绵羊CYP19基因卵巢启动子的克隆及其表达活性研究
2015-03-22孙洪新张英杰刘月琴陈晓勇敦伟涛
孙洪新 ,张英杰 ,刘月琴,陈晓勇,敦伟涛
(1.河北农业大学动物科技学院,保定 071000; 2.河北省畜牧兽医研究所,保定 071000)
绵羊CYP19基因卵巢启动子的克隆及其表达活性研究
孙洪新1,2,张英杰1*,刘月琴1,陈晓勇2,敦伟涛2
(1.河北农业大学动物科技学院,保定 071000; 2.河北省畜牧兽医研究所,保定 071000)
本研究旨在克隆绵羊CYP19基因卵巢启动子序列片段,构建真核表达载体,并在细胞水平检测其组织特异性表达情况。参考已知序列设计特异性引物,PCR扩增绵羊CYP19基因卵巢启动子1.1和0.5 kb两个片段,并与已公布的序列进行同源性比较;将测序正确的两个片段分别定向克隆到去除CMV的pEGFP-N2载体骨架中,构建真核表达载体pCYP19-1.1-EGFP-N2和 pCYP19-0.5-EGFP-N2,重组质粒在脂质体LipofectamineTMLTX+PLUS介导下,分别转染绵羊的颗粒细胞和胎儿成纤维细胞,并于转染后24、48 和72 h观察EGFP表达情况。结果表明,扩增获得片段与已公布序列高度同源;应用转录因子结合位点预测软件对所得序列分析表明,该扩增片段含有类似于TATA-box核心启动顺式元件,并含有多个潜在转录因子的结合位点。转染24 h后,发现pCYP19-1.1-EGFP-N2在颗粒细胞可观测到绿色荧光表达,48 h荧光细胞数量有所增加;转染72 h时荧光细胞数最多;在胎儿成纤维细胞也有少量EGFP基因表达。pCYP19-0.5-EGFP-N2在颗粒细胞和成纤维细胞中均未检测到荧光。结果表明,扩增所得的绵羊CYP19基因卵巢启动子1.1 kb片段可引导外源基因在颗粒细胞中的表达,可用于与繁殖相关的基因功能及转基因动物研究,但并非卵巢特异性启动子。
绵羊;CYP19;卵巢启动子;表达
CYP19基因属于P450(Aromatase cytochrome P450 gene)超家族基因,是编码芳香化酶P450 arom的基因。芳香化酶P450arom能够催化睾酮和雄烯二酮不可逆的转化为雌激素,是雌激素生物合成中的关键酶和限速酶。该过程在鱼类、两栖类、爬行类、鸟类以及哺乳动物的性腺分化和性器官发育中是必不可少的,同时对脑的发育及功能也具有重要影响[1-3]。
CYP19基因有多个启动子,由它们介导CYP19基因在不同组织中的表达。不同物种CYP19基因启动子的数量和结构有所区别,命名也不相同。人的CYP19基因位于染色体15q21.1区带[4],由10个外显子和9个内含子组成[5]。迄今,已发现了人CYP19基因的10个组织特异性启动子(P1.1、P1.2、P1.3、P1.4、P1.5、P1.6、P1.7、PII、P2a和P1.f)[4-5],均位于外显子2 5′端上游约93 kb 长的外显子1区域[6-7]。绵、山羊和牛CYP19基因分别位于染色体7q24-q31、10q26和q2.6区带,在牛羊也发现了多个组织特异性启动子,其中牛上有7个P1.1、P1.2a、P1.3、P1.4、P1.5、P2、P1.2a/b[8-11];羊CYP19基因启动子P1.1、P1.4 、P1.5和P2[11]。在鼠类中仅发现了两个启动子。
