太湖湖滨带浮游植物群落特征及指示种筛选
2015-04-26徐东炯
陈 桥,陆 茸,徐东炯,张 咏,汤 云
1.常州市环境监测中心,江苏省环境保护水环境生物监测重点实验室,江苏 常州 213001
2.江苏省环境监测中心,江苏 南京 210036
湖滨带是湖泊流域水生态系统与陆地生态系统间一种非常重要的生态过渡带,是湖泊的一道保护屏障[1]。天然的湖滨带由陆向辐射带、水位变幅带、水向辐射带组成[2]。然而,由于水利防洪的需要,全长405 km[3]的太湖湖滨岸线有73%[4]修建了防洪大堤,原有水陆交错的物质、能量流动被阻隔,生态系统结构的连续性与完整性被破坏,导致了湖滨带生态系统功能的退化。浮游植物是湖泊生态系统的初级生产者,是水环境的重要指示生物[5]。由于浮游植物大量生长引发的藻类水华问题已经成为太湖水生态系统最突出的矛盾。目前,关于太湖湖体浮游植物的研究较多[6-12],主要涉及群落结构的演替以及与氮磷等营养盐关系研究方面,针对湖滨带的相关研究工作较少。鉴于此,以太湖湖滨带为对象,开展浮游植物群落结构特征及其与环境因子的关系研究,探寻能够指示关键变量的指示物种,为生态修复、环境监测及管理提供支撑。
1 实验部分
1.1 点位布设
根据李春华等[1]的研究,太湖湖滨带的范围为大堤以内(水向)50~100 m的环形区域,并可以分为贡湖湾、梅梁湾、竺山湖、西部沿岸、南部沿岸、东太湖、东部沿岸7个区域。竺山湖、梅梁湾、西部沿岸湖滨带以人工直立驳岸为主,水生植物破坏严重,仅在极少部分岸带有芦苇分布,总体水质感官较差,夏季蓝藻水华频发,其中竺山湖水质最差,西部沿岸蓝藻水华堆积最为严重。东太湖是典型的养殖基地,水体及沉积物有机质相对较高。东部沿岸以自然山体驳岸为主,水生植被丰富,水质较好。贡湖湾是重要的引调水湖区。南部沿岸生境条件介于西部沿岸及东部沿岸之间。以均匀布点的方式在湖滨带范围内布设49个监测点位,具体见图1。
1.2 样品采集与处理
按照水期,分别于2009年12月、2010年4、8月监测太湖湖滨带浮游植物群落结构及水质状况。浮游植物样品的采集、预处理及镜检方法参照《水和废水监测分析方法》(第四版)。根据检索资料[13-14],将标本鉴定至尽量低的分类单元。使用YSI6600现场测定水温(T)、溶解氧(DO)、pH,使用塞氏盘测定透明度(SD),同时采集水样[15]分析总磷(TP)、总氮(TN)、氨氮(NH3-N)、硝酸盐氮(NO3-N)、高锰酸盐指数(CODMn)、叶绿素a(chl-a)、悬浮物(SS),计算综合营养状态指数(TSI)[16]。
