,马克美.我国夏季最低表层水温海区——琼东沿岸上升流区的研究[J].海洋与湖沼,1990,21(3):267-275.
Han Wuying,Wang Mingbiao,Ma Kemei.On the lowest surface water temperature area of China Sea in summer——the upwelling along the east coast of hainan Island[J].Oceanologia et Limnologia Sinica,1990,21(3):267-275.
[16] 谢玲玲张书文,赵辉.琼东上升流研究概述[J].热带海洋学报,2012,31(4):35-41.
Xie Lingling,Zhang Shuwen,Zhao Hui.Overview of studies on Qiongdong upwelling[J].Journal of Tropical Oceanography,2012,31(4):35-41.
[17] Cao Z M,Dai M H,Zheng N,et al.Dynamics of the carbonate system in a large continental shelf system under the influence of both a river plume and coastal upwelling[J].Journal of Geophysical Research,2011,116(G2),doi:10.1029/2010JG001596.
[18] 靳少非,孙军,刘志亮.2010秋季东海今生颗石藻的空间分布[J].生态学报,2013,33(1):120-131.
Jin Shaofei,Sun Jun,Liu Zhiliang.The distribution of living coccolithophore in East China Sea in autumn,2010[J].Acta Ecologica Sinica,2013,33(1):120-131.
[19] 孙军,安佰正,戴民汉,等.夏季南海西部今生颗石藻[J].海洋与湖沼,2011,42(2):170-178.
Sun Jun,An Baizheng,Dai Minhan,et al.Living coccolithophores in the western South China Sea in summer 2007[J].Oceanologia et Limnologia Sinica,2011,42(2):170-178.
[20] Bollmann J,Cortés M Y,Haidar A T,et al.Techniques for quantitative analyses of calcareous marine phytoplankton[J].Marine Micropaleontology,2002,44(3/4):163-185.
[21] Jordan R W,Cros L,Young J R.A revised classification scheme for living haptophytes[J].Micropaleontology,2004,50(S1):55-79.
[22] Frada M,Young J,Cachão M,et al.A guide to extant coccolithophores(Calcihaptophycidae,Haptophyta)using light microscopy[J].Journal of Nannoplankton Research,2010,31(2):58-112.
[23] 孙军,靳少非.中国近海今生颗石藻物种多样性初步研究[J].生物多样性,2011,19(6):787-797.
Sun Jun,Jin Shaofei.Species diversity of living coccolithophores in Chinese sea waters[J].Biodiversity Science,2011,19(6):787-797.
[24] 孙军,刘东艳.多样性指数在海洋浮游植物研究中的应用[J].海洋学报,2004,26(1):62-75.Sun Jun,Liu Dongyan.The application of diversity indices in marine phytoplankton studies[J].Haiyang Xuebao,2004,26(1):62-75.
[25] 王勇军,陈木宏,陆钧,等.南海表层沉积物中钙质超微化石分布特征[J].热带海洋学报,2007,26(5):26-34.
Wang Yongjun,Chen Muhong,Lu Jun,et al.Distribution of calcareous nannofossils in surface sediments of South China Sea[J].Journal of Tropical Oceanography,2007,26(5):26-34.
[26] Lee Chen Y L,Chen H Y,Chung C W.Seasonal variability of coccolithophore abundance and assemblage in the northern South China Sea[J].Deep-Sea Research PartⅡ:Topical Studies in Oceanography,2007,54(14/15):1617-1633.
[27] 陈芳,黄永样,段威武,等.南海西部表层沉积中的钙质超微化石[J].海洋地质与第四纪地质,2002,22(3):35-40.
Chen Fang,Huang Yongyang,Duan Weiwu,et al.Calcareous nannoplankton in surface sediments in the west of the South China Sea[J].Marine Geology&Quaternary Geology,2002,22(3):35-40.
[28] Yang T N,Wei K Y,Chen L L.Occurrence of coccolithophorids in the northeastern and central South China Sea[J].Taiwania,2003,48(1):29-45.
[29] Yang T N,Wei K Y,Gong G C.Distribution of coccolithophorids and coccoliths in surface ocean off northeastern Taiwan[J].Botantical Bulletin of Academia Sinica,2001,42:287-302.
