考虑生活史的太湖沉水植物优势种遥感监测*
2015-05-10周兴东罗菊花
王 琪,周兴东,罗菊花,陈 冲
(1:江苏师范大学城建与环境学部,徐州 221000)
(2:中国科学院南京地理与湖泊研究所,南京 210008)
考虑生活史的太湖沉水植物优势种遥感监测*
王 琪1,周兴东1,罗菊花2,陈 冲1**
(1:江苏师范大学城建与环境学部,徐州 221000)
(2:中国科学院南京地理与湖泊研究所,南京 210008)
水生植物是浅水湖泊的重要类群,也是湖泊环境变化的指示物,快速监测水生植物的时空分布对湖泊生态修复和管理具有重要的指导意义.基于多时相环境星影像,构建太湖水生植物分类决策树模型,将太湖水生植物分成挺水、浮叶和沉水植物3种类别,结合沉水植物优势种生活史特征,提出了一种考虑生活史信息的太湖沉水植物优势种遥感监测方法,并对太湖7大沉水植物优势种进行时空监测.通过2013年7、8和9月野外调查样点验证,3期遥感影像分类后总的精度分别为83.04%、81.82%和85.47%,2013年太湖沉水植物优势种总体识别精度为62.20%.提出的水生植物遥感分类及沉水植物优势种识别方法为太湖管理部门开展水草打捞和生态修复提供依据和参考,同时为研究太湖沉水植物的历史变迁奠定基础.
水生植物;太湖;遥感;优势种;生活史;决策树
近年来浅水湖泊的生态修复成为水环境保护的工作热点.研究表明,浅水湖泊生态系统退化的重要标志之一是大型水生植物的消失,尤其是沉水植物[1].生物修复技术是改善水体富营养化状态的有效方式之一,不同水生植物生长体系对水体中的磷、氮及有机物的吸收能力各有差别,进而对水体的净化能力也有较大差异[2].因此,大面积、准确、快速地监测浅水湖泊水生植物的时空分布对湖泊管理和藻华治理具有重要意义.
传统的水生植物监测方法仍然是人工调查法,该方法费时、费力,代表性差,且很多水生植物生长的地方人难以到达.遥感技术能够快速获取大面积连续地物信息,故在水生植物时空分布信息监测方面具有传统方法不可比拟的优势.早期一些学者就基于航空照片开展了水生植物的研究,如Valta-Hulkkonen等[3]利用数字假彩色航拍照片对湖泊中几种水生植物优势种进行识别;Zhu等[4]利用航空相片对1972、1980和1999-2002年的北美安大略湖沉水植物的空间分布和覆盖度进行定量化监测.近年来国内外很多学者试图利用卫星遥感数据监测水生植物的时空分布,并取得了一些成果,如Zhao等[5]提出基于Landsat ETM+数据的光谱指数阈值分类树模型用于太湖水生植物的分类;Ma等[6]在太湖水生植物的遥感提取中,考虑到水体中的悬浮物和藻类等与沉水植物容易产生“异物同谱”现象,使得大面积的水体被误分为沉水植物,将水体透明度和水深的先验知识引入分类决策树,较好地消除了分类过程的“异物同谱”现象,较准确地提取了太湖的水生植物.
对于沉水植物优势种遥感识别研究,大都基于地面高光谱遥感数据研究不同沉水植物优势种的光谱特征.如Yuan等[7]利用地面实测的高光谱遥感数据研究不同覆盖度的沉水植物旋叶苦草的光谱特征;Han等[8]利用便携式光谱仪研究不同深度、不同透明度及藻类水体中沉水植物的光谱响应特征;袁琳等[9]利用地面实测的高光谱遥感数据对上海崇明国际湿地公园人工湖中不同盖度的沉水植物穗花狐尾藻和苦草的反射光谱进行研究;李俊生等[10]利用地面实测的光谱数据,通过4种光谱指数建立了水华、浮叶植物、沉水植物和水体的判别公式.综上,虽然国内外在沉水植物光谱研究方面取得了一些进展,但对基于卫星遥感数据的大面积沉水植物优势种遥感识别研究还鲜有报道.
本研究针对太湖沉水植物优势种的大面积遥感识别,获取多时相环境星(HJ-CCD)影像数据,构建太湖水生植物分类决策树模型,利用GIS技术提取HJ-CCD影像中的沉水植物,在去除梅梁湾、竺山湾和西太湖蓝藻水华的前提下,结合沉水植物优势种的生活史信息,提出考虑生活史信息的太湖沉水植物优势种遥感监测方法,对太湖水生植物及沉水植物优势种的空间分布进行研究.
1 研究区概况与数据源
1.1 研究区概况
太湖(30°55′40″~31°32′58″N,119°52′32″~120°36′10″E)是我国第3大淡水湖泊,位于长江三角洲南翼坦荡的太湖平原上,平均水深只有1.9m,是典型的浅水湖泊,同时也是我国3大富营养化湖泊之一.为了便于本研究对沉水植物监测结果的定性描述和定量分析,参考已有资料和文献[11],初步将太湖划分为9大区,包括竺山湾、梅梁湾、贡湖湾、湖心区、南太湖、东太湖、胥口和东山湾、镇湖和光福湾、西山岛附近.多年观测结果显示,湖心区、竺山湾和梅梁湾水体中含有高浓度的藻类和悬浮物质;东太湖存在大规模河蟹生态围网养殖区;每年夏季来临,胥口湾、东太湖航道及两侧分布大量的水生植被,太湖环保部门清理任务艰巨.
1.2 遥感数据源及预处理
本研究采用HJ-CCD遥感数据,包括蓝、绿、红(波长分别为0.43~0.52、0.52~0.60、0.63~0.69μm)3个可见光波段和1个近红外波段(波长为0.76~0.90μm),空间分辨率为30m.本研究共计获取了2013年11景无云状况下的影像,获取时间分别为:1月18日、2月20日、3月12日、4月25日、5月24日、7月11日、8月16日、9月26日、10月28日、11月17日和12月27日.通过ENVI软件对环境星影像进行辐射校正(辐射定标和大气校正),消除一切与辐射有关的误差.此外,利用精校正过的覆盖太湖的Landsat TM 8影像对所有影像开展几何校正,其校正误差控制在0.5个像元之内.