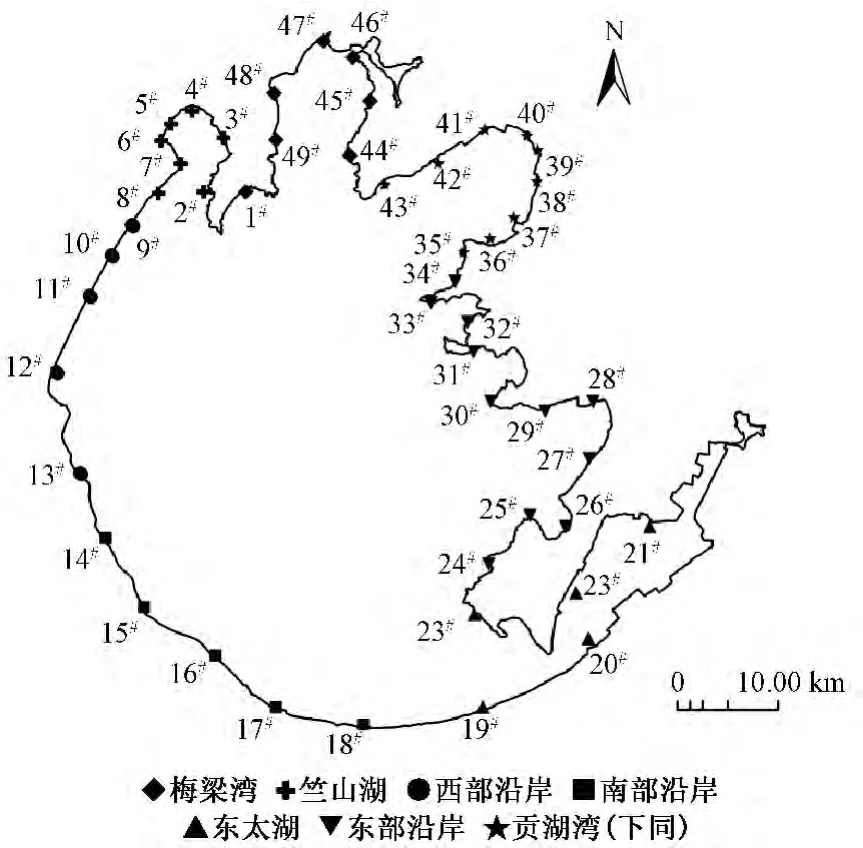
图1 太湖湖滨带形状及监测点位
1.3 数据分析
1.3.1 优势种及优势度
由式(1)来确定:

式中:y为优势度,fi为i种的空间出现频率,pi为i种数量占总个体数的比例,y>0.02时,i种定为优势种[17]。同时统计i种成为优势种的空间频率,即优势频率(φ)。
1.3.2 多样性指数
采用 Shannon-Wiener多样性指数(H)及Pielou均匀度指数(J)来描述物种多样性,见式(2)。
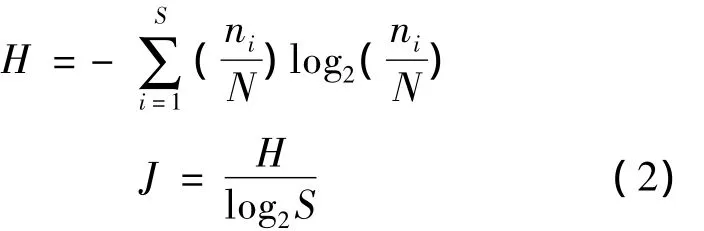
式中:S为总分类单元数,N为总个体数,ni为i种个体数。
1.3.3 排序分析及指示物种筛选
分析物种与环境变量的关系时,暂不考虑浮游植物群落与水质的时间演替,将3次结果算术平均后进行排序分析。为了降低稀有物种对分析结果的干扰,剔除出现频率低于5%的物种,并对剩余物种数据进行对数转换[lg(x+1)]。转换后的物种数据进行除趋势对应分析(DCA),确定湖滨带浮游植物群落的分布特征。将经过以上步骤预处理的浮游植物数据与水质进行冗余分析(RDA),利用向前引入法逐步筛选,通过蒙特卡罗置换检验(Monte-Carlo permutation test,P<0.05)评估每个备选环境变量对于解释物种变异的贡献。最后,剔除环境变量中方差膨胀因子大于5的变量。将贡献最大的环境变量作为关键变量,与物种作排序图,显示适合度范围为50% ~100%的物种,筛选指示关键变量的浮游植物指示种。
基础数据统计采用Excel2007,相关性分析在SPSS15.0中进行,排序分析在 CANOCO4.5中进行,空间分布图在Mapinfo10.0中绘制,其他图形在Origin8.0中绘制。
2 结果与分析
2.1 湖滨带水质及综合营养状况
太湖湖滨带各湖区水质及综合营养状况结果见表1。
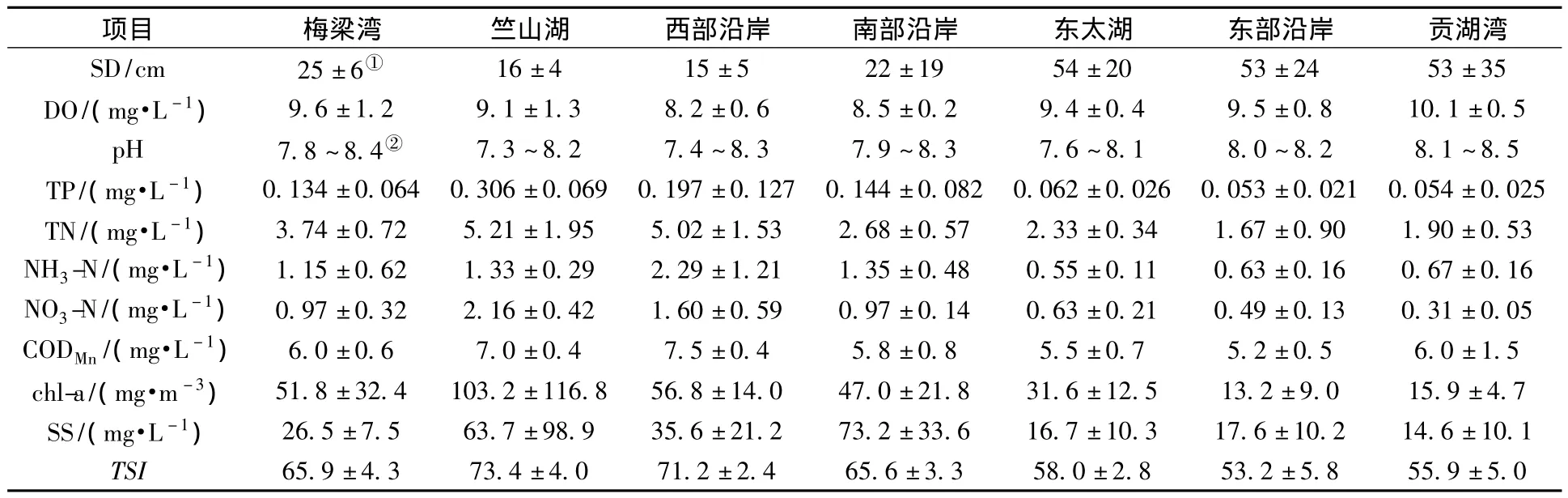
表1 太湖湖滨带各湖区水质及综合营养状况
由表1可以看出,各指标之间存在显著的空间差异(t=2.57~2.94 ,P<0.05)。东部湖区(东太湖、东部沿岸、贡湖湾)SD相对较高,营养盐、CODMn及chl-a浓度相对较低,总体处于轻度富营养化;西部湖区(竺山湖、西部沿岸)SD相对较低,营养盐、CODMn及chl-a浓度相对较高,总体处于重度富营养化;梅梁湾及南部沿岸介于东部湖区及西部湖区之间。从水质类别来看[18],东部沿岸介于Ⅴ类~劣Ⅴ类,其余湖区则均为劣Ⅴ类,主要超标污染物为TN、TP。各湖区TN、TP从大到小依次为竺山湖>西部沿岸>梅梁湾>南部沿岸>东太湖>贡湖湾>东部沿岸。
