内皮素3在不同毛色绵羊皮肤中的差异表达
2015-03-22杨姗姗白俊明范瑞文耿建军焦丁兴董常生
杨姗姗,白俊明,范瑞文,耿建军,焦丁兴,周 英,董常生*
(1.山西农业大学动物科技学院,太谷 030801;2.河北省清河县农业局,清河 054000)
内皮素3在不同毛色绵羊皮肤中的差异表达
杨姗姗1#,白俊明1#,范瑞文1,耿建军1,焦丁兴2,周 英1,董常生1*
(1.山西农业大学动物科技学院,太谷 030801;2.河北省清河县农业局,清河 054000)
本研究旨在探讨内皮素3在绵羊皮肤毛色中的作用。采用实时荧光定量PCR、Western blotting和免疫组织化学对内皮素3在黑色和白色绵羊皮肤中的表达差异进行分析。实时荧光定量PCR结果显示,内皮素3在黑色绵羊皮肤中的相对表达量较高,是其在白色绵羊皮肤中的表达量的2.89倍,且差异极显著(P<0.01);Western blotting结果显示,内皮素3在黑色绵羊皮肤总蛋白中的表达量极显著高于其在白色绵羊皮肤总蛋白中的表达量(P<0.01);免疫组织化学结果表明,内皮素3在白色绵羊皮肤毛囊的毛乳头上方、毛球底部及外根鞘呈阳性表达,而在黑色绵羊皮肤毛囊的毛球底部和部分外根鞘呈阳性表达。综上表明,内皮素3可能参与绵羊毛色形成。
内皮素3;皮肤;绵羊
内皮素(Endothelin,Edn)是由21个氨基酸组成的多肽,存在于血管内皮和各种组织及细胞中,参与心血管功能的调节,在维持基础血管张力与心血管系统稳态方面有着重要作用。内皮素包含3种异构体:Edn1、Edn2和Edn3[1]。在这3种内皮素异构体中,Edn3对来源于神经脊(NC)细胞的黑素细胞的发育有着主要作用[1]。在胚胎发生过程中,Edn3不仅控制起源神经嵴黑素细胞转化的开始并可以维持其表型;在体外Edn3可以促进培养的正常的人体黑色素细胞、痣细胞和黑色素瘤细胞的生长和迁移[2]。越来越多的研究表明:Edn3在促进黑色素细胞增殖的同时对黑色素的合成也有重要的调节作用。另有研究证明内皮素信号在黑色素细胞的发育和存活的过程中有着至关重要的作用[3]。目前,人们对Edn3在黑色素细胞中黑色素生成和皮肤色素沉积的研究仅仅局限在体外对人、鼠[4-6]、鹌鹑[7]等少数动物,而对毛用动物体内的研究甚少。
绵羊是一种经济价值较高的毛用动物,毛色是绵羊的重要性状之一,化学染色是羊毛实现色彩多样性的主要手段,对人类健康以及环境保护产生不良影响。因此,从分子水平改变绵羊毛色将具有重要的意义,而发掘毛色基因及其功能将为此提供理论依据。动物毛色由黑色素的含量与分布决定,由多个基因调控[8]。关于Edn3在绵羊不同毛色形成过程中的作用未见报道,因此,本试验以白色和黑色绵羊为研究对象,对Edn3在皮肤中的表达进行研究。
1 材料与方法
1.1 材料
选取白色和黑色成年雄性绵羊各3只。选取绵羊背侧皮肤,净毛后,用取皮器取绵羊皮肤组织3块(0.8 cm2)。其中两块置于液氮中,分别用于实时荧光定量PCR和Western blotting; 1块置于4% 的Bouin’s固定液中,固定24 h,石蜡包埋,切成5 μm的切片,用于免疫组织化学试验。
1.2 主要试剂
兔抗内皮素3多克隆抗体(博奥森),山羊抗兔二抗(康为世纪),DAB显色试剂盒(武汉博士德),Trizol Reagent(Invitrogen),2×Es Taq MasterMix(康为世纪),DL2000 DNA Marker(TaKaRa),反转录试剂盒(TaKaRa),SYBR®PrimeSciptTM RT-PCR Kit(Life)。