1.3 野外样点获取
根据HJ-CCD影像的获取时间,分别于2013年开展3次同步野外实验,调查时间分别为7月11-13日,8月15-21日及9月23-26日,共设置了400多个样点(图1),测定项目包括水生植物优势种的类型、盖度、透明度和位置等.
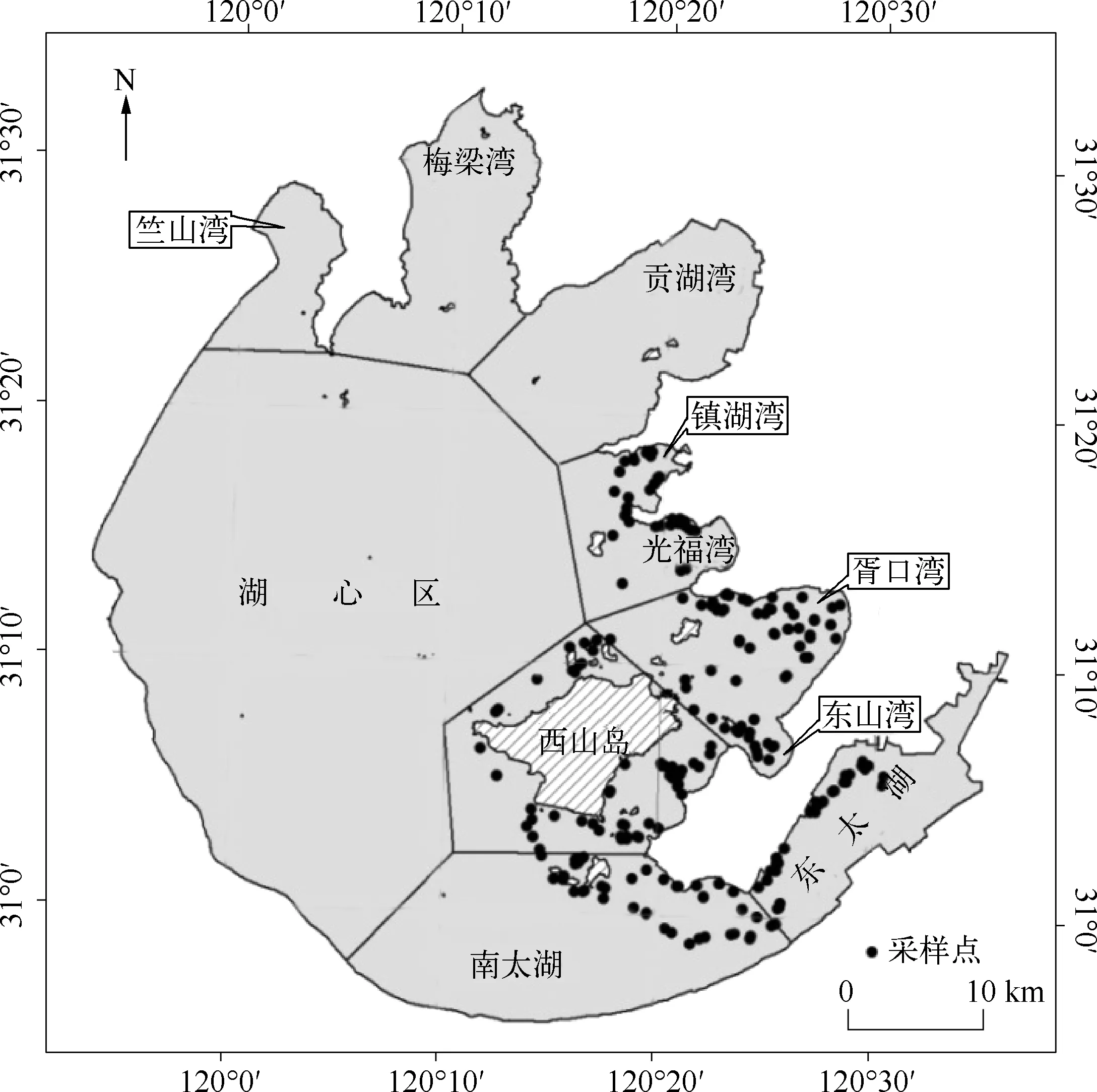
图1 研究区调查点的空间分布Fig.1 Study area and distribution of sampling sites
2 研究方法
2.1 沉水植物优势种生活史信息提取
2013年野外实地调查发现,太湖主要分布3种植物类型,分别为挺水、浮叶和沉水植物.挺水植物主要是芦苇,多分布在岸边的浅水域;浮叶植物中荇菜最为常见,主要分布在胥口湾和西山岛东部的航道两侧,南太湖、光福湾和镇湖湾有零星分布;沉水植物种类繁多,优势种共有7种.本研究首先提取了太湖沉水植物优势种的生活史信息(表1).7大优势种的生活史信息分别为:(1) 马来眼子菜(Potamogetonmalaianus):不能越冬生长,每年12月到次年2月进入越冬期,3月份进入萌发期,4-6月进入缓慢生长期,7-9月进入高速生长期,10-11月生长速度减缓并逐渐枯萎死亡,在10月下旬生物量达到最大值[12-14];(2) 伊乐藻(Elodeanuttalli):能够忍受0℃以下的寒冷,在冬季以营养体越冬,5-6月进入高速生长期,在6月下旬到7月上旬伊乐藻的生物量达到最大值,随后生长随高温停滞,到了秋季再度生长,形成了新的种群[15-17];(3) 苦草(Vallisnerianatans):不能越冬生长,3月进入萌发期,4-6月进入缓慢生长期,7-9月份进入高速生长期,在10月上旬生物量达到最大值[18-19];(4) 微齿眼子菜(Potamogetonmaackianus):不能越冬生长,4月上旬开始大量萌生,在7月份生物量达到最大值,随后生长速度减缓并逐渐枯萎死亡[20];(5) 金鱼藻(Ceratophyllumdemersum):不能越冬生长,4-5月份进入缓慢生长期,6-8月份进入高速生长期,在8月下旬生物量达到最大值,随后生长速度减缓并逐渐枯萎死亡[21,23];(6) 穗花狐尾藻(Myriophyllumspicatum):不能越冬生长,4-6月份进入高速生长期,在7月份生物量达到最大值,8-9月份生长速度减缓并逐渐枯萎,在10月份其生物量达到最小值[22-23];(7) 菹草(Potamogetoncrispus):较为耐寒,能够忍受0℃以下的寒冷,3-4月份进入高速生长期,在4-5月份生物量达到最大值,随后很快枯萎死亡,在7月份其生物量达到最小值,8-10月份进入休眠状态,11月份进入萌发期,12月至次年2月份进入缓慢生长期[24].