CYP19基因的表达模式是选择性地利用不同启动子和相应的外显子[12],CYP19基因在不同物种和特定组织中的表达是通过选择性的使用不同启动子实现的。不同启动子选择性地把自身外显子1 5′端非翻译区域(5′UTR) 剪接到位于外显子1翻译起始位点(ATG) 上游处的剪切位点(AG/GACT)上,调控 P450arom mRNA水平。但是CYP19 基因编码区是恒定的,无论剪切模式如何,不同组织CYP19基因所编码的芳香化酶蛋白总是相同的[13]。不同启动子的上游基因可以携带不同的调节基因,结合于该区域的核蛋白受组织类型和内分泌环境的影响,物种的差异使基因在表达的各个环节中可能产生或多或少的差异,特定组织中CYP19基因的表达产物芳香化酶P450的含量、活性及其最终发挥的组织特异性功能各有不同。人胎盘中CYP19 基因主要采用 P1.1 进行转录[14],其次是PI.2 和 P2a;在卵巢中PⅡ(反刍动物称P2)是主要的启动子;睾丸和前列腺中也主要是PⅡ特异的转录物,故PⅡ常被称为生殖腺特异的启动子[15]。人和啮齿类黄体期主要的启动子是PII[14,16],而反刍动物主要为P1.1[17]。牛羊在妊娠过程中使用不同的启动子引导CYP19基因的表达,牛胎盘和黄体中主要是P1.1,绵羊胎盘中主要是P1.5;黄体中是P1.1和P1.5。在水牛卵泡发育成熟和黄体化过程中存在启动子的转换现象,主要表现为P2下调,P1.1上调。研究发现,该过程中CYP19基因组织特异性表达的变化和P2的组织特异性甲基化作用以及P1.1组蛋白修饰造成的染色质重组相一致[18]。推测,卵泡发育形成和黄体化过程中,启动子活性变化和P2的组织特异性甲基化作用以及P1.1组蛋白修饰造成的染色质重组有关。但有关启动子转换调节CYP19基因组织特异性表达的具体转录调节机制还不清楚。
J.Zhou等[19]研究表明,乳腺癌细胞能通过促进 C/EBPb 与 PⅡ调控区域相结合上调CYP19基因在脂肪成纤维细胞中表达,-517~278 bp为 PⅡ基本活性区域,在-317~304 bp为CCAAT增强子结合区域,与乳腺癌细胞共培养后,能使-517 bp 启动子活性上调 5.7 倍,而当通过定点突变CCAAT结合区域后,这一上调作用消失。M.Pannetier等[20]克隆了山羊卵巢特异性启动子中不同长度的5个片段,pAro1(1 104 bp)、pAro2(507 bp)、pAro3(180 bp)、pAro4(110 bp)和pAro5(40 bp),并构建了荧光素酶报告基因载体(pCYP19-Luc),进行了绵羊颗粒细胞转染试验。于今非等[21]克隆了水牛、黄牛CYP19启动子0.9和1.6 kb各两个片段,并构建了真核表达载体在细胞水平进行检测。结果显示黄牛、水牛的CYP19启动子(0.9 kb)可启动EGFP在卵巢颗粒细胞中特异表达,但效率非常低。而有关绵羊CYP19基因启动子P2完整序列及其在颗粒细胞中的特异转录活性研究未见报道。本研究旨在参考已知的CYP19基因启动子P2序列,克隆绵羊CYP19 基因卵巢启动子序列的不同区域片段,利用分子生物学软件进行功能分析预测,并分别构建真核表达载体,在细胞水平上对其活性进行检测和比较,为与繁殖相关的转基因动物研究提供基础。
