接种混合发酵剂生产的低酸度川味香肠加工过程中的蛋白质降解
2015-05-05卿丹丹李跃文
孙 霞,巩 洋,杨 勇,杨 敏,卿丹丹,李跃文,李 静,何 利,李 诚,胡 滨
(四川农业大学食品学院,四川雅安 625014)
接种混合发酵剂生产的低酸度川味香肠加工过程中的蛋白质降解
孙 霞,巩 洋,杨 勇*,杨 敏,卿丹丹,李跃文,李 静,何 利,李 诚,胡 滨
(四川农业大学食品学院,四川雅安 625014)
接种发酵剂并在控温控湿条件下生产低酸度川味香肠,对蛋白质降解情况进行研究,并与自然条件下生产的传统川味香肠(对照组)相比较,揭示低酸度川味香肠加工过程中蛋白质变化规律。结果显示:干燥结束时,低酸度川味香肠的总氮、非蛋白氮和氨基酸态氮分别为4.40%、0.426%和0.366%,均高于对照组;低酸度川味香肠的游离氨基酸总量为605.06mg/100g,高于对照组的497.47mg/100g(p<0.01);肌浆蛋白和肌原纤维蛋白在成熟过程中逐渐降解,大分子条带在干燥过程中发生了明显降解(p<0.05),低酸度川味香肠的蛋白质降解更为显著(p<0.05)。
低酸度川味香肠,蛋白质降解,游离氨基酸,肌浆蛋白,肌原纤维蛋白
川味香肠是我国特色的传统腌腊肉制品,因其香气浓郁、风味独特、麻辣爽口而深受消费者的喜爱。低酸度川味香肠一般通过接种发酵剂,利用现代控温控湿工艺使香肠pH达到5.50以上[1]。在香肠的发酵、成熟和干燥过程中,肌肉蛋白降解是发酵香肠加工过程中重要的生化反应,蛋白质降解后形成的多种低分子量的化合物如氨基酸、肽类、醛类和有机酸等是重要的风味物质,对发酵香肠的风味有重要影响[2]。
目前,国内外学者已对发酵香肠加工过程中蛋白质降解及其降解产物进行了系统的研究。Frederic等[3]发现发酵香肠中蛋白质的降解产物肽,在氨肽酶的作用下降解为游离氨基酸。Casaburi等[4]发现肌浆蛋白和肌原纤维蛋白的降解成分为肽、游离氨基酸这些成分是发酵香肠风味物质的重要来源,其中寡肽、游离氨基酸仍会进一步的酶解和化学反应。Careri等[5]研究发现蛋白质降解虽然有利于发酵香肠风味的形成,但是若过度降解(高于30%),则产生不愉快的口味如苦味和金属味。张雪梅[6]对四川传统发酵香肠在加工过程中蛋白质的降解规律进行了研究,发现未检测到天冬氨酸、丝氨酸和胱氨酸,其余的各种氨基酸含量相对于原料肉都有不同程度的增加,其中丙氨酸、脯氨酸、谷氨酸含量最高。帅瑾[7]通过接种植物乳杆菌和葡萄球菌生产川式萨拉米香肠,发现必需氨基酸总量比自然发酵组提高38.45%,游离氨基酸总量比自然发酵组提高21.32%,但是产品pH为4.63,口感较酸,与传统四川香肠的口感差异很大。目前关于低酸度川味香肠在加工过程中蛋白质降解情况的研究鲜有报道。
本研究通过以植物乳杆菌、戊糖片球菌、葡萄球菌作为发酵剂,采用控温控湿现代工艺对低酸度川味香肠在加工过程中总氮、非蛋白氮、氨基酸态氮、游离氨基酸、肌浆蛋白和肌原纤维蛋白的变化情况进行研究,以期为提高低酸度川味香肠的品质提供初步的理论依据。
1 材料与方法
1.1 材料与仪器