2.2 浮游植物物种组成及优势类群
3次共采集到浮游植物128个分类单元,其中蓝藻、绿藻、硅藻、裸藻、甲藻、隐藻、金藻的分类单元数分别为 12、53、36、17、5、3、2。从个体密度来看,太湖湖滨带浮游植物密度总体较高,平均达5.97×107cells/L,多样性及均匀性水平较低,分别仅为 1.15、0.43。空间上,竺山湖、梅梁湾、西部沿岸的密度较高,多样性及均匀性较低,贡湖湾、南部沿岸、东部沿岸、东太湖的密度相对较低,多样性及均匀性相对较高(图2~图3)。其中,竺山湖浮游植物个体密度最高,为3.63×106~1.11 ×109cells/L,平均达2.23 ×108cells/L,多样性及均匀性指数平均分别为1.32、0.37。东太湖的密度相对最低,平均为2.09×106cells/L,多样性及均匀性指数平均分别为1.61、0.59。物种组成上,湖滨带浮游植物群落以蓝藻为主,平均占总密度的88.2%,各湖区排序为梅梁湾(96.8%)>南部沿岸(96.6%)>西部沿岸(95.9%)>竺山湖(90.0%)>贡湖湾(89.9%)>东部沿岸(77.6%)>东太湖(77.5%)。
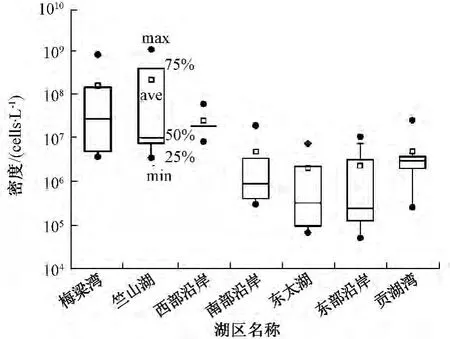
图2 各湖区浮游植物个体密度分布
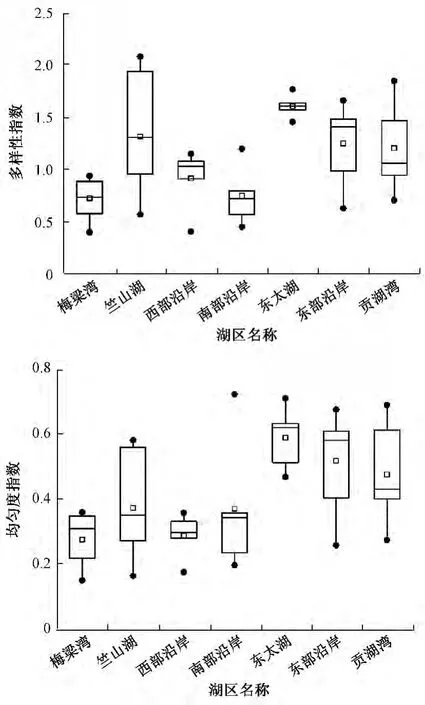
图3 各湖区浮游植物群落多样性
从浮游植物优势类群演替特征(表2)来看,太湖湖滨带浮游植物枯水期(2009年12月)共18种优势种,其中5种蓝藻,6种绿藻,5种硅藻,2种隐藻,各优势种优势度均较低,相对较为均衡,平均优势度前三位均为蓝藻,分别为鱼腥藻(Anabaena sp.)、微囊藻(Microcystis sp.)、颤藻(Oscillatoria sp.);平水期(2010年4月)共16种优势种,其中4种蓝藻,5种绿藻,5种硅藻,2种隐藻,鱼腥藻仍为第一优势种,但优势度(y=0.48 ±0.28)及优势频率(φ =85.71%)较冬季明显上升,绿藻门四尾栅藻(Scenedesmus quadricauda,y=0.13 ±0.09,φ =73.47%)、四角十字藻(Crucigenia quadrata,y=0.13 ±0.09,φ=73.47%)成为第二、三优势种;丰水期(2010年8月)共13种优势种,其中7种蓝藻,4种绿藻,2种隐藻,蓝藻优势明显,微囊藻成为绝对优势种(y=0.63 ±0.29,φ =85.71%)。