1 材料与方法
1.1 试验材料
pEGFP-N2质粒为本实验室保存,pMD-19T(Simple)载体、限制性核酸内切酶购自TaKaRa公司;胶回收纯化试剂盒、质粒提取试剂盒购自上海生工,基因组提取试剂盒和大肠杆菌DH5a购自全式金公司。小尾寒羊卵巢和胎儿取自唐县屠宰场。
1.2 试验方法
1.2.1 卵巢采集及DNA提取 小尾寒羊屠宰后立即取出双侧卵巢组织,放入液氮中,带回实验室备用。卵巢组织DNA提取按组织DNA试剂盒操作步骤进行。
1.2.2 引物设计与合成 根据已公布的绵山羊基因组序列,利用Primer Premier 5.0软件设计引物,并在引物序列的正反两端,分别添加AseI和NheI酶切位点,用于绵羊CYP19基因卵巢启动子-683~-1 199 bp和-31~-1 130 bp扩增,预期片段大小为517和1 100 bp两个片段,分别记为CYP19 P 0.5和CYP19 P 1.1。引物由上海生物工程公司合成,具体引物序列见表1。
表1 PCR引物
Table 1 Primers of PCR
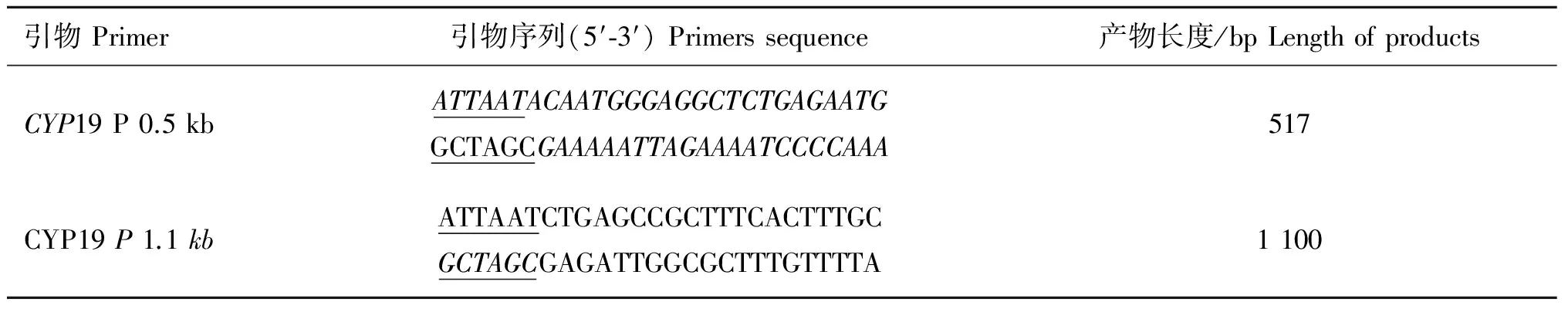
引物Primer引物序列(5′⁃3′)Primerssequence产物长度/bpLengthofproductsCYP19P0.5kbATTAATACAATGGGAGGCTCTGAGAATGGCTAGCGAAAAATTAGAAAATCCCCAAA517CYP19P1.1kbATTAATCTGAGCCGCTTTCACTTTGCGCTAGCGAGATTGGCGCTTTGTTTTA1100
1.2.3 绵羊CYP19基因卵巢启动子片段的扩增和克隆 PCR扩增反应体系为25 μL:其中引物1.0 μL;Mix 10 μL;DNA 1.0 μL;ddH2O 13 μL。CYP19 P 0.5 kb扩增程序:95 ℃ 2 min;95 ℃ 30 s,57 ℃ 30 s ,72 ℃ 1.5 min,30 循环;72 ℃5 min。CYP19 P 1.1扩增程序:95 ℃ 5 min;95 ℃ 30 s,63 ℃ 30 s,72 ℃1.5 min,35 循环;72 ℃10 min。
用1.5%琼脂糖凝胶进行电泳检测,凝胶成像系统下,割取目的条带,并用小量胶回收试剂盒回收,与PMD19TM(Simple)载体进行连接。转化E.coliDH5α感受态细胞。涂平板37 ℃过夜,挑取白色单菌落,在添加氨苄青霉素的液体LB培养基中培养8~12 h,进行菌液PCR。对PCR为阳性的菌液进行质粒小提,送北京华大生物公司进行测序。