表1 太湖沉水植物优势种生活史信息*
Tab.1 Living history of dominant species of submerged vegetation in Lake Taihu
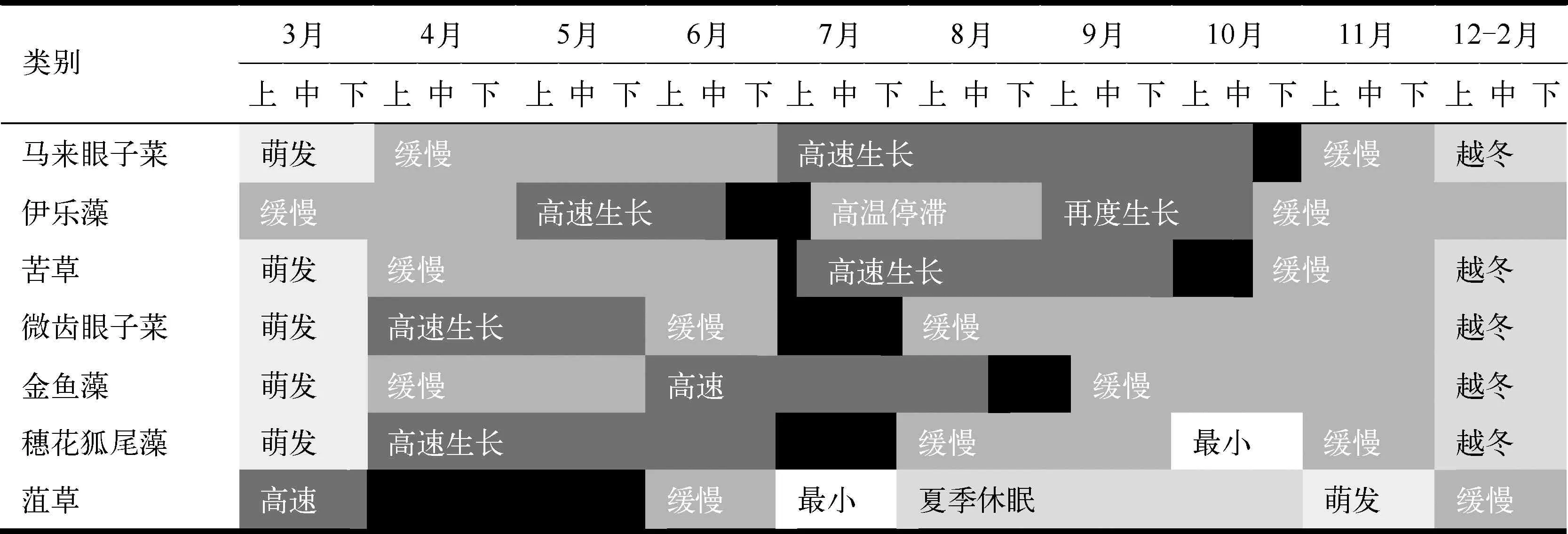
*黑色代表生物量达到最大值,白色代表生物量达到最小值.
2.2 水生植物决策树分类模型构建
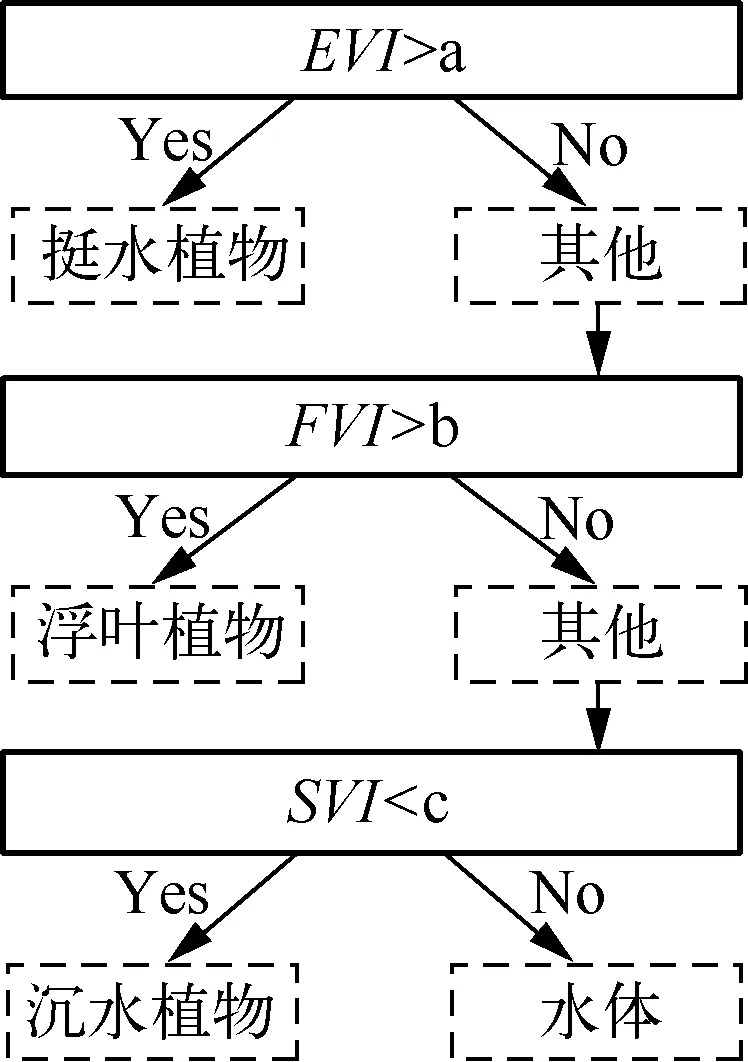
图2 基于CART算法构建的决策树(a、b、c为阈值)Fig.2 The decision tree based on CART
2.3 去除蓝藻水华对沉水植物准确监测的影响
水生植物遥感监测研究表明,蓝藻水华和悬浮物浓度较高的水体与生长沉水植物的水体光谱特征相似,容易形成“异物同谱”现象,从而造成不同地物的混淆,严重降低沉水植物的提取精度[6].根据大量的监测研究和实测调查,太湖梅梁湾、竺山湾和湖心区是蓝藻水华的频发区[27],且除了梅梁湾有少量菹草分布外[28],竺山湾和湖心区基本无沉水植物分布,本研究将竺山湾和湖心区的沉水植物遥感分类结果划为藻华频发区进行掩膜,不参与沉水植物优势种的分类.针对梅梁湾藻华区的提取,中国科学院南京地理与湖泊研究所湖泊流域科学数据共享平台的蓝藻水华监测报告显示,2013年3月12日和5月24日梅梁湾没有监测到蓝藻水华,而3月和5月正逢菹草的繁殖旺季,此时梅梁湾沉水植物遥感分类结果正是菹草的分布范围,再结合剩余9个月梅梁湾沉水植物遥感分类结果来划定2013年太湖梅梁湾的藻华分布区(图3).