1.3 引物设计
通过对GenBank上的各个物种Edn3序列进行Blast后,获取保守序列并利用Primer5.0引物设计软件,设计荧光定量PCR扩增引物,通过NCBI初步检测引物的特异性。引物由华大基因公司合成。引物见表1。
表1 目的基因引物序列和退火温度
Table 1 Primer sequences and annealing temperature of PCR amplification of target genes

目的基因Targetgene引物序列(5′⁃3′)Primersequence产物/bpProductsEdn3F:TTTGCCTCTTCTTCCTTA,R:TATTCTCGGTAGTGTCCC163β⁃actinF:CCTGACCGGGAAGAGGAATC,R:AAAACAGGGGGTTGAACTC150
1.4 RNA提取和cDNA合成
用Trizol法提取总RNA。各取白色和黑色绵羊皮肤总RNA 1 μL,快速电泳检测其质量。
按照TaKaRa公司反转录试剂盒进行绵羊皮肤cDNA合成,反应体系:Oligo dT(50 μmol·L-1)或Random 6 mers(50 μmol·L-1)1 μL,dNTP Mixture(10 mmol·L-1each)1 μL,总RNA ≤5 μg,用RNase free水补足至10 μL。反应条件:65 ℃ 5 min,反应结束,立即置于冰上冷却。在此体系中再加入5×PrimeScript II Buffer 4 μL,RNase Inhibitor(40 U·μL-1)0.5 μL,PrimeScript II RTase(200 U·μL-1)1 μL,用RNase free水补足至20 μL。反应条件:42 ℃30~60 min,95 ℃5 min。反应结束后,置于冰上冷却,-20 ℃保存备用。
按照Life试剂盒进行实时荧光定量PCR,每个样本设3次重复,依次在冰上加样:SYBR®Select Master Mix 10 μL,上下游引物(10 μmol·L-1)各0.4 μL,cDNA 1 μL,用RNase free水补足至20 μL。反应条件:95 ℃30 s;95 ℃5 s,55 ℃30 s,72 ℃10 s,40个循环,内参基因同样反应条件。反应结束,由熔解曲线判定PCR反应的特异性,根据标准曲线以及荧光曲线的Ct值计算定量结果。数据用SPSS16.0软件,应用方差分析进行生物统计学分析。不同样本的2-△△CT法[9]用单因子方差分析,分析结果用“平均值± 标准误(Means±SE)”表示,内皮素3 mRNA的表达差别倍数用2-△△CT表示。
1.5 免疫组织化学
固定24 h后的绵羊皮肤进行脱水透明后,在石蜡中进行包埋,然后切片烘干进行免疫组织化学试验。试验步骤:石蜡切片常规脱蜡、脱水,每张切片滴加一滴3%H2O2,置于37 ℃烘箱中,20 min用于灭活内源性过氧化物酶[8];PBS清洗3次,滴加5%牛血清白蛋白,37 ℃烘箱放置20 min进行封闭,甩掉封闭液,滴加1∶100兔抗Edn3多克隆抗体,4 ℃过夜;取出后,25 ℃反应30 min,PBS清洗3次。滤纸吸干PBS,滴加二抗,37 ℃孵育30 min,PBS清洗3次。滴加DAB显色试剂,显色5 min,根据显色情况终止显色。脱水、透明后,用中性树胶封片。显微镜下观察结果。用PBS取代一抗作为阴性比照,对染色结果的阳性产物进行分析。