[30] 陈小庆,陈斌,黄备,等.夏季舟山渔场及邻近海域浮游动物群落结构特征分析[J].动物学研究,2010,31(1):99-107.
Chen Xiaoqing,Chen Bin,Huang Bei,et al.Analysis on community structure of zooplankton in Zhoushan fishing ground and its adjacent area in summer[J].Zoological Research,2010,31(1):99-107.
[31] 吴荣军,李瑞香,朱明远,等.应用PRIMER软件进行浮游植物群落结构的多元统计分析[J].海洋与湖沼,2006,37(4):316-321.
Wu Rongjun,Li Ruixiang,Zhu Mingyuan,et al.Multivari ateanalysis with primer on marine phytoplankton community structure in mesocosm system[J].Oceanologia et Limnologia Sinica,2006,37(4):316-321.
[32] 许金电,蔡尚湛,宣莉莉,等.2006年夏季琼东、粤西沿岸上升流研究[J].海洋学报,2013,35(4):11-18.
Xu Jindian,Cai Shangzhan,Xuan Lili,et al.Study on coastal upwelling in eastern Hainan Island and western Guangdong in summer,2006[J].Haiyang Xuebao,2013,35(4):11-18.
[33] Silva A,Palma S,Moita M T.Coccolithophores in the upwelling waters of Portugal:four years of weekly distribution in Lisbon bay[J].Continental Shelf Research,2008,28(18):2601-2613.
Study on modern coccolithophores in coastal region along the east Hainan Island
Liu Haijiao1,2,Sun Jun1,2,Feng Yuanyuan1,2
(1.College of Marine and Environmental Sciences,Tianjin University of Science and Technology,Tianjin 300457,China;2.Tianjin Key Laboratory of Marine Resources and Chemistry,Tianjin University of Science and Technology,Tianjin 300457,China)
Multidisciplinary investigations of modern coccolithophores community in the Qiongdong upwelling water of the northern South China Sea(SCS)were performed during the summer cruise of the year 2012.The present study reported the coccolithophores community structure and distribution feature.Total 34 species were identified under polarizing microscope by the filtered membrane samples.The preponderant species wereEmilianiahuxleyi,Gephyrocapsaoceanica,Umbellosphaeratenuis,Helicosphaeracarteri,Coronosphaeramediterranea.Coccolith abundance ranged within 0-2 040.23×103cells/L,with an average value of 8.10×103cells/L;and coccosphere abundance was 0-68.90×103cells/L,with an average value of 5.01×103cells/L.majority of species distributed in the strongly upwelled water.The central area of Qiongdong had largerH′value than that of east of Leizhou Peninsula.TheH′andJvalue of coccolith exhibited mosaic pattern.The results of cluster and MDS showed that coccolith and coccosphere were classified into four groups and five groups respectively,and ANOSIM test proved it to be significant.The redundancy analysis(RDA)indicated that N/P,PO3-4,NO-2and SiO2-3are the major controlling factors influencing the modern coccolithophore community.
coccolithophore;community structure;dominant species;Qiongdong upwelling;cluster analysis;redundancy analysis
Q948.8
A
0253-4193(2015)12-0027-14
刘海娇,孙军,冯媛媛.琼东海域今生颗石藻群落研究[J].海洋学报,2015,37(12):27—40,
10.3969/j.issn.0253-4193.2015.12.004
Liu Haijiao,Sun Jun,Feng Yuanyuan.Study on modern coccolithophores in coastal region along the east Hainan Island[J].Haiyang Xuebao,2015,37(12):27—40,doi:10.3969/j.issn.0253-4193.2015.12.004
2015-05-18;
2015-10-08。
新世纪优秀人才支持计划(NCET-12-1065);国家重点基础研究发展计划(2015CB954002);国家自然科学基金(41176136,41276124,41306118)。
刘海娇(1990—),女,天津市人,主要从事浮游植物生态学研究。
*通信作者:孙军,教授,主要从事海洋生态学研究。E-mail:phytoplankton@163.com