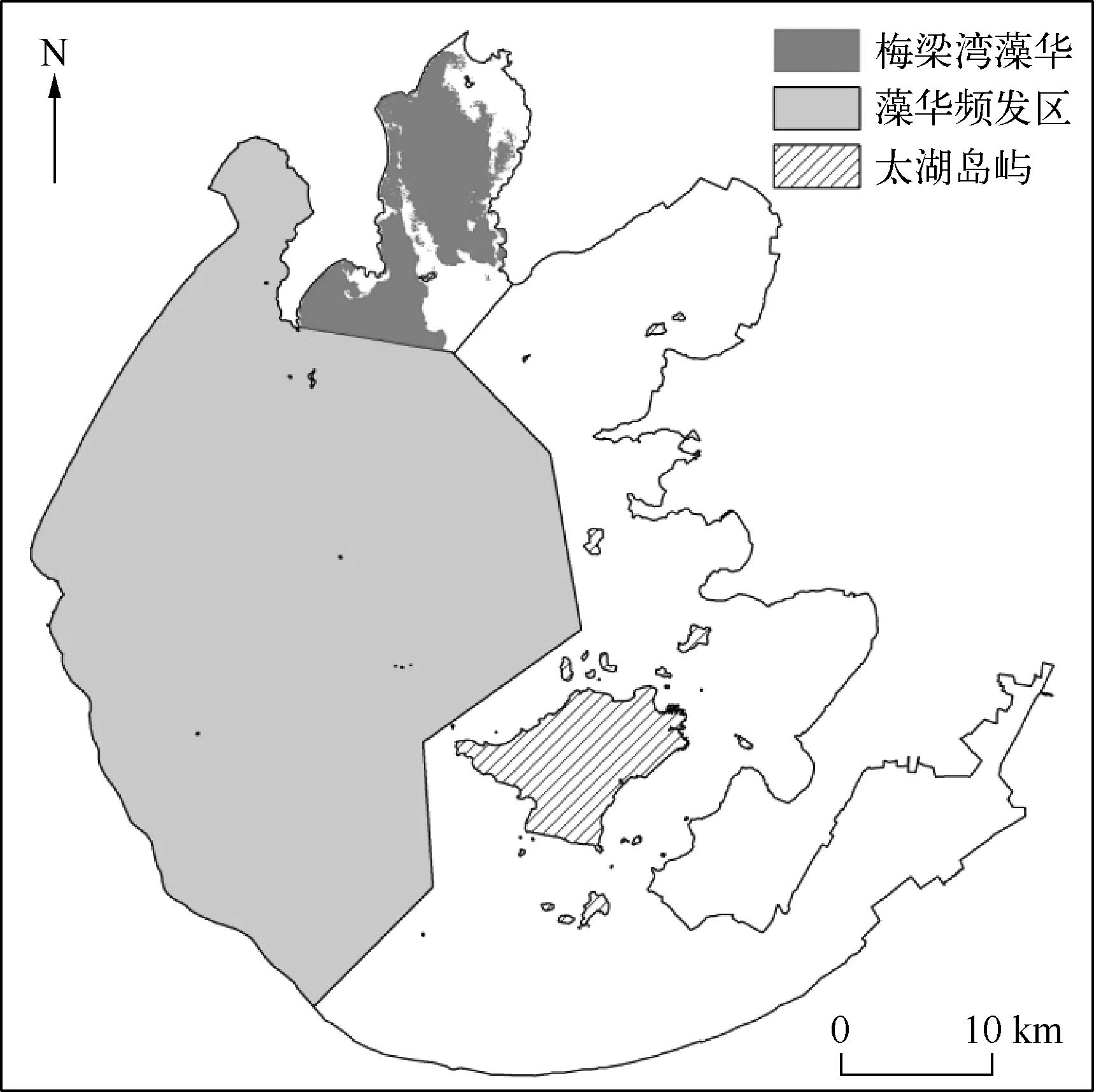
图3 太湖藻华频发区Fig.3 Location of cyanobacteria blooms in Lake Taihu
2.4 沉水植物优势种分类的方法
本研究借助GIS技术,提取了2013年11个月HJ-CCD影像中的沉水植物,在去除湖心区、竺山湾和梅梁湾蓝藻水华的基础上,提出了考虑生活史的太湖沉水植物优势种分类方法(图4),在分类的过程中,多次使用ArcGIS空间叠加分析中的擦除与合并功能,具体操作流程为:(1) 马来眼子菜、微齿眼子菜、穗花狐尾藻、金鱼藻和苦草为喜温类沉水植物,在0℃以下基本消亡,伊乐藻和菹草可以在0℃以下继续生长繁殖,冬季1月最低气温低于0℃,此时太湖主要分布的沉水植物是伊乐藻和菹草,7月菹草的生物量达到最小值[24],而此时伊乐藻的生物量达到最大值[15],7月的沉水植物图层(伊乐藻较多)擦除1月的沉水植物图层(菹草和伊乐藻)得到菹草图层,菹草图层擦除1月的沉水植物图层得到伊乐藻图层;(2) 在7月穗花狐尾藻生物量达到最大值,10月其生物量相对最低[23],10月的沉水植物图层(穗花狐尾藻较少)擦除7月沉水植物图层(穗花狐尾藻最多)得到穗花狐尾藻+伊乐藻+菹草图层,通过擦除操作去除已经分类出的伊乐藻+菹草图层得到穗花狐尾藻图层;(3) 微齿眼子菜在4-5月大量萌生[20],3月沉水植物图层(微齿眼子菜较少)擦除5月沉水植物图层(微齿眼子菜较多)得到微齿眼子菜+伊乐藻+菹草+穗花狐尾藻图层,通过擦除操作去除已经分类出的伊乐藻+菹草+穗花狐尾藻图层,得到微齿眼子菜图层;(4) 金鱼藻在8月下旬到9月初达到最大生物量,随后很快消亡[23],10月沉水植物图层(金鱼藻较少)擦除8月沉水植物图层(金鱼藻最多),得到金鱼藻+伊乐藻+菹草+穗花狐尾藻+微齿眼子菜图层,通过擦除操作去除已经分类出的伊乐藻+菹草+穗花狐尾藻+微齿眼子菜图层得到金鱼藻图层;(5) 分别合并11个月份和前4步提取的5种沉水植物图层,继而用菹草+伊乐藻+穗花狐尾藻+微齿眼子菜+金鱼藻图层擦除总沉水植物图层得到马来眼子菜+苦草图层,马来眼子菜在10月下旬达到最大生物量[15],10月下旬的沉水植物图层擦除马来眼子菜+苦草图层得到苦草图层,苦草图层擦除马来眼子菜+苦草图层得到马来眼子菜图层.