1.6 总蛋白的提取与Western blotting检测
提取液氮内剩余的绵羊皮肤组织的总蛋白。用BCA法测定蛋白浓度,每孔总蛋白上样量为100 μg,进行聚丙烯酰胺凝胶电泳,转移至PVDF膜,PVDF膜用5%脱脂奶粉封闭,室温封闭1.5~2.0 h;将PVDF膜放入含有稀释好的一抗的平皿中,4 ℃摇床过夜孵育;用TBST洗膜,每次10 min,共3次;加入二抗,37 ℃ 摇床孵育30 min,BST洗膜,加入ECL发光剂[9],用显影剂和定影粉进行胶片曝光处理。用Image proplus 6.0软件(Olympus)对绵羊皮肤组织总蛋白中的Edn3和β-actin免疫印迹结果进行半定量分析:蛋白总量=条带面积×灰度值,Edn3与内参蛋白β-actin对比。数据用SPSS16.0软件进行生物统计学分析,不同样本的光密度用单因子方差分析,所得结果用“平均值±标准误(Means±SE)”表示。
2 结 果
2.1Edn3基因的mRNA表达结果
从扩增动力学曲线上看,本研究结果符合标准的“S”形荧光增长曲线,目的基因Edn3和内参基因β-actin的扩增动力学曲线整体平行性较好,基线无明显上扬趋势(图1A)。从扩增熔解曲线上看,曲线为单峰,表明在实时荧光定量PCR过程中,没有非特异性的扩增产物,荧光强度均来于特异性的扩增产物,目的基因Edn3及内参基因β-actin没有非特异性扩增及引物二聚体产生(图1B)。结果表明,在mRNA水平,Edn3基因在黑色绵羊皮肤中的相对表达量是白色绵羊皮肤的2.89倍。且二者表达差异极显著(P<0.01)(图2)。
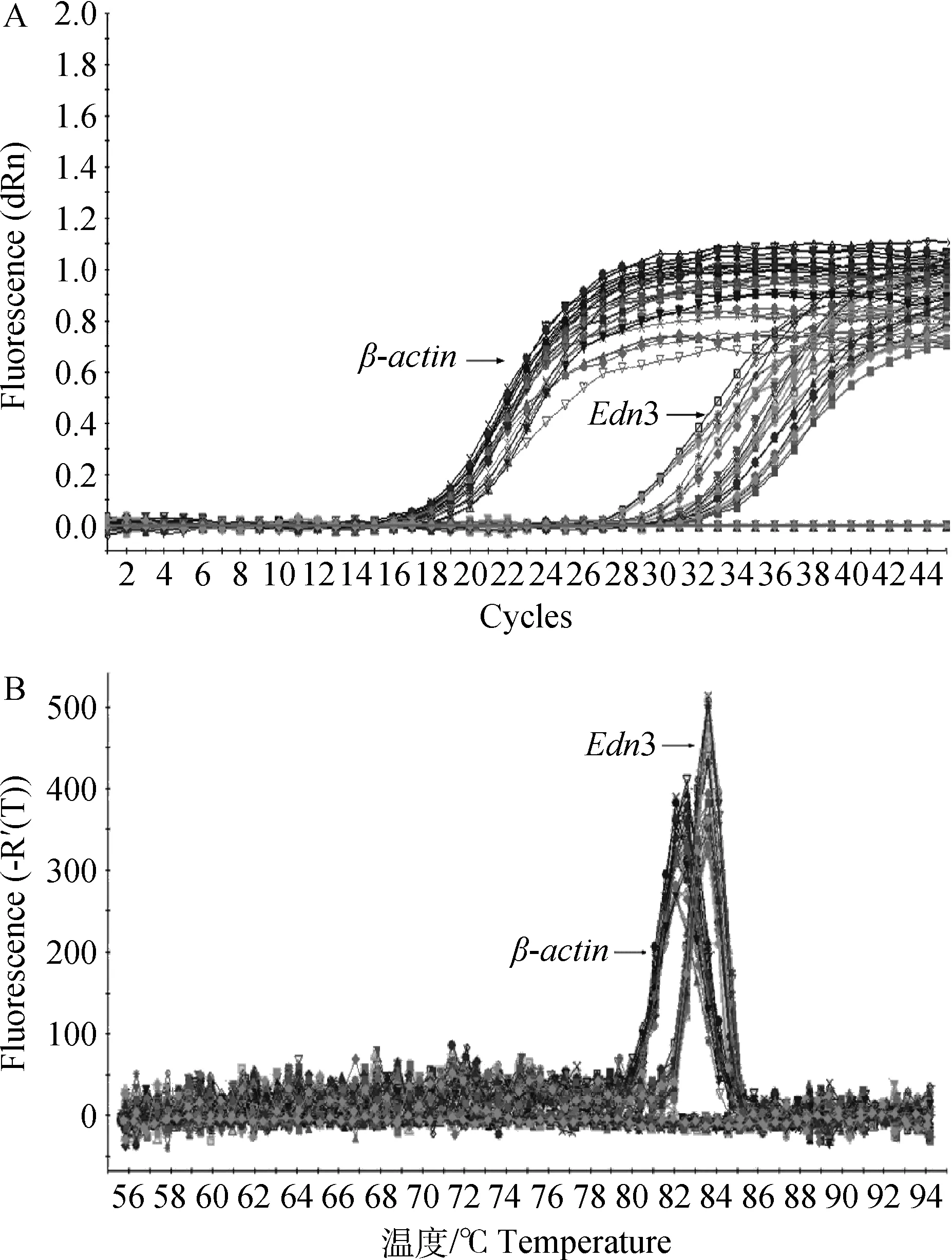
图1 Edn3基因和内参基因的扩增动力学曲线(A)和熔解曲线(B)Fig.1 PCR amplification plot(A)and dissociation curve(B) of Edn3 and β-actin gene

**.P<0.01。下同**.P<0.01.The same as below图2 Edn3 mRNA在不同毛色绵羊皮肤中的表达Fig.2 Edn3 mRNA expression in sheep skins with different hair color
2.2 绵羊皮肤中Edn3蛋白表达结果
Western blotting结果显示,绵羊皮肤总蛋白中存在与兔抗Edn3多克隆抗体发生免疫阳性反应的蛋白条带,且分子质量符合实际大小。对不同毛色的绵羊皮肤内Edn3蛋白表达进行定性分析。从图中可以看出在黑色绵羊皮肤中,Edn3蛋白水平表达较显著,而在白色绵羊皮肤中检测到的信号较弱(图3A)。应用Inage proplus 6.0软件对Edn3、内参β-actin的免疫印迹进行半定量分析,结果显示,Edn3蛋白在白色绵羊皮肤中的平均蛋白含量为0.18±0.074;在黑色绵羊皮肤中的平均蛋白含量为1.47±0.12,两者差异极显著(P<0.01)(图3B)。
2.3 绵羊Edn3蛋白在皮肤组织中的分布