可以看出,从枯水期到丰水期,太湖湖滨带浮游植物优势类群均以蓝藻为主,第一优势种呈现“鱼腥藻-鱼腥藻-微囊藻”的演替规律,优势度及优势频率逐渐升高,物种逐渐单一,与太湖湖体较为相似[6,9,19]。
2.3 排序分析及关键环境变量指示种筛选
浮游植物群落数据DCA分析结果表明第一排序轴轴长较短(1.88),环境梯度较小,因此,RDA适用于水质指标与浮游植物群落的排序分析。通过蒙特卡罗置换检验,最终筛选出在显著水平能够最大程度解释太湖湖滨带浮游植物群落结构的环境解释变量组合为 TN、CODMn、SS、pH、SD(表3),5个变量共同解释了31.1%的群落演替。图4为监测点位与解释变量的二维排序图。
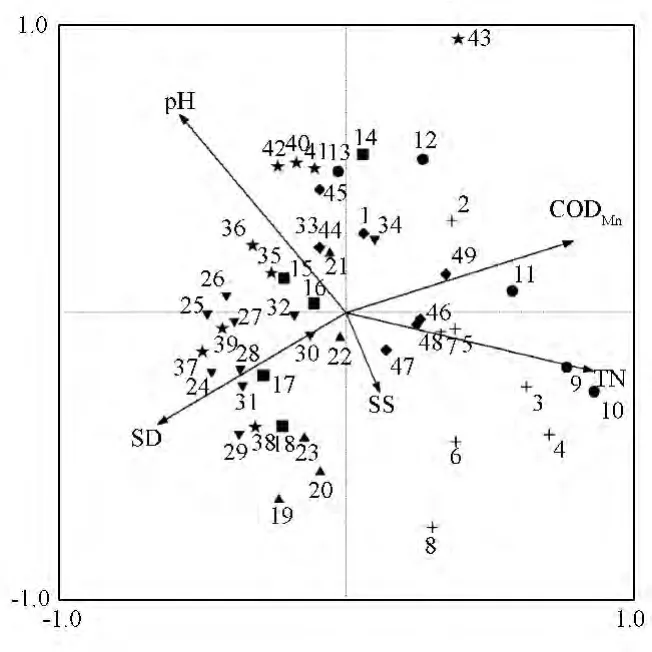
图4 点位与水质因子的RDA第Ⅰ、Ⅱ轴排序图
由图4可见,第Ⅰ轴、第Ⅱ轴的特征值分别为0.15、0.06,与物种-环境变量的相关系数分别为0.82、0.80,分别解释了物种-环境变异的57.4%、22.0%。可以看出,筛选出的环境解释变量组合对湖滨带浮游植物群落具有较好的区分度。
将5个解释变量对浮游植物群落结构变异的解释量进行方差分解,结果表明,TN是影响太湖湖滨带浮游植物分布的关键解释变量,解释了11.7%的群落演替,CODMn、SS、pH 以及 SD 分别解释了 4.0%、3.1%、3.8%和 3.2%的群落演替,5个解释变量之间交互作用解释了5.3%的群落演替。在点位与浮游植物物种的排序图中,只显示适合度介于50% ~100%的物种,筛选发现四尾栅藻(Scenedesmusquadricauda)及弓型藻(Schroederia sp.)与解释变量具有较高的适合度。针对关键解释变量TN分别作四尾栅藻及弓型藻的RDA等值线图(图5a、b),同时将反映太湖富营养化水平的另外2个参数(TP、TSI)也作为解释变量分别作等值线图(图5c、d)。
由图5可以发现,太湖湖滨带的四尾栅藻及弓型藻等值线与TN呈较好的梯度变化,与TN、TP、TSI的组合变量也呈较好的梯度变化,其中四尾栅藻丰度随着TN以及组合变量的升高而降低,弓型藻丰度随着TN以及组合变量值的升高而增加。