葡萄球菌(Staphylococcus)、植物乳杆菌(Lactobacillusplantarum)和戊糖片球菌(Pediocossuspentosaceus) 均由四川农业大学肉品研究室从传统自然发酵四川香肠中分离得到[7-8];新鲜猪后腿肉、肠衣、辣椒、花椒和食盐等调味料 均购于四川雅安农贸市场;考马斯亮蓝G-250、牛血清白蛋白、过硫酸铵、丙烯酰胺、甲叉丙烯酰胺、四甲基乙二胺 均为Sigma公司产品;溴酚蓝、三氯乙酸、甘油、碘化钾、硫酸铜、硫酸钾、浓硫酸、氢氧化钠 以上试剂均为分析纯。
LHS-250SC恒温恒湿培养箱 上海荣丰科学仪器有限公司;ST16R高速冷冻离心机 北京博仪恒业科技发展有限公司;TS-2000A型摇床 海门市麒麟医用仪器厂;HWS24电热恒温水浴锅 上海一恒公司;721分光光度计 上海第三分析仪器厂;日立L-8900氨基酸自动分析仪 日本;DYY-6D电泳仪 DYC2-24DN电泳槽 北京六一仪器厂;Gel-Doc-XR+凝胶成像仪 BIO-RAD。
1.2 低酸度川味香肠的制作
参照张雪梅[6]的配方:猪肉100kg(皮下脂肪20kg,瘦肉80kg),食盐2.5kg,辣椒粉1.0kg,花椒粉0.4kg,白砂糖1.0kg,味精0.15kg,白酒1.0kg,十三香0.05kg,NaNO30.025kg,NaNO20.0075kg。
采用现代控温控湿工艺制作。工艺流程:原料肉→预处理→绞碎→添加配料→接种→混合→灌肠→发酵→成熟→干燥→成品
发酵剂的制作:30℃条件下液体培养基培养筛选菌株14h后,置于4℃条件下4000r/min冷冻离心10min,用无菌生理盐水洗涤一次再离心,收集菌体并适量稀释,运用血球计数法计数,接种量为106cfu/g,植物乳杆菌、戊糖片球菌和葡萄球菌按1∶1∶1的比例混合加入肉料中。
本研究在预实验、单因素实验、正交实验的基础上,确定低酸度川味香肠(pH为5.50)的工艺参数:发酵温度20℃,发酵相对湿度75%,发酵时间12h,葡萄糖添加量0.10%;成熟温度13℃,成熟相对湿度60%,成熟时间4d;干燥温度55℃,干燥时间24h。实验组是接种混合发酵剂并在控温控湿条件下生产的低酸度川味香肠,对照组是不接种发酵剂在自然条件下生产的传统川味发酵香肠。
1.3 样品采集
取川味香肠在加工过程中的7个工艺点,0h(原料肉)、12h(发酵结束)、36h(成熟1d)、60h(成熟2d)、84h(成熟3d)、108h(成熟4d)、132h(干燥结束)的香肠作为指标测定样品。
1.4 实验方法
1.4.1 总氮测定 称取绞碎的香肠样品1.5g于消化管中,参照GB/T5009.5-2010的半微量凯氏定氮法测定总氮含量[6]。
1.4.2 非蛋白氮测定 称取绞碎香肠样品10g于三角瓶中,加入90mL蒸馏水,振摇40min,取25mL滤液,并加入25mL 10%三氯乙酸溶液,过滤后取10mL滤液消化,参照GB/T5009.5-2010的半微量凯氏定氮法测定非蛋白氮含量。
1.4.3 氨基酸态氮测定 采用甲醛滴定法进行测定[6]。
1.4.4 游离氨基酸测定 参照程燕等[9]的方法:精确称取香肠样品5.000g,加入25mL 10%的磺基水杨酸,置于4℃冰箱中沉淀2h,吸取上清液冷冻离心(10000r/min,4℃,15min),经纱布过滤后用旋转蒸发仪浓缩干燥,加入去离子水再浓缩干燥(重复操作一次),然后用0.02mol/L盐酸定容至10mL,用超滤膜过滤后取20μL滤液用氨基酸分析仪测定。