免疫组织化学染色结果显示,Edn3在白色绵羊皮肤和黑色绵阳皮肤的毛囊中均有免疫阳性反应,但不同毛色的绵羊皮肤毛囊其表达部位不同(图4)。在白色绵羊中(图4A、B),Edn3免疫阳性反应细胞分布在皮肤毛囊的毛乳头上方,毛球底部和外根鞘;在黑色绵羊中(图4C、D),Edn3免疫阳性细胞主要分布在皮肤毛囊的毛球底部和部分外根鞘。

A.1~3、4~6.白色和黑色绵羊皮肤总蛋白中的阳性印迹信号;B.印迹条带的平均光密度分析。内参为β-actin,数据用 “平均值±标准误”表示A.The bands were the positively blotting signal with the rabbit Edn3 polyclonal antibody and goat anti-rabbit IgG conjugated HRP as the second antibody.1-3 from sheep skins with white hair color and 4-6 from sheep skins with black hair color;B.The average intensity of blotting band was analyzed.The level of protein was normalized relative to β-actin.Bars in each panel represent the mean ± standard error图3 Edn3蛋白在绵羊皮肤中的表达Fig.3 Edn3 protein expression in sheep skin
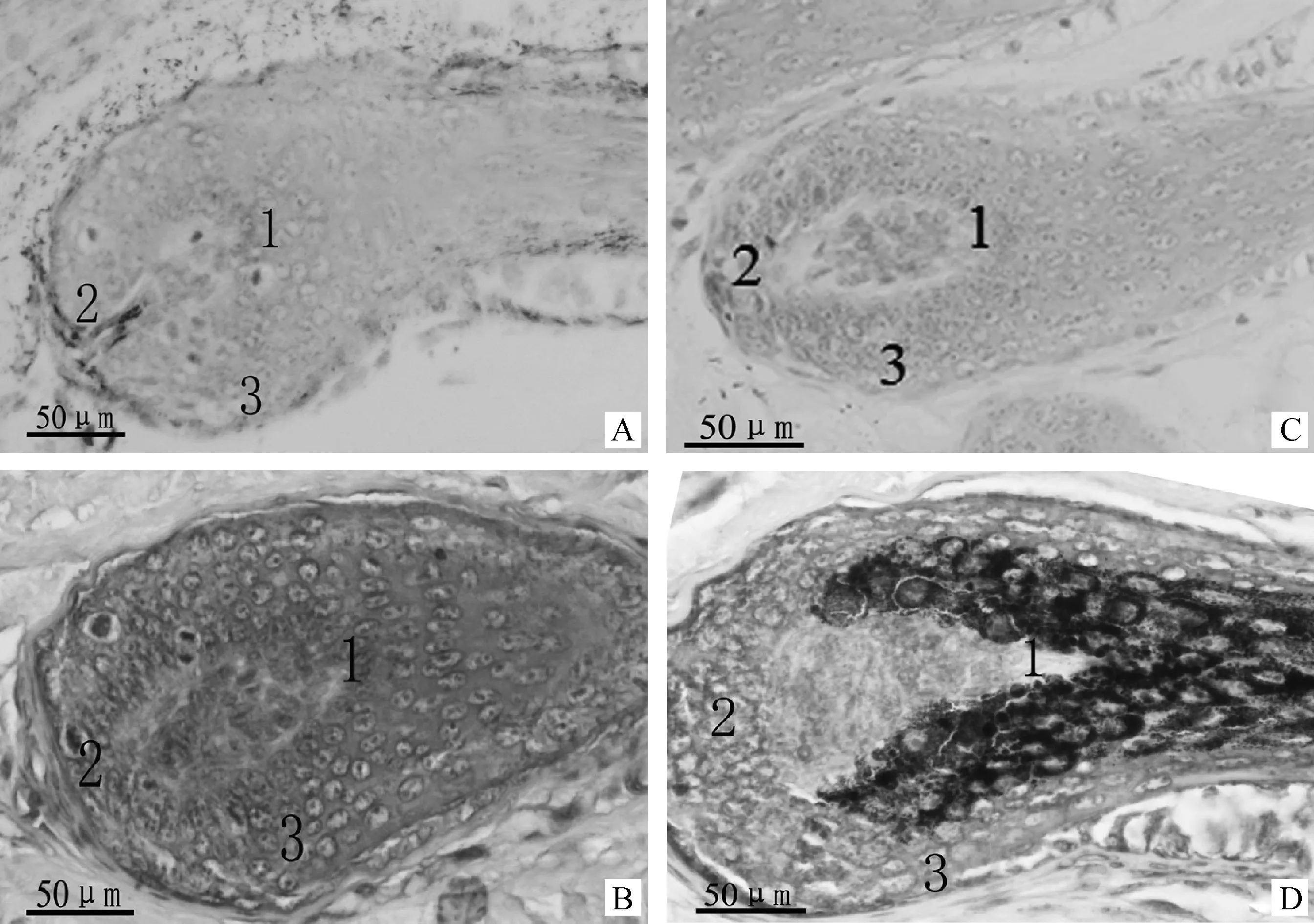
A和B.Edn3在白色绵羊皮肤中的定位;C和D.Edn3在黑色绵羊皮肤中的定位;A和C.阴性对照;B和D.试验组。1.毛乳头上方;2.毛球;3.外根鞘A,B.Edn3 localization in sheep skin with white hair color;C,D.Edn3 localization in sheep skin with black hair color.A,C.Negative control;B,D.Experiment groups;1.Upper hair dermal papilla;2.Hair bulb;3.Outer root sheath图4 Edn3蛋白在绵羊皮肤中的定位40×Fig.4 Localization of Edn3 protein in sheep skin 40×
3 讨 论
哺乳动物被毛的颜色主要是由沉着在皮肤内黑色素的量及其种类决定的,黑色素主要包括真黑色素(Eumelanin)和褐黑色素(Pheomelanin)两类,真黑色素和褐黑色素的相对数量决定了动物皮肤与被毛颜色[10]。黑色素细胞是哺乳动物毛色形成的基础,在胚胎发育过程中黑色素细胞由神经嵴细胞分化并定向迁移到全身各个组织,其定向分化、迁移与MITF、FGF-2、内皮素以及诸多因子有关[11]。