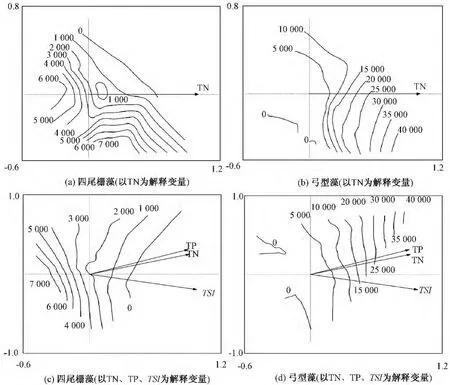
图5 四尾栅藻和弓型藻物种丰度RDA等值线
3 讨论
3.1 湖滨带与湖体浮游植物群落结构比较
2009、2010 年《太湖健康报告》[20-21]显示,太湖湖体浮游植物群落的物种组成基本以蓝藻、绿藻、硅藻为主,其中绿藻、硅藻分类单元数相对较多,蓝藻分类单元相对较少,而在数量上蓝藻占绝对优势,以微囊藻为优势类群。秦伯强等[8]、钱奎梅等[9]在太湖浮游植物群落的多年演替研究上也得到类似结论,太湖湖体在20世纪90年代之后便呈现出微囊藻为优势的特征。可以看出,太湖湖体与湖滨带的浮游植物群落组成以及优势类群相似。
在浮游植物密度以及水华空间分布方面,湖滨带与湖体也有较好的相似性。竺山湖、梅梁湖及西部沿岸为蓝藻水华暴发高频区域,密度较高,易发“湖泛”;属于草型湖泊的南部湖区、东太湖以及东部湖区浮游植物密度明显较低。但是,由于受到风力、水流、湖滨水生植物等因素作用[4,22],浮游植物上浮聚集形成表面水华后比较容易在湖滨堆积,因此相同湖区湖滨带的浮游植物密度比湖体高。周涛等[12]2010年夏季在竺山湖监测到最大浮游植物密度值为1.27×108cells/L,沈爱春等[23]2010年夏季在西部沿岸湖区监测到最大浮游植物密度值4.56×108cells/L,而研究于2010年8月监测到湖滨带浮游植物密度最大值出现在竺山湖西岸,达3.32×109cells/L,相比于湖体基本要高1个数量级。
3.2 浮游植物的环境指示
沈韫芬等[24]研究指出,早在1908年Kolkwitz和Marrson就提出了污水生物系统的概念,列举了不同污染带的“指示生物”,经过百年的发展,美国和欧洲已将生物指标广泛应用于环境管理。然而,国内的水环境管理以水质目标管理为主,“指示生物”方面的研究较少,并且考虑到生物存在明显的区域差异,直接引用国外成果往往会出现“水土不服”。因此,筛选对主要污染因子具有指示作用的本土物种对区域环境管理具有重要的现实意义。利用多元回归方法寻找生物群落与环境因子的关系已经得到广泛应用[5,11-12,19,23,25],可以有效地找出影响群落结构变化的最小环境变量解释组合。吴东浩等[25]利用RDA排序方法在西苕溪筛选出了影响西苕溪底栖动物分布的首要污染因子为氨氮,并找出了与氨氮浓度匹配较好的指示物种。针对太湖湖滨带浮游植物群落以及水体富营养化问题,开展“群落-环境”的排序分析,发现与湖体氮的分布情况匹配度最高的为绿藻,分别是四尾栅藻以及弓型藻某种。