1.4.5 肌浆蛋白与肌原纤维蛋白的SDS-PAGE凝胶电泳 肌浆蛋白和肌原纤维蛋白的提取参照Doerscher等[10]和Molina等[11]的方法。样品处理方法和电泳参数参照汪家政等[12]和郭尧君[13]的方法,选分离胶浓度为12%,浓缩胶浓度为4%进行电泳。
1.5 数据处理与统计分析
实验数据为三次统计的平均值,使用SPSS17.0统计软件描述性分析和方差分析程序进行处理,并进行多重比较(p<0.05)。
2 结果与讨论
2.1 低酸度川味香肠加工过程中总氮的变化
由图1可知,总氮初始含量为3.6%,在发酵结束,实验组和对照组的总氮含量均呈下降趋势(p<0.05),实验组总氮含量下降速度大于对照组,这可能是由于实验组发酵温度高,水分散失程度大,导致水溶性蛋白质随着盐水的渗出而流失。在成熟期,实验组和对照组的总氮含量呈明显的上升趋势(p<0.01),且实验组总氮含量高于对照组,一方面可能是由于实验组接种了乳酸菌和葡萄球菌,促进蛋白质降解为游离氨基酸、小分子肽以及部分风味成分[14];另一方面可能是由于此阶段实验组的pH逐渐下降并维持在5.50~5.70,此条件下组织蛋白酶活性增强。在干燥期,实验组和对照组的总氮含量均呈缓慢的下降趋势(p<0.05),最终含量分别达到4.40%和4.09%,这可能是由于高温促使水分大量散失,食盐浓度增大,组织蛋白酶活性受到抑制,从而使蛋白质降解变慢,这与吕舒[15]等对传统发酵四川香肠的研究结果是一致的。

图1 低酸度川味香肠加工过程中总氮的变化Fig.1 Changes of total nitrogen in low acidity Sichuan-style sausage during processing
2.2 低酸度川味香肠加工过程中非蛋白氮的变化
非蛋白氮含量是指除了蛋白质以外的所有游离氨基酸、多肽和短肽的总含量。非蛋白氮是表征蛋白质降解的一个重要指标,是川味香肠产生风味的重要前提物质。由图2可知,川味香肠在整个加工过程中,实验组和对照组的非蛋白氮含量呈逐渐上升趋势(p<0.05),这是因为组织蛋白酶等对蛋白质的降解作用贯穿于川味香肠加工的整个过程,其降解产生的游离氨基酸和肽类物质不断积累,促使非蛋白氮含量持续上升,这与Soriano[16]等的研究结相似。在发酵期,实验组非蛋白氮含量上升速度大于对照组(p<0.01),这可能是由于实验组接种发酵剂且发酵温度较高,发酵剂在适宜条件下产生降解蛋白的酶,高温促使组织蛋白酶和寡肽酶的活性增强,使蛋白质水解产生游离氨基酸和肽类物质,这与朝鲜娜[17]等研究发酵剂对熏马肠成熟过程中蛋白质降解的影响结果是相似的。在成熟期,实验组和对照组的非蛋白氮含量呈缓慢上升的趋势,一方面可能是由于成熟温度的降低抑制了组织蛋白酶活性和微生物活性;另一方面也可能是由于水分不断减少,高浓度食盐抑制了组织蛋白酶活性,从而使蛋白质降解变慢。在干燥期,实验组和对照组的非蛋白氮含量呈缓慢上升的趋势,最终含量分别为0.426%和0.215%,这可能是由于较高的干燥温度使食盐含量降低,微生物活动增强,使非蛋白氮含量升高。
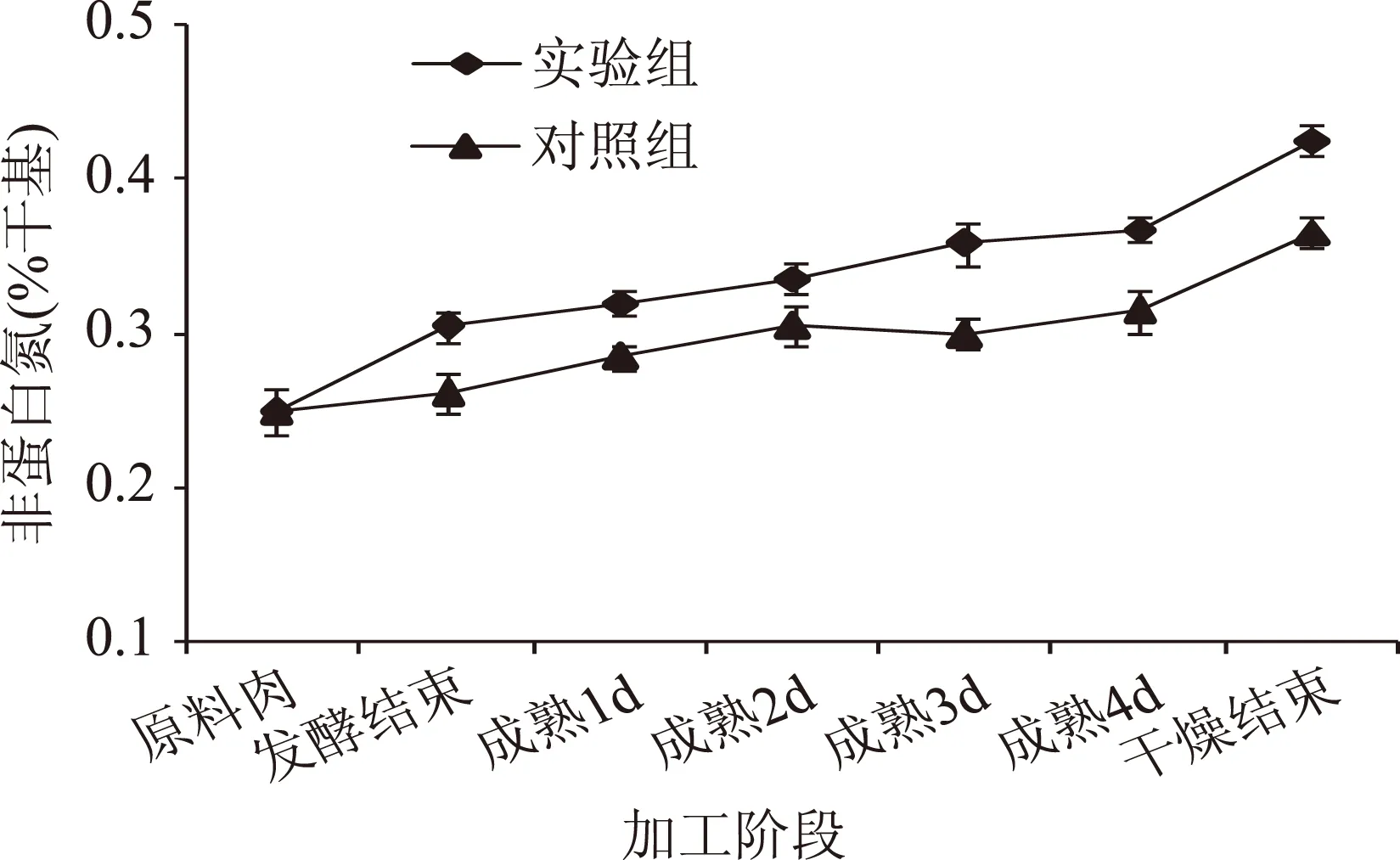
图2 低酸度川味香肠加工过程中非蛋白氮的变化Fig.2 Changes of non-protein nitrogen in low acidity Sichuan-style sausage during processing