1.2.4 克隆片段序列的生物信息学分析 利用BioEdit 7.0对克隆获得的片段序列的峰图进行比对,用ContingExpress进行序列拼接,利用 BLAST比较克隆序列与已知序列的同源性。采用TATA signal prediction(http://zeus2.itb.cnr.it/webgene/wwhc-tata.html)以及转录结合位点(http://mbs.cbrc.jp/research/db/TFSEARCH.html)、MatInspector(www.genomatix.de)、 TRANSFAC(http://www.gene-regulation.com/pub/programs.html)、CpG岛预测软件(http://www.uscnorris.com/cpgislands2/cpg.aspx和http://zeus2.itb.cnr.it/cgibin/wwwcpg.pl)软件对克隆序列潜在的调控元件及其CpG岛等进行分析,对其转录活性进行分析预测。
1.2.5 重组表达载体的构建 将测序正确的含CYP19 1.1 kb和CYP19 0.5 kb片段的质粒与pEGFP-N2质粒载体分别应用AseI和NheI进行双酶切,电泳获得的片段与去除CMV的pEGFP-N2载体片段应用DNA凝胶试剂盒回收。用T4DNA连接酶16 ℃过夜连接回收产物,之后转化DH-5α感受态大肠杆菌,在卡那霉素抗性平板上挑选阳性克隆,提取质粒,应用AseI和NheI进行酶切鉴定并测序。对酶切和测序结果均正确的样品,用去除内毒素的试剂盒进行质粒中提,测定浓度后备用。构建的载体分别命名为pCYP19-0.5-EGFP-N2和pCYP19-1.1-EGFP-N2。
1.2.6 细胞培养
1.2.6.1 胎儿成纤维细胞的原代培养和纯化:取怀孕50日龄小尾寒羊胎儿,参照文献[22]方法,进行细胞采集和培养。原代细胞采用DMEM/F12添加15%胎牛血清进行贴壁培养,细胞长成致密单层后,用0.25%胰酶消化,待大部分细胞变圆时,加培养液终止消化,并反复吹打,只收集脱壁细胞。成纤维细胞消化脱壁比上皮细胞快,待细胞刚变圆即终止消化,可得到纯化的成纤维细胞。细胞生长至70%~80%汇合状态时即可冷冻保存,以备基因转染使用。
1.2.6.2 卵巢颗粒细胞的分离培养:从屠宰场采集卵巢迅速放入37 ℃含双抗的生理盐水(100 IU·mL-1青霉素和0.1 mg·mL-1链霉素)的保温瓶中带回实验室。将卵巢过75%乙醇,用预热好的37 ℃生理盐水洗涤卵巢3次。用消毒过的眼科剪将卵巢附带的脂肪和结缔组织剔除干净,再用预热好的37 ℃生理盐水洗涤卵巢2次。用注射器连12号针头吸37 ℃预热的DMEM/F12 2 mL左右,吸取直径为3~6 mm卵泡中的卵泡液,捡出结构致密的卵丘-卵母细胞复合体(COCs),PBS 清洗干净后,用0.1%透明质酸酶作用,用移液枪轻轻吹打,挑出裸卵,将收集到的卵丘细胞1 000 r·min-1离心5 min,用适量含10%FBS的DMEM/F12(Hyclone)稀释,制成颗粒细胞悬浮液,调整细胞浓度为1×105mL-1,接种于25 mL培养瓶(Corning)中,在37 ℃ 5%CO2饱和湿度条件下进行培养,用于后续的细胞转染试验。