内皮素是参与黑色素细胞生物学过程的主要旁分泌因子之一,在其早期发育过程中起着重要作用[12]。本研究发现,在白色绵羊皮肤毛囊的毛乳头上方、毛球底部及外根鞘和黑色绵羊皮肤毛囊的毛球底部及外根鞘均存在Edn3阳性免疫反应物。而毛球是向毛干提供黑色素的部位,其含有大量黑色素细胞,胞质中含有黑色素颗粒[13]。在哺乳动物黑色素细胞中,G蛋白偶联内皮素受体B(Ednrb) 途径功能的丢失或突变都会导致胚胎中黑色素细胞前体的数量减少,从而引起毛色的改变[14]。Edn3是Ednrb的配体[15]。说明长期的Edn3通过激活Ednrb2可以促进黑色素细胞的分化。另有资料报道,Ednrb与MITF具有一定的机制关联性[16],MITF 可结合到与黑色素合成有主要关系的TYRP2 启动子区域来控制它们的转录,TYRP2可以直接参与黑色素生成调控过程,调节黑色素细胞的发育、生存和功能。证明Edn3结合Wnt3a 和 Kit-配体(Kitl、SLF和SCF)诱导人胚胎干细胞表达生黑素标记物,Mitf 和Sox10,产生黑色素小体,从而产生黑色素[17]。因此,笔者认为Edn3的表达差异使Ednrb和KIT差异性调控MITF的表达,进而调控黑色素合成路径上TYRP2和TYR的表达,导致黑色绵羊皮肤中色素含量升高。本试验结果也发现黑色绵羊皮肤中Edn3基因的表达水平显著高于白色绵羊,可能支持Edn3与毛色相关的结论。
在体内,缺失Ednrb或Kit信号肽都会引起部分成黑色素细胞的丢失,Edn3与Kit1共同作用可以促进黑色素细胞的增殖、分化和存活。只有在Edn3和Kit1同时存在的条件下,才会引起成黑素细胞数量的显著增加和产生具有功能完全的黑色素细胞[12]。KIT的激活作用可以快速引起MAPK介导的MITF磷酸化,与激活因子家族相互作用进而调控MITF的转录活性,从而调控黑色素沉着。但Edn3蛋白是否在毛囊的黑色素细胞中通过上调KIT和Ednrb表达,促使KIT和Ednrb与MITF作用,上调毛囊黑色素细胞中MITF蛋白的表达,参与绵羊毛色的形成,有待于进一步研究。
4 结 论
Edn3定位于绵羊皮肤毛囊中黑色素细胞分布的部位,且在不同毛色的绵羊皮肤中存在差异性表达,结果说明Edn3与绵羊毛色形成过程具有一定的相关性。
[1] AMY S,LIDIA K.Roles of endothelin signaling in melanocyte development and melanoma[J].PigmentCellMelanomaRes,2010,23(2):160-170.
[2] REID K,TURNLEY A M,MAXWELL G D,et al.Multiple roles for endothelin in melanocyte development:regulation of progenitor number and stimulation of differentiation[J].Development,1996,122(12):3911-3919.
[3] 耿建军,白俊明,范瑞文,等.内皮素-3对羊驼黑色素细胞特征及细胞内毛色基因表达的影响[J].畜牧兽医学报,2013,44(7):1070-1077. GENG J J,BAI J M,FAN R W,et al.The effects of endothelin-3 on the melanocyte characteristic and expression of hair color genes in melanocytes from alpaca(Lamapacos)invitro[J].ActaVeterinariaetZootechnicaSinica,2013,44(7):1070-1077.(in Chinese)