2.3 低酸度川味香肠加工过程中氨基酸态氮的变化
由图3可知,在川味香肠整个加工过程中,氨基酸态氮呈上升趋势(p<0.05)。在发酵结束,实验组氨基酸态氮含量明显高于对照组(p<0.01),这可能是由于实验组接种了发酵剂,微生物在较高的发酵温度下产生的蛋白酶分解肌原纤维蛋白从而促进蛋白质的降解,使香肠中氨基酸态氮含量提高。在成熟期,实验组和对照组的氨基酸态氮变化不显著(p>0.05),这可能是由于成熟温度较低,抑制了微生物活动和组织蛋白酶活性使蛋白质降解作用减少,此外部分的氨基酸态氮参与了川味香肠风味的形成,自身被消耗。在干燥期,实验组和对照的氨基酸态氮含量迅速上升(p<0.01),这可能是因为干燥期水分含量降低,使氨基酸态氮相对浓度增大,也可能是因为水分散失导致食盐含量增加,蛋白质结构遭到破坏,使蛋白酶更易作用于蛋白质[18]。最终实验组和对照组氨基酸态氮分别为0.366%和0.185%,实验组高于对照组,可能是因为实验组的pH在5.50左右,此时的酸性环境使蛋白质变性,实验组水分散失量和食盐含量均大于对照组,实验组中的蛋白酶更易作用于蛋白质,从而使氨基酸态氮含量高于对照组。

图3 低酸度川味香肠加工过程中氨基酸态氮的变化Fig.3 Changes of amino acidic nitrogen in low acidity Sichuan-style sausage during processing
2.4 低酸度川味香肠加工过程中游离氨基酸的变化
由表1可知,除Cys外,低酸度川味香肠加工结束时的其余各种游离氨基酸含量与原料肉相比有明显的增加(p<0.05),这与国内外有关发酵香肠的研究报道相似[19-21]。经过发酵,除Ser、Gly、Ala、Cys和Leu含量下降外,其余各种氨基酸含量呈整体上升趋势,实验组发酵结束后游离氨基酸的总量比原料肉中游离氨基酸总量提高81.0%,对照组发酵结束后游离氨基酸的总量比原料肉中游离氨基酸总量提高66.2%,且实验组比对照组高8.4%,说明接种的发酵剂促进了原料肉中蛋白质降解,产生游离氨基酸,这与Candogan等[22]的研究结果一致。低温成熟阶段,Asp、Ser、Glu、Gly、Ala、Val、Met、Ile、Leu、Tyr、Phe、Arg和Pro都有所增加,成熟结束后实验组的游离氨基酸总量比对照组提高13%,这可能是由于实验组pH在5.50~5.70之间,有利于蛋白酶作用于蛋白质,使蛋白质降解产生游离氨基酸。干燥结束后,实验组的游离氨基酸总量为605.06mg/100g,明显高于对照组的497.47mg/100g(p<0.01),这说明接种的发酵剂促进了蛋白质的降解;实验组的游离氨基酸总量是成熟结束后的1.95倍,对照组的游离氨基酸总量是成熟结束后的1.81倍,这说明较高的干燥温度促进了蛋白质降解,产生更多的游离氨基酸。

表1 低酸度川味香肠加工过程中游离氨基酸的变化(干基mg/100g)Table 1 Changes of free amino acids in low acidity Sichuan-style sausage during processing(mg/100g)
注:表中数据为均值±标准差;同行均值有共同上标字母者表示差异不显著(p>0.05)。