1.2.7 脂质体介导的重组质粒转染绵羊细胞研究 细胞转染使用 LipofectamineTMLTX + PLUS Reagent试剂盒进行。转染前将G1代的小尾寒羊颗粒细胞和G3代的小尾寒羊胎儿成纤维细胞分别接种到6孔板中,进行常规培养,待细胞长到 80%~90% 汇合度时,将重组质粒pCYP19-1.1-EGFP-N2和pCYP19-0.5-EGFP-N2在脂质体介导下,分别转染两种细胞。每组设3个重复。6孔板中每孔质粒DNA 使用量为2.5 μg,脂质体用量为12 μL,根据转染试剂说明进行转染。在125 μL细胞培养液中稀释2.5 μg核酸,加入3 μL PLUS,轻轻混匀后备用,在125 μL细胞培养液中稀释12 μL脂质体。将上述2种稀释液轻轻混合后在室温下孵育5 min形成250 μL复合体。将复合体加入6孔板中,轻轻摇动混合。在37 ℃ 5% CO2条件下培养6 h后,更换10% FBS的DMEMF12培养液。转染24 h后,开始利用倒置荧光显微镜观察EGFP表达情况,待转染72 h后,收集转染细胞样品,用流式细胞仪检测转染效率。
2 结 果
2.1 CYP19 卵巢启动子片段的扩增
以小尾寒羊卵巢DNA为模板,进行PCR扩增,用1.5%的琼脂糖电泳进行检测,扩增产物为约500和1 100 bp的片段(图1),条带单一、清晰明亮,可用于后续试验。

1、2.PCR产物;M.DL2000 DNA marker1,2 .PCR products;M.DL2000 DNA marker图1 PCR产物电泳图Fig.1 Electrophoresis of PCR products
2.2 CYP19 卵巢启动子片段克隆及测序
将电泳得到的两个片段分别进行切胶回收、连接、转化,克隆入PMD19TM(Simple),提取阳性质粒,送上海生工测序。利用ContingExpress、BioEdit和Chromas软件对测序结果进行分析和拼接,结果获得了1 100 bp(1.1 kb)和517 bp(0.5 kb)的序列。经比对发现克隆得到的两个序列与已有序列高度同源。其中1.1 kb序列片段与DQ122857序列的同源性达98.92%。517 bp片段与已公布的绵羊序列AJ012144.1完全一致,相似度达到100%,与已知山羊序列的同源性为99.34%。克隆小尾寒羊CYP19卵巢启动子片段(1.1 kb)与部分已知序列的同源性比对结果见图2。

图2 克隆小尾寒羊CYP19卵巢启动子片段与部分已知序列的同源性比对Fig.2 The homology of cloning CYP19 ovarian promoter fragment of small-tailed han sheep with some known sequence
2.3 扩增序列的转录因子结合位点分析