图4 沉水植物优势种分类流程(L1~L5和L6~L11分别为1-5月和7-12月沉水植物矢量图层)Fig.4 Classification process of dominant species of submerged vegetation
3 结果与分析
3.1 沉水植物空间分布特征分析
执行上述分类操作流程,合并太湖7大沉水植物优势种,得到2013年太湖沉水植物优势种空间分布(图5).统计2013年太湖7大沉水植物优势种群落区域面积(图6),结果显示:(1) 马来眼子菜群落是目前太湖分布面积最大、总生物量最高的群落,总分布面积约为111.0km2,主要分布于东太湖、南太湖、胥口湾、东山湾和西山岛附近,在胥口湾构成马来眼子菜+微齿眼子菜+金鱼藻群落,在东山湾以单优势种分布,在南太湖构成马来眼子菜+金鱼藻+苦草群落;(2) 苦草群落的总分布面积约为61.3km2,主要分布于东太湖,呈小片状分布于贡湖湾、南太湖和西山岛附近,伴生种有马来眼子菜、穗花狐尾藻、伊乐藻和菹草等;(3) 菹草群落的总分布面积约为47.4km2,主要呈单优势种分布于梅梁湾,多以伴生种的形式分布于东太湖;(4) 金鱼藻群落的总分布面积约为30.1km2,主要零散地分布于南太湖、东太湖和西山岛附近,伴生种有马来眼子菜、苦草和微齿眼子菜等;(5) 穗花狐尾藻群落的总分布面积约为25.3km2,主要分布于镇湖湾和光福湾,常构成穗花狐尾藻+苦草+马来眼子菜群落和穗花狐尾藻+微齿眼子菜+马来眼子菜群落,多以伴生种分布于东太湖网围养殖区;(6)微齿眼子菜群落的总分布面积约为20.6km2,主要分布于胥口湾和东山湾,在胥口湾构成微齿眼子菜+马来眼子菜+金鱼藻群落,在东太湖的伴生种有马来眼子菜、穗花狐尾藻和苦草等;(7) 伊乐藻群落的总分布面积约为9.4km2,主要分布于东太湖网围养殖区.
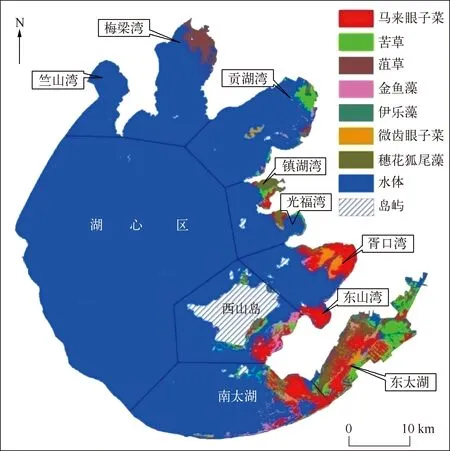
图5 沉水植物优势种的空间分布Fig.5 Spatial distribution of dominant species of submerged vegetation