最终实验组游离氨基酸总量的比对照组提高21.6%,实验组的必需氨基酸(Lys、Phe、Met、Ile、Thr、Leu、Val)总量比对照组提高31.5%。在整个加工过程中,含量较高的游离氨基酸有苏氨酸、谷氨酸、丙氨酸等,其余各种游离氨基酸的含量都超过其成为阈值,对发酵香肠滋味的形成十分重要[23]。胱氨酸含量变化不显著(p>0.05),这是因为胱氨酸是半胱氨酸的氧化形式,而胱氨酸具有活泼的硫氢基,易于参加氧化反应过程失去氨基酸特性,这与赵改名[24]的研究结果相似。
2.5 低酸度川味香肠加工过程中肌肉蛋白质的降解变化
2.5.1 肌浆蛋白的降解变化 由图4可知,在低酸度川味香肠整个加工过程中,实验组的肌浆蛋白条带发生了明显变化,分子量为157ku(f1)、97.2ku(f2)、44.3ku(f4)、29.0ku(f5)、23ku(h1)、14.3ku(h2)的蛋白条带从发酵期到干燥期就逐渐变弱甚至消失,说明在加工阶段发生了降解;对照组各阶段变化不明显,只有分子量在97.2ku(f2)、44.3~66.4ku附近的蛋白条带干燥期变弱甚至消失。
在发酵期,实验组肌浆蛋白的分子量在97.2ku(f2)的蛋白条带颜色略微变弱,而对照组的蛋白条带基本无变化,这可能是由于实验组接种了发酵剂并且发酵温度较高,微生物产生的蛋白酶作用,这与Mauriello等[25]的研究结果相似。在成熟期,实验组肌浆蛋白的分子量在157ku(f1)、97.2ku(f2)、44.3ku(f4)、29.0ku(f5)、23ku(h1)、14.3ku(h2)的蛋白条带逐渐变弱,在成熟到第4d部分蛋白发生了明显的降解,生成了一些分子量较小的片段,这些片段通过其它的生化反应,生成了低酸度香肠的风味和滋味物质[26],这可能是由于实验组接种了发酵剂并在成熟后期香肠的pH在5.50~5.70之间,此时组织蛋白酶活性较高,也可能是由于在低酸性环境下蛋白质变性,蛋白酶更易作用于蛋白质,从而使大分子的蛋白条带发生了部分降解,这与Casaburi等[27]的研究结果相似;而对照组肌浆蛋白的条带变化不明显,降解程度非常小,这可能是由于自然环境下较低的温度抑制了蛋白酶的活性。
在干燥期,实验组肌浆蛋白的分子量在157ku(f1)、97ku(f2)的蛋白条带消失,发生了明显的降解,而对照组肌浆蛋白的条带是由粗变细,这可能是由于实验组香肠的pH在5.50~5.70,使蛋白质变性,再加上干燥期较高的干燥温度使水分大量散失,导致食盐含量增加,蛋白质结构遭到破坏,蛋白酶更易作用于蛋白质,从而使大分子量的蛋白发生降解;实验组和对照组肌浆蛋白的分子量在66.4ku(f3)、29.0~44.3ku之间的蛋白条带变粗,这可能是由于较高的干燥温度使小分子量的蛋白发生了聚合[28],从而使此分子量的蛋白条带变粗。

图4 低酸度川味香肠加工过程中肌浆蛋白SDS-PAGE电泳图Fig.4 SDS-PAGE of sarcoplasmic proteins in low acidity Sichuan-style sausage during processing注:M-标准蛋白;1-原料肉;2-发酵结束;3-成熟1d;4-成熟2d;5-成熟3d;6-成熟4d;7-干燥结束,图5同。