CpGIsland Searcher和CpG Island Prediction工具预测,发现克隆得到的两个片段区域无符合条件的CpG岛。进一步应用TFSEARCH(http://mbs.cbrc.jp/research/db/TFSEARCH.html)、TRANSFAC(http://www.gene-regulation.com/pub/programs.html)和MatInspector(www.genomatix.de) 软件对1 100和517 bp片段序列进行分析表明,具有类似于TATA-box、Oct-1等启动子的特征元件和多个C/EBPbeta、NF5、CRE、FOXL2、GATA-1、SRY、 AP1、MZF1等调控元件结合位点。部分转录因子结合位点见图3。

图3 克隆1.1 kb序列及部分潜在转录调控元件Fig.3 Nucleotide sequence of the 1.1 kb and partial transcriptional regulation sequences
2.4 重组载体的构建及鉴定
将酶切后的目的片段与去除CMV的pEGFP-N2线性化片段进行连接构建表达载体。过夜连接后转化培养,随机挑取10个克隆进行PCR检测,对阳性克隆菌液进行扩大培养,提取质粒后,用AseI和NheI进行酶切和测序,电泳酶切产物结果产生1.1 kb与4.1 kb和517 bp片段(图4),测序结果正确,表明CYP19 P0.5、CYP19 P1.1已成功克隆入去掉CMV的pEGFP-N2质粒,pCYP19-0.5-EGFP-N2和 pCYP19-1.1-EGFP-N2载体构建成功,重组质粒图谱见图5。

1、2.双酶切产物;M.DL5000 DNA marker;3.质粒对照1,2.Double enzyme products;M.DL5000 DNA marker;3.Plasmid control图4 pCYP19-0.5-EGFP-N2和pCYP19-1.1-EGFP-N2的双酶切鉴定Fig.4 Identification of pCYP19-0.5-EGFP-N2 and pCYP19-1.1-EGFP-N2
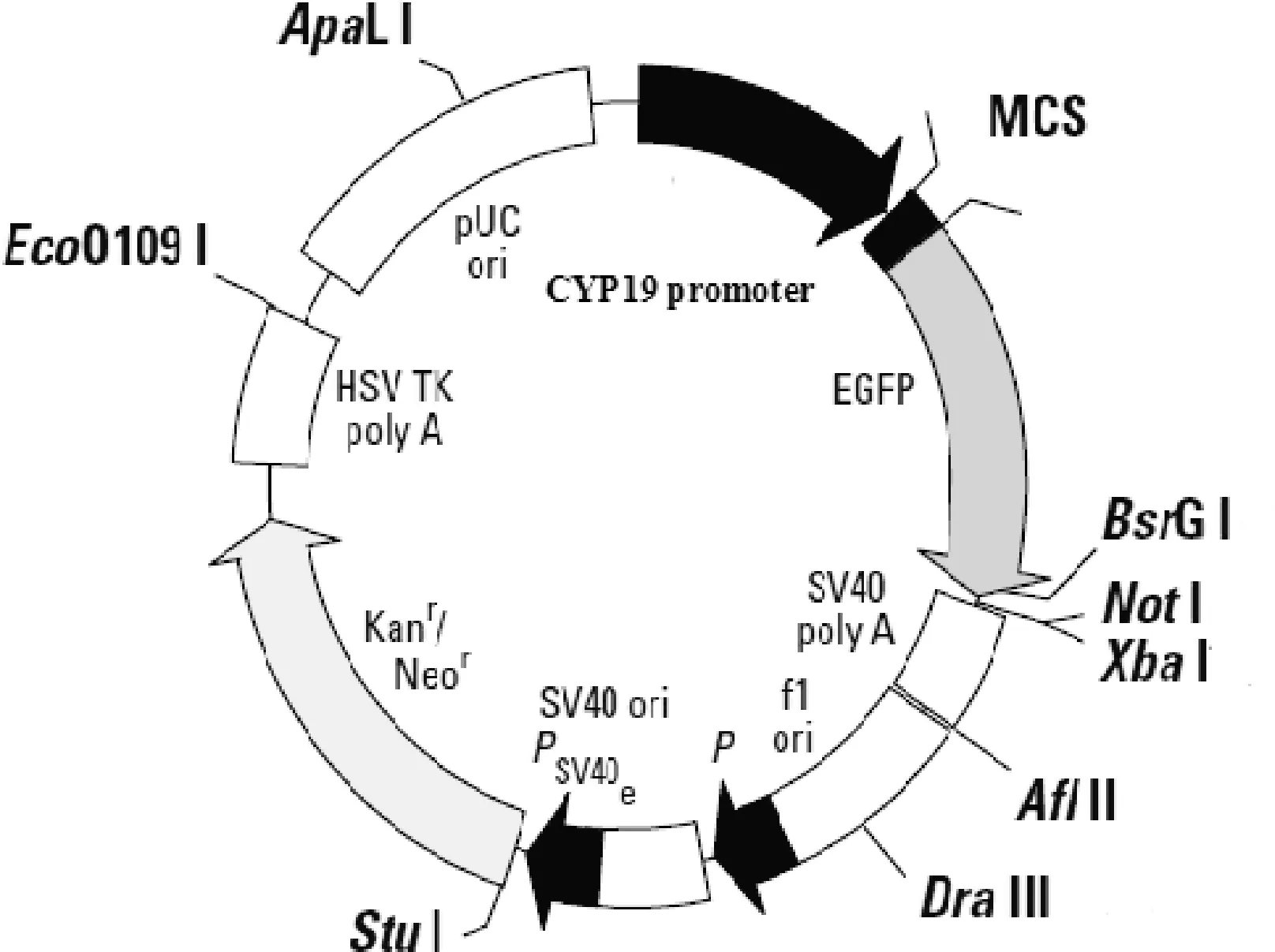
图5 重组质粒图谱Fig.5 Vector map of recombinant plasmid
2.5 重组质粒在绵羊细胞中的表达
pCYP19-0.5-EGFP-N2和pCYP19-1.1-EGFP-N2分别经脂质体LipofectamineTMLTX+PLUS包埋后,瞬时转染绵羊卵巢颗粒细胞和胎儿成纤维细胞。转染24 h后在倒置荧光显微镜下观察,可见细胞内的绿色荧光,表明质粒转染成功,报告基因开始表达,48 h后细胞荧光亮度有所增加,随着转染时间延长,阳性细胞数增多,72 h后最多。在绵羊胎儿成纤维细胞中也观察到了类似有少量绿色荧光表达(图6和7),表明克隆CYP19 1.1 kb(-31~-1 130 bp)序列具有生物学活性。而pCYP19-0.5-EGFP-N2经LipofectamineTMLTX+PLUS介导转染小尾寒羊颗粒细胞和胎儿成纤维细胞后,均未观察到荧光,表明克隆0.5 kb(-683~-1 199 bp)无转录活性,启动子关键活性部位未包含在该区域中。
收集转染72 h后的颗粒细胞,以空白作对照,用流式细胞仪测定转染效率。pCYP19-1.1-EGFP-N2转颗粒细胞的平均转染效率为41.66%,转小尾寒羊胎儿成纤维细胞的平均转染效率为20.16%。流式细胞仪测定结果见图8。