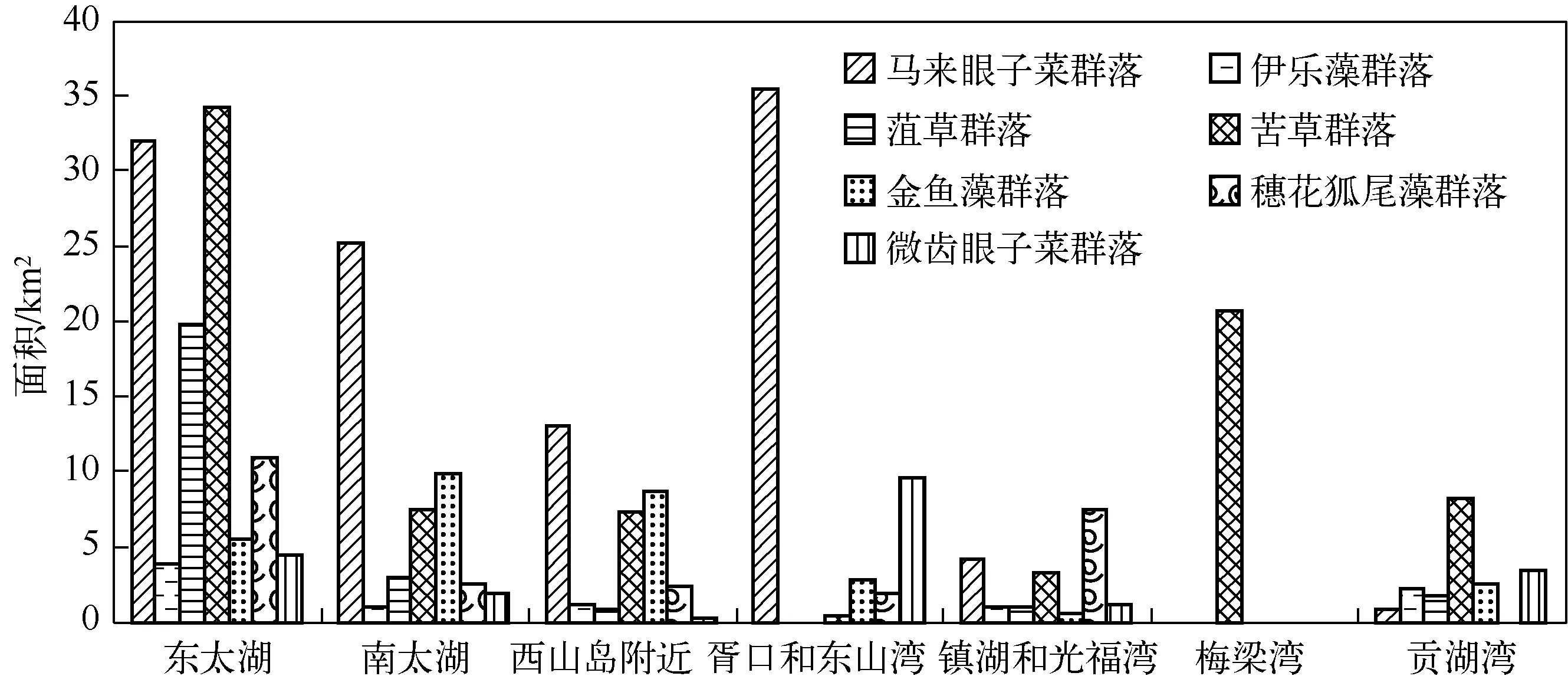
图6 2013年沉水植物优势种群落区域面积统计Fig.6 Regional distribution area of dominant species of submerged vegetation in 2013
3.2 精度验证
为了验证本研究提出的分类特征和决策树对太湖水生植物的分类潜力和普适性,利用水生植物实测数据对2013年7月11日、8月16日和9月26日获取的3期影像分类结果进行验证. 其验证混淆矩阵如表2所示,本研究构建的分类特征和决策树对3期影像分类后总的精度分别为83.04%、81.82%和85.47%.
同时,利用沉水植物优势种的野外调查数据,对本研究得出的2013年太湖沉水植物优势种分布结果进行验证,其验证结果如表3所示,沉水植物优势种总体识别精度为62.20%,Kappa系数为0.53.其中马来眼子菜识别的生产者精度为53.84%,使用者精度为66.67%.19.23%的马来眼子菜被误判成苦草,苦草生产者精度为66.67%,使用者精度为62.50%,20.00%的苦草被误判成马来眼子菜,这说明采用本文提出的生活史判别法,马来眼子菜与苦草仍然具有一定的相关性,主要是因为马来眼子菜和苦草的生活史极其相似,10月下旬马来眼子菜生物量达到最大值,而苦草生物量恰恰也是在10月初达到最大值,本文采用10月28日的影像,日期上已经接近11月,但仍有少量苦草存活下来,这样就影响了判别精度,对于大面积沉水植物遥感识别而言,生活史判别法满足监测的要求.穗花狐尾藻识别的生产者精度为57.89%,使用者精度为78.57%,微齿眼子菜识别的生产者精度为77.78%,使用者精度为63.63%,在7月份穗花狐尾藻和微齿眼子菜同时达到最大生物量,4-5月都为它们的生长旺季,两者之间并没有产生大面积的误判,因为生活史判别法避开了它们暴发的高峰期,根据最小生物量先提取穗花狐尾藻,保证了分类的精度;金鱼藻识别的生产者精度为69.23%,使用者精度为75.00%,独特的生活史特征使其识别精度较高.由于伊乐藻大部分分布在东太湖的网围养殖区,水生植物太过茂密而船只无法通过,采集的样点极少,夏季是菹草的休眠期,也未在同期获得验证样点,所以两者未开展精度验证.