图5 低酸度川味香肠加工过程中肌原纤维蛋白SDS-PAGE电泳图Fig.5 SDS-PAGE of myofibrillar proteins in low acidity Sichuan-style sausage during processing
2.5.2 肌原纤维蛋白的降解变化 由图5可知,在低酸度川味香肠整个加工过程中,对照组肌原纤维蛋白的条带变化不明显,实验组与对照组相比,肌原纤维蛋白发生了一定的降解,随着加工的进行,实验组低分子量的蛋白条带越来越少,尤其是14.3ku(h2)附近的条带明显变弱;分子量在44.3ku的蛋白条带在整个加工过程中几乎没发生降解,这与王振宇等[28]的研究结果相似。
在发酵期,实验组肌原纤维蛋白的分子量在200ku(f1)的蛋白条带明显变细,对照组蛋白条带变化较小,这可能是由于实验组接种发酵剂,微生物产生了蛋白酶并在较高的发酵温度下使大分子量的蛋白降解。在成熟期,实验组肌原纤维蛋白的分子量在52ku(f3)、24ku(f5)、18ku(h1)、14.3ku(h2)的蛋白条带逐渐变浅且比对照组蛋白条带变化明显,此阶段肌原纤维蛋白片段缓慢降解,生成了一些新的条带,但降解作用并不剧烈,这可能是由于成熟温度较低,抑制了组织蛋白酶活性,蛋白降解缓慢,但在成熟的整个过程来看,蛋白质的降解作用仍在继续进行。
在干燥期,实验组和对照组肌原纤维蛋白的分子量在200ku(f1)、97.2ku(f2)、24ku(f5)、18ku(h1)、14.3ku(h2)的蛋白条带变浅变细,蛋白质发生了明显降解,并且实验组比对照组降解的更加剧烈,这可能是由于实验组香肠的酸性环境使蛋白质变性,蛋白酶更易作用于蛋白质;而分子量在29.0~44.3ku之间的部分条带和分子量为52ku(f3)的蛋白条带变粗,这可能是由于较高的干燥温度使小分子量的蛋白发生聚合。
3 结论
采用控温控湿工艺及接种发酵剂生产的低酸度川味香肠(实验组)和自然条件下生产的传统川味香肠(对照组)在加工过程中非蛋白氮、氨基酸态含量差异显著(p<0.05)。低酸度川味香肠在整个加工过程中,非蛋白氮和氨基酸态氮显著上升(p<0.01),实验组分别为0.426%和0.366%,均高于对照组的0.215%和0.185%,说明实验组接种的发酵剂和较高的发酵温度共同促进了蛋白质的降解。总氮在加工过程中呈先降低后升高再下降的趋势,但整体变化都比较平缓,最终实验组和对照组的含量分别达到4.40%和4.09%。随着加工工艺的进行,蛋白质降解加剧,最终实验组和对照组的游离氨基酸总量分别达到605.06mg/100g和497.47mg/100g。从SDS-PAGE电泳图谱可以看出,肌浆蛋白和肌原纤维蛋白在成熟过程中逐渐降解,大分子条带在干燥过程中发生明显降解,且实验组的蛋白质降解比对照组更明显。
[1]凌静. 发酵肉制品的现状和发展趋势[J].肉类研究,2007,13(10):5-7.
[2]Toldra F. Proteolysis and lipolysis in flavour development of dry-cured meat products[J]. Meat Science,1998,49(1):101-110.
[3]Frederic L,Jurgen V,Luc D V. Functional meat starter cultures for improved sausage fermentation[J]. International Journal of Food Microbiology,2006,106(2):270-285.