3 讨 论
本试验成功克隆了绵羊CYP19基因卵巢启动子的1.1和0.5 kb两个片段,对所得序列进行生物信息学分析发现了类似于TATA-box的启动子的特征元件和多个C/EBP、SRY、MZF-1、AP-1、GATA-1等若干对表达调控具有重要作用的序列元件。进一步将该序列与pEGFP-N2基因载体融合,替换掉该载体中的CMV启动子序列,成功构建了pCYP19-0.5-EGFP-N2和 pCYP19-1.1-EGFP-N2。利用脂质体LipofectamineTMLTX+PLUS介导,分别转染绵羊卵巢颗粒细胞和胎儿成纤维细胞。发现经pCYP19-1.1-EGFP-N2转染后在卵巢颗粒细胞和成纤维细胞中均有绿色荧光表达,而pCYP19-0.5-EGFP-N2在绵羊卵巢颗粒细胞和胎儿成纤维细胞中均未检测到荧光,意味着pCYP19- 1.1-EGFP-N2在颗粒细胞和成纤维细胞中均具有表达活性,能引导外源基因在其中的特异表达;而pCYP19-0.5-EGFP-N2不具有表达活性,调控绵羊CYP19基因卵巢启动子表达的核心序列区域不含在此0.5 kb(-683~-1 199 bp)序列。
研究发现,在卵巢特异性启动子的近端含有许多保守调控元件结合位点,如调控cAMP 信号通路的类cAMP反应元件序列((cAMP response element)-like sequence;CLS)[23-24]、孤核转录因子NR5A1/NR5A2和叉头框家族FOXL2[25]、AP3(Activation protein 3)和GATA结合位点等。研究证实,在卵巢不同种类的细胞中这些顺式作用元件起着不同的作用,对克隆1.1 kb序列的转录因子结合位点分析预测表明,1.1 kb也包含有CRE、NR5A1/NR5A2和叉头框家族FOXL2等调控元件结合位点,预示克隆片段具有卵巢特异性启动子活性。此外, FOXL2在不同物种间的作用不同,在人和小鼠中,表现为抑制PII活性[26-27],在山羊中则表现为促进P2活性[20]。尽管本试验克隆的1.1 kb和山羊P2以及牛PII高度同源,但仍存在一些单碱基变异,这些变异可能会导致一些未知潜在调控元件结合位点的变化,进而影响其活性和卵巢表达的特异性。C.H.Tu等[28]报道,能够在卵巢组织中特异表达的启动子仅有OSP-1、OSP-2。CYP19、抑制素a(Inhibin a)和苗勒管抑制物质Il型受体(Mulledan inhibiting substance typeII receptor,MISIIR)等针对卵巢的特异性启动子除了能调控基因在卵巢组织高效表达外,在其它组织也有相当水平的启动功能,缺乏高度专一性。pCYP19-1.1-EGFP-N2在颗粒细胞和成纤维细胞中转染效率的不同,显示克隆所得1.1 kb卵巢启动子在组织特异性上缺乏高度专一性。有研究发现,人CYP19基因外显子2的P2 启动子可以启动标记基因在性腺组织的特异表达[29]。本试验克隆得到的绵羊CYP19基因卵巢启动子的1.1 kb虽具有活性,但并非真正卵巢特异性表达。