[4] KADONO S,MANAKA I,KAWASHIMA M,et al.The role of the epidermal endothelin cascade in the hyperpigmentation mechanism of lentigo senilis[J].JInvestDermatol,2001,116(4):571-577.
[5] MANAKA L,KADONO S,KAWASHIMA M,et al.The mechanism of hyperpigmentation in seborrhoeic keratosis involves the high expression of endothelin converting enzyme1 alpha and TNF alpha,which stimulate secretion of endothelin1[J].BrDermatol,2001,145:895-903.
[6] OKAZAKI M,YOSHIMURA K,UCHIDA G,et al.Epidermal hyperpigmentation in non-syndromic solitary cafe-au-lait macules may be associated with increased secretion of endothelin-1 by lesional keratinocytes[J].ScandJPlastReconstrSurgHandSurg,2005,39(4):213-217.
[7] LAHAV R,DUPIN E,LECOINE L,et al.Endothelin-3 selectively promotes survival and proliferation of neural crest-derived glial and melanocytic precursorsinvitro[J].ProcNatlAcadSciUSA,1998,95:14214-14219.
[8] 张俊珍,范瑞文,杨姗姗,等.羊驼皮肤中可溶性鸟苷酸环化酶的表达差异[J].畜牧兽医学报,2013,44(7):1057-1062. ZHANG J Z,FAN R W,YANG S S,et al.The different expression of soluble guanylate cyclase in alpaca skin[J].ActaVeterinariaetZootechnicaSinica,2013,44(7):1057-1062.(in Chinese)
[9] 张丹丽,田 雪,范瑞文,等.APC在羊驼皮肤组织中的表达和定位分析[J].畜牧兽医学报,2014,45(2):327-332. ZHANG D L,TIAN X,FAN R W,et al.Expression and location analysis of APC in alpaca skin[J].ActaVeterinariaetZootechnicaSinica,2014,45(2):327-332.(in Chinese)
[10] SIMON J D,PELES D N.The red and the black[J].AccChemRes,2010,43(11):1452-1460.