这2种藻在太湖流域分布较为广泛,属常见种,具备一定的适用性。四尾栅藻是典型的富营养指示种,相关研究报道较多。郑晓宇等[26]通过室内培养实验研究发现,四尾栅藻的生长受氮磷的影响明显,喜高氮磷的水体,现存量与氮磷浓度变化具有较好的线性相关关系。与太湖优势种铜绿微囊藻的竞争关系研究发现,在不同光照条件[27]、氮磷浓度[28]、温度条件[29]下两者之间存在明显的竞争抑制关系。对应于目前太湖夏季的光照、温度及营养条件,铜绿微囊藻对四尾栅藻的抑制作用较明显,这也印证了图5中显示的湖滨带水体TN、TP浓度及TSI与四尾栅藻丰度呈负相关关系的结论。在实际的水体中,生物之间的竞争以及他感等作用相对实验室的条件实验研究显得更为复杂,四尾栅藻对太湖湖滨带富营养化的定量化指示意义尚不明确,弓形藻与富营养化的关系方面尚未有相关报道,均有待进一步研究。
4 结论
太湖湖滨带浮游植物群落以蓝藻为优势类群,多样性整体较低。从枯水期到丰水期,物种逐渐单一化,微囊藻优势明显。空间上,竺山湖、梅梁湾以及西部沿岸湖滨带的藻密度相对较高,东部沿岸、东太湖及南部沿岸相对较低。总体而言,太湖湖滨带浮游植物群落结构与湖体相似,密度比湖体高1个数量级。
通过RDA排序分析,筛选出在显著水平上最大程度解释湖滨带浮游植物分布的最小解释变量组合为 TN、CODMn、SS、pH、SD,且方差分解分析结果显示TN是相对最重要的环境变量。
以TN为环境变量,作环境与物种变量等值线图,在适合度50% ~100%范围内,与TN具有较好梯度响应关系的物种为四尾栅藻和弓型藻。这2个种在太湖均为常见种,并且与TN、TP、TSI的组合变量也有较好的梯度响应关系,对太湖湖滨带的富营养化具有一定的指示意义。但两者与太湖湖滨带污染胁迫的定量响应关系尚不明朗,对富营养化的定量指示意义需进一步研究。
致谢:研究中,太湖监测点位水化学数据由中国环境科学研究院湖泊工程技术中心叶春研究员团队提供,谨致谢忱。
[1]李春华,叶春,赵晓峰,等.太湖湖滨带生态系统健康评价[J].生态学报,2012,32(12):3 806-3 815.
[2]叶春.洱海湖滨带生态恢复工程模式研究[D].北京:中国环境科学研究院,1999.
[3]孙顺才,黄漪平.太湖[M].北京:海洋出版社,1993:3-9.
[4]叶春,李春华,王秋光,等.大堤型湖滨带生态系统健康状态驱动因子-以太湖为例[J].生态学报,2012,32(12):3 681-3 690.