M.Margaret等[30]克隆了PIIa 43、278和2 700 bp不同长度的启动子片段,并连接人生长激素基因作为表达基因,制备了转基因小鼠,结果显示,278和 2 700 bp两种片段的小鼠在脑、肾、肺、肝、皮肤、大脑、卵巢等脏器中均有表达,P II a43只在卵巢特异表达,并在雌性小鼠中,该启动子将目的基因定位在颗粒细胞、黄体以及黄体化间质细胞上。而将转基因小鼠组织表达分析得知,278 bp片段表达程度较2 700 bp的片段高,而43 bp片段没有表达,说明调控外显子PIIa卵巢特异表达的区域在43~278 bp。由此推断,调控CYP19基因卵巢特异表达的核心、关键区域可能集中在1.1 kb序列的某一区域。

A.转染24 h; B.转染48 h;C.转染72 h。下同A.Transfection for 24 h;B.Transfection for 48 h;C.Transfection for 72 h.The same as Fig.7图6 pCYP19-0.5-EGFP-N2转染绵羊颗粒细胞100×Fig.6 The transfection of pCYP19-0.5-EGFP-N2 in sheep granulosa cells 100×

图7 pCYP19-1.1-EGFP-N2转染绵羊成纤维细胞100×Fig.7 The transfection of pCYP19-1.1-EGFP-N2 in sheep fetal fibroblast cells 100×
A.颗粒细胞;B.成纤维细胞A.Granulose cells;B.Fetal fibroblast cells图8 重组质粒转染颗粒细胞和成纤维细胞72 h流式图Fig.8 The transfection of CYP191.1 in granulose cells and fetal fibroblast cells for 72 h assayed by flow cytometry
本研究仅克隆了绵羊CYP19基因卵巢启动子的1.1和0.5 kb两个片段,对其调控元件的分析仅限于推测,其活性研究仅限于绵羊颗粒细胞和成纤维细胞的体外尝试。有关CYP19基因启动子1.1 kb片段的活性功能验证还有待于在更多种类细胞进行试验。要确定绵羊CYP19基因卵巢启动子的核心区域,尤其是在卵巢中特异表达的区域,还需进一步研究证实。
[1] GUIGUEN Y,FOSTIER A,PIFERRER F,et al.Ovarian aromatase and estrogens:A pivotal role for gonadal sex differentiation and sex change in fish[J].GenCompEndocrinol,2010,165(3):352-366.
[2] KELLER J M,MCCLELLAN-GREEN P.Effects of organochlorine compounds on cytochrome P450 aromatase activity