[4]Casaburi A,Monaco R D,Cavella S,et al. Proteolytic and lipolytic starter cultures and their effect on traditional fermented sausages ripening and sensory traits[J]. Food Microbiology,2008,25(2):335-347.
[5]Careri M,Mnagia A,Babrieril G,et al. Sensory Property relationships to chemical data of ltalina-type dry-cured ham[J]. Journal of Food Science,1993,58(5):968-972.
[6]张雪梅. 四川香肠生产过程中理化特性、微生物特性及产香葡萄球菌的筛选与应用[D]. 雅安:四川农业大学,2011.
[7]帅瑾.传统自然发酵四川香肠中乳酸菌的分离、鉴定及其应用[D]. 雅安:四川农业大学,2013.
[8]杨勇,张雪梅,程艳,等. 四川香肠中产香葡萄球菌的分离与鉴定[J]. 食品与发酵工业,2011,37(6):29-34.
[9]程燕,杨勇,帅谨,等. 腐生葡萄球菌S25在四川香肠加工过程中对脂肪和蛋白质的影响[J]. 食品科学,2012,33(23):223-227.
[10]Doerscher D R,Briggs J L,Lonergan S M. Effects of pork collagen on thermal and viscoelastic properties of purified porcine myofibrillar protein gels[J]. Meat Science,2004,66(1):181-188.
[11]Molina I,Toldra F. Detection of Proteolytic activity in microorganisms isolated from dry-cured ham[J]. Journal of Food Science,2006,57(6):1308-1310.
[12]汪家政,范明.蛋白质技术手册[M].北京:科学出版社,2001:77-110.
[13]郭饶君. 蛋白质电泳实验技术[M].北京:科学出版社,2001:86.
[14]张波,李开雄,卢士玲,等.发酵剂对风干羊肉理化特性变化和蛋白质降解的影响[J].食品工业科技,2013,34(14):194-199.
[15]吕舒.四川香肠加工贮藏过程中表层、中层、内层的理化特性及微生物的变化[D].雅安:四川农业大学,2013.
[16]Soriano A,Garca A. Lipolysis,proteolysis,physic-chemical and sensory characteristics of different types of Spanish ostrich salchichon[J]. Meat Science,2007,75(4):661-668.