[11] 耿建军,穆晓丽,孙乐天,等.内皮素受体B在不同毛色羊驼皮肤中的表达与定位[J].畜牧兽医学报,2010,41(11):1478-1484. GENG J J,MU X L,SUN L T,et al.Expression and immunolocalization of endothelin receptor B in alpaca skin of different colors[J].ActaVeterinariaetZootechnicaSinica,2010,41(11):1478-1484.(in Chinese)
[12] SALDANA-CABOVERDE A,KOS L.Roles of endothelin signaling in melanocyte development and melanoma[J].PigmentCellMelanomaRes,2010,23(2):160-170.
[13] SLOMINSKI A,WORTSMAN J,PLONKA P M,et al.Hair follicle pigmentation[J].JInvestDermatol,2005,124(1):13-21.
[14] 张 晖,戈婷婷,彭沙沙,等.泰和乌骨鸡皮肤黑色素与多泡脂肪细胞的超微结构研究[J].畜牧兽医学报,2014,45(5):815-820. ZHANG H,GE T T,PENG S S,et al.Ultrastructural investigation on melanin and multilocular adipocyte in skin of taihe black-bone fowl[J].ActaVeterinariaetZootechnicaSinica,2014,45(5):815-820.(in Chinese)
[15] LAHAV R.Endothelin receptor B is required for the expansion of melanocyte precursors and malignant melanoma[J].IntJDevBiol,2005,49(2-3):173-180.
[16] KAYO S,EMI K N,EIJIRO A,et al.Epistatic connections between microphthalmiaassociated transcription factor and endothelin signaling in Waardenburg syndrome and other pigmentary disorders[J].FASEBJ,2008,22:1155-1168.
[17] FANG D,LEISHEARE K,NGUYEN T K,et al.Defining the conditions for the generation of melanocytes from human embryonic stem cells[J].StemCells,2006,24(7):1668-1677.
(编辑 程金华)
The Different Expression of Endothelin 3 in Sheep Skins
YANG Shan-shan1#,BAI Jun-ming1#,FAN Rui-wen1,GENG Jian-jun1,JIAO Ding-xing2,ZHOU Ying1,DONG Chang-sheng1*
(1.CollegeofAnimalScienceandVeterinaryMedicine,ShanxiAgriculturalUniversity,Taigu030801,China;2.QingheCountyAgriculturalBureauofHebeiProvince,Qinghe054000,China)
To investigate the function of endothelin 3(Edn3) in sheep skins and the regulation of hair color formation,the differential expression of Edn3 in skins of black versus white sheep was examined.Quantitative real-time PCR(qRT-PCR),Western blotting and immunohistochemistry methods were performed.The results showed that the quantity expression of Edn3 mRNA in black sheep was 2.89 times than in white sheep(P<0.01);the average protein expression of Edn3 in black sheep skin was higher than in white sheep skin on the optical density(P<0.01).The results of immunohistochemistry revealed that Edn3 was localized in the hair dermal papilla,lower of hair bulb and outer root sheath in hair follicles of sheep with white hair color,but was localized in lower of hair bulb and outer root sheath in hair follicles of sheep with black hair color.The differential expression and localization of Edn3 in skin of white versus black sheep suggests a potential role in hair color regulation.
endothelin3(Edn3);skin;sheep
10.11843/j.issn.0366-6964.2015.11.026
2015-01-08
国家“863”计划(2013AA102506);公益性行业(农业)科研专项(201303119);农业部“948”项目(2011-Z33)
杨姗姗(1987-),女,河北石家庄人,博士生,主要从事羊驼毛色研究,E-mail:shanshan0321@163.com;白俊明(1984-),男,山西忻州人,硕士,主要从事羊驼毛色研究,E-mail:baibai84aa@163.com。杨姗姗与白俊明为并列第一作者
*通信作者:董常生,教授,主要从事生物工程研究,E-mail:cs-dong@sxau.edu.cn
S852.1
A
0366-6964(2015)11-2111-06