[5]Marchetto A,Padedda B M,Mariani M A,et al.A numerical index for evaluating phytoplankton response to changes in nutrient levels in deep mediterranean reservoirs[J].Journal of Limnology,2009,68(1):106-121.
[6]Chen Y,Qin B,Teubner K,et al.Long-term dynamics of phytoplankton assemblages:Microcystis-domination in Lake Taihu,a large shallow lake in China[J].Journal of Plankton Research,2003,25(4):445-453.
[7]Chen Y,Fan C,Teubner K,et al.Changes of nutrients and phytoplankton chlorophyll-a in a large shallow lake,Taihu,China:an 8-year investigation[J].Hydrobiologia,2003,506(1-3):273-279.
[8]秦伯强,胡维平,陈伟明.太湖水环境演化过程与机理[M].北京:科学出版社,2004:255-266.
[9]钱奎梅,陈宇炜,宋晓兰.太湖浮游植物优势种长期演化与富营养化进程的关系[J].生态科学,2008,27(2):65-70.
[10] Xu H,Paerl H W,Qin B Q,et al.Nitrogen and phosphorus inputs control phytoplankton growth in eutrophic Lake Taihu,China[J].Limnology and Oceanography,2010,55(1):420-432.
[11]苏玉,文航,王东伟,等.太湖武进港区域浮游植物群落特征及其主要水质污染影响因子分析[J].环境科学,2011,32(7):1 945-1 951.
[12]周涛,李正魁.太湖浮游植物与营养盐相互关系[J].农业环境科学学报,2013,32(2):327-332.
[13]胡鸿均,魏印心.中国淡水藻类-系统,分类及生态[M].北京:科学出版社,2006.
[14] Komárek J,MaresˇJ.An update to modern taxonomy(2011) of freshwater planktic heterocytous cyanobacteria[J].Hydrobiologia,2012,698(1):327-351.
[15]HJ 494—2009 水质采样技术指导[S].
[16]王明翠,刘雪芹,张建辉.湖泊富营养化评价方法及分级标准[J].中国环境监测,2002,18(5):47-49.
[17]Lampitt R S,Wishner K F,Turley C M,et al.Marine snow studies in the northeast Atlantic:distribution,composition and roles as a food source for migrating plankton[J].Marine Biology,1993,116:689-702.
[18]GB/T 3838—2002 地表水环境质量标准[S].
[19]冯露露,李正魁,周涛.太湖浮游植物和各形态无机氮的时空分布特征[J].湖泊科学,2012,24(5):739-745.
[20]太湖流域管理局.太湖健康状况报告(2009)[EB/OL].(2010-09-29)[2014-09-01].http://www.tba.gov.cn//tba/content/TBA/lygb/thjkzkbg/index.html.
[21]太湖流域管理局.太湖健康状况报告(2010)[EB/OL].(2013-04-03)[2014-09-01].http://www.tba.gov.cn//tba/content/TBA/lygb/thjkzkbg/index.html.
[22]王成林,张咏,张宁红,等.太湖藻源性“湖泛”形成机制的气象因素分析[J].环境科学,2011,32(2):401-408.
[23]沈爱春,徐兆安,吴东浩.太湖夏季不同类型湖区浮游植物群落结构及环境解释[J].水生态学杂志,2012,33(2):43-47.
[24]沈蕴芬,章宗涉,龚循矩,等.微型生物监测新技术[M].北京:中国建筑工业出版社,1990:120-139.
[25]吴东浩,张勇,于海燕,等.影响浙江西苕溪底栖动物分布的关键环境变量指示种的筛选[J].湖泊科学,2010,22(5):693-699.
[26]郑晓宇,顾詠洁,金妍,等.不同氮、磷质量浓度下四尾栅藻的生长研究[J].生态环境学报,2010,19(11):2 663-2 668.
[27]胡小贞,金相灿,储昭升,等.太湖铜绿微囊藻与四尾栅藻的光竞争及模拟优势过程初探[J].农业环境科学学报,2005,24(3):538-543.
[28]万蕾,朱伟,赵联芳.氮磷对微囊藻和栅藻生长及竞争的影响[J].环境科学,2007,28(6):1 230-1 235.
[29]郑忠明,白培峰,陆开宏,等.铜绿微囊藻和四尾栅藻在不同温度下的生长特性及竞争参数计算[J].水生生物学报,2008,32(5):720-727.