[17]朝鲜娜,卢士玲,李开雄.发酵剂对熏马肠成熟过程中蛋白质降解的影响[J].食品与发酵工业,2013,37(1):215-219.
[18]Zhao G.M,Zhou G.H,Wang Y L,et al. Time-related changes in cathepsin B and L activities during processing of Jinhua ham as a function of pH,salt and temperature[J]. Meat Science,2005,70(2):381-388.
[19]Hughes M C,Arendt E K,Kerry JP,et al. Characterization of proteolysis during ripening of semi-dry fermented sausages[J]. Meat Science,2002,62(3):205-216.
[20]Rabie M A,Peres C,Malcata F X. Evolution of amino acids and biogenic amines throughout storage in sausages made of horse,beef and turkey meats[J]. Meat Science,2014,96(1):82-87.
[21]赵俊仁,孔保华. 复合发酵剂生产风干肠对产品质构及游离氨基酸的影响[J].食品工业科技,2007,28(10):132-135.
[22]Candogan K.,Wardlaw F B,Actonb J C. Effect of starter culture on proteolytic changes during processing of fermented beef sausages[J]. Food Chemistry,2009,116(3):731-737.
[23]Herranz B,Hoz L,Hierro E,et al. Improvement of the sensory properties of dry-fermented sausages by the addition of free amino acids[J]. Food Chemistry,2005,91(4):673-682.
[24]赵改名,柳艳霞,高晓平,等.现代工艺金华火腿中游离氨基酸含量的研究[J].食品科学,2009,30(21):152-154.
[25]Mauriello G,Casaburi A,Villani F. Proteolytic activity of Staphylococcus xylosus strains on pork myofibrillar and sarcoplasmatic proteins and use of selected strains in the production of “Naples type” salami[J]. Journal of Applied Microbiology,2002,92(3):482-490.
[26]Spaziani M,Torre M D,Stecchini M L. Changes of physicochemical,microbiological,and textural properties during ripening of Italian low-acid sausages. Proteolysis,sensory and volatile profiles[J]. Meat Science,2009,81(1):77-85.
[27]Casaburi A,Monaco R D,Cavella S,et al. Proteolytic and lipolytic starter cultures and their effect on traditional fermented sausages ripening and sensory traits[J]. Food Microbiology,2008,25(2):335-347.
[28]王振宇,刘欢,马长伟. 热处理下的猪肉蛋白质的特性[J]. 食品科学,2008,29(5):73-77.
Protein degradation of low acidity Sichuan-stylesausage inoculated mixed strains during processing
SUN Xia,GONG Yang,YANG Yong*,YANG Min,QING Dan-dan,LI Yue-wen,LI Jing,HE Li,LI Cheng,HU Bin
(College of Food Science,Sichuan Agricultural University,Ya’an 625014,China)
The low acidity Sichuan-style sausage was processed with inoculated starter under controlling temperature and humidity,compared with traditional Sichuan-style natural fermentation sausage(control group)to verify the protein change during processing. The results showed that total nitrogen,non-protein nitrogen and amino acidic nitrogen were 4.40%,0.426% and 0.366%,respectively,which were higher than control group at the end of drying. The content of all free amino acid of low acidity Sichuan-style sausage was 605.06mg/100g(p<0.01),which was higher than control group(497.47mg/100g)at the end of drying. The sarcoplasmic protein and myofibrillar protein were degraded gradually during ripening,and the protein of molecular weight greater were degraded obviously(p<0.05).The protein of low acidity Sichuan-style sausage was degraded significantly during processing compared with control group(p<0.05).
low acidity Sichuan-style sausage;protein degradation;free amino acid;sarcoplasmic protein;myofibrillar protein
2014-11-05
孙霞(1989-),女,硕士研究生,研究方向:肉品科学与技术。
*通讯作者:杨勇(1969-),男,博士,教授,研究方向:肉品科学与技术。
四川省科技厅成果转化项目(2013NC0052)。
TS251.65
A
1002-0306(2015)13-0066-06
10.13386/j.issn1002-0306.2015.13.005