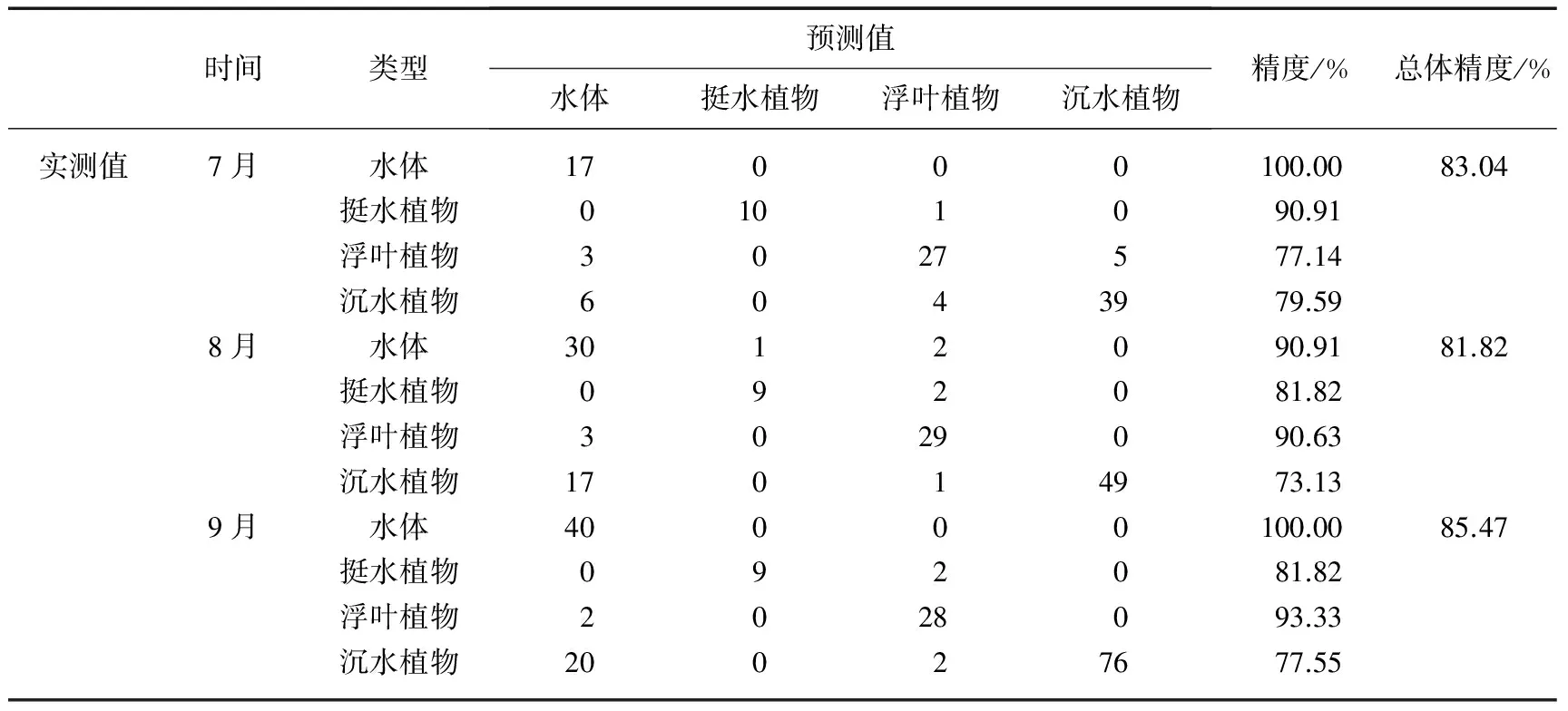
表2 基于决策树的3期影像分类后识别混淆矩阵
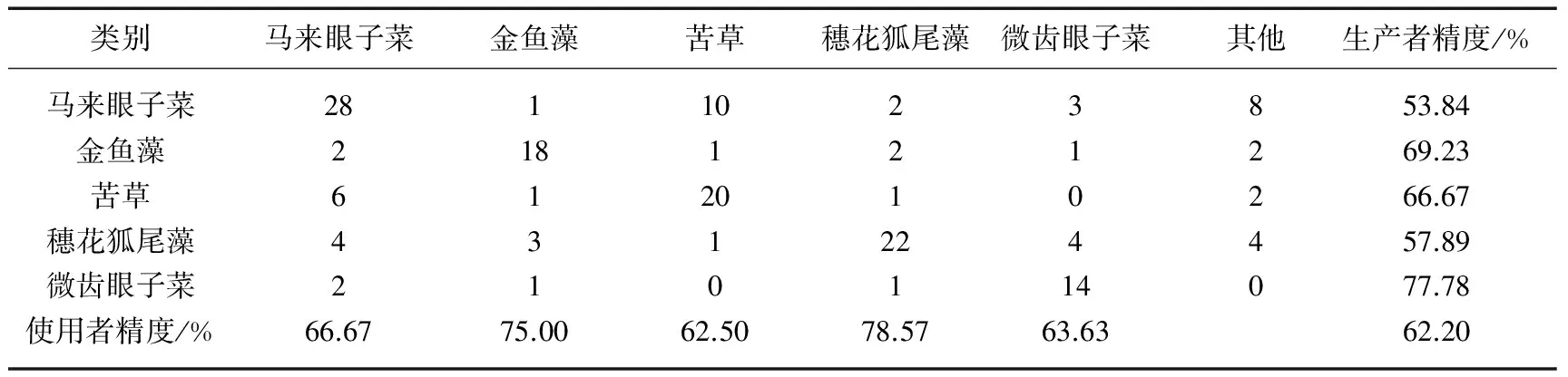
表3 基于生活史的沉水植物优势种分类后识别混淆矩阵
4 结论
以太湖为研究区,基于HJ-CCD影像的波段特征和影像变换技术,提出了太湖挺水、浮叶和沉水植物的影像分类指数,利用CART算法确定分割阈值,分别构建了2013年11个月份的太湖水生植物的决策树分类模型,将太湖水生植物分成挺水、浮叶和沉水植物3种类别.通过实测样点验证,3期影像总体分类精度分别为83.04%、81.82%和85.47%,其中挺水植物分类精度分别为90.91%、81.82%和81.82%;浮叶植物分类精度分别为77.14%、90.63%和93.33%;沉水植物分类精度分别为79.59%、73.13%和77.55%.挺水植物和浮叶植物识别精度较高,沉水植物的识别精度偏低,20.09%的沉水植物被误判成水体,且误判像元覆盖度均低于30%,因此,利用中高分辨率多光谱数据很难识别出覆盖度较低的沉水植物.
根据大量的文献调研和野外实测数据,获取了太湖沉水植物优势种生活史信息,基于多时相HJ-CCD影像的水生植物分类结果,提出了一种考虑生活史信息的太湖沉水植物遥感监测方法,该方法对太湖沉水植物优势种总体识别精度达到62.20%,对于沉水植物中等分辨率遥感影像识别而言实属不易,为太湖沉水植物优势种大面积监测开创了一种新的思路和方法.
[1] 金相灿,刘树坤,章宗涉.中国湖泊环境:第1册.北京:海洋出版社,1995: 234-302.
[2] 王庆海,段留生,李瑞华等.几种水生植物净化能力的比较.华北农学报,2008,23(2): 217-222.
[3] Valta-Hulkkonen K, Pellikka P, Tanskanen Hetal. Digital false colour aerial photographs for discrimination of aquatic macrophyte species.AquaticBotany, 2003, 75(1): 71-88.
[4] Zhu B, Fitzgerald DG, Susan BHetal. Quantification of historical change of two bays of Lake Ontario with three complementary methods.JournalofGreatLakesResearch, 2007, 33: 122-135.
[5] Zhao DH, Jiang H, Yang TW. Remote sensing of aquatic vegetation distribution in Taihu Lake using an improved classification tree with modified thresholds.JournalofEnvironmentalManagement, 2012,95(1): 98-107.
[6] Ma RH, Duan HT, Gu XHetal. Detecting aquatic vegetation changes in Taihu Lake. China using multi-temporal satellite imagery.Sensors, 2008,8(6): 3988-4005.
[7] Yuan L, Zhang L. Identification of the spectral characteristics of submerged plantVallisneriaspiralis.ActaEcologicaSinica, 2006, 26(4): 1005-1011.
[8] Han L, Rundquist DC. The spectral responses ofCeratophyllumdemersumat varying depths in an experimental tank.InternationalJournalofRemoteSensing, 2003, 60: 1463-1471.
[9] 袁 琳.大型沉水植物遥感监测研究[学位论文].上海:华东师范大学,2007.
[10] 李俊生,吴 迪,吴远峰等.基于实测光谱数据的太湖水华和水生高等植物识别.湖泊科学,2009,21(2): 215-222. DOI 10.18307/2009.0209.
[11] 刘伟龙,胡维平,陈永根等.西太湖水生植物时空变化.生态学报,2007,27(1): 1-12.
[12] 蒲云海,李 伟.中国竹叶眼子菜的生态学研究.武汉植物学研究,1999,17(增刊): 65-72.
[13] 刘伟龙,胡维平,谷孝鸿.太湖马来眼子菜生物量变化及影响因素.生态学报,2007,27(8): 3324-3333.
[14] 蒲云海,刘贵华,李 伟.竹叶眼子菜种子的萌发实验研究.武汉植物学研究,2005,23(2): 179-182.
[15] 何 俊,谷孝鸿,刘国锋.东太湖水生植物及其与环境的相互作用.湖泊科学,2008,20(6): 790-795. DOI 10.18307/2008.0618.
[16] 谷孝鸿,陈开宁,胡耀辉等.东太湖伊乐藻的营养繁殖及对渔业污水的净化.上海环境科学,2002,21(1): 43-45.
[17] 连光华,张圣照.伊乐藻等水生高等植物的快速营养繁殖技术和栽培方法.湖泊科学,1996,8(Z1): 11-16. DOI 10.18307/1996.sup02.
[18] 陈开宁,兰策介,史龙新等.苦草繁殖生态学研究.植物生态学报,2006,30(3): 487-495.
[19] 熊秉红,李 伟.我国苦草属(VallisneriaL.)植物的生态学研究.武汉植物学研究,2000,18(6): 500-508.
[20] 靳宝锋,郭友好.微齿眼子菜繁殖生物学特性的初步研究.水生生物学报,2001,25(5): 439-448.
[21] 王 珺,顾宇飞,朱增银等.不同营养状态下金鱼藻的生理响应.应用生态学报,2005,16(2): 337-340.
[22] 栾会妮.若干环境因子对穗状穗花狐尾藻光合作用的影响及鱼草混养研究[学位论文].重庆:西南农业大学,2005.
[23] 程 玉,李 伟.洪湖主要沉水植物群落的定量分析.水生生物学报,1999,23(3): 240-244.
[24] 光玮洁.不同湖泊沉积物营养特征与沉水植物菹草生长的应答关系研究[学位论文].武汉:华中农业大学,2008.
[25] Breiman L, Friedman JH, Olshen RAetal. Classification and regression tree. Monterey: Wadsworth International Group, 1984:1-358.
[26] 陈 云,戴锦芳.基于遥感数据的太湖蓝藻水华信息识别方法.湖泊科学,2008,20(2): 179-183. DOI 10.18307/2008.0207.
[27] 马荣华,孔繁祥,段洪涛等.基于卫星遥感的太湖蓝藻水华时空分布规律认识.湖泊科学,2008,20(6): 687-694. DOI 10.18307/2008.0605.
[28] 陈书琴,李法松,许秋瑾等.菹草在太湖梅梁湾生存对策分析.环境保护科学,2008,34(6): 4-7.
The remote sensing monitoring of dominant species of submerged vegetation of Lake Taihu with the consideration of their living histories
WANG Qi1,ZHOU Xingdong1,LUO Jühua2& CHEN Chong1
(1:DepartmentofUrbanConstructionandEnvironment,JiangsuNormalUniversity,Xuzhou221000,P.R.China)
(2:NanjingInstituteofGeographyandLimnology,ChineseAcademyofSciences,Nanjing210008,P.R.China)
Aquatic vegetation is not only an important group/type of shallow lakes, but also an indicator of lake environment. It plays an important role in ecological restoration and management of shallow lakes. To monitor real-time spatial-temporal distributions of aquatic vegetation in this study, classification tree(CT) model for mapping aquatic vegetation types was developed through HJ-CCD images. Aquatic vegetation of Lake Taihu was classified into emergent, floating-leaved and submerged vegetation by CT model. Living histories of dominant species of submerged vegetation were extracted from literatures and surveys. Combining with living histories and multi-temporal HJ-CCD images, a method was proposed for mapping dominant species of submerged vegetation of Lake Taihu. Based on the method, submerged vegetation in Lake Taihu was subdivided into seven dominant species. By verification of the using field survey points, the overall accuracies of CT model for mapping aquatic vegetation were 83.04%, 81.82% and 85.47%, respectively during three periods of the investigations on July, August and September of 2013. The overall accuracy of dominant species of submerged vegetation was 62.20%. Therefore, CT model and the method for mapping dominant species of submerged vegetation proposed in this study could be helpful for lake management including guiding aquatic vegetation harvesting and ecological restoration. Meanwhile, it lays foundation for further research of historical changes in submerged vegetation of Lake Taihu.
Aquatic vegetation; Lake Taihu; remote sensing; dominant species; living histories; classification tree
*国家自然科学基金重大项目(41230853)、国家自然科学基金项目(41301375)和江苏省普通高校研究生科研创新计划项目(KYLX_1434)联合资助.2014-11-23收稿;2015-03-11收修改稿.王琪(1990~),男,硕士研究生;E-mail: wangqi099x@163.com.
J.LakeSci.(湖泊科学), 2015, 27(5): 953-961
DOI 10.18307/2015.0523
©2015 byJournalofLakeSciences
**通信作者;E-mail:zxdzy111@sina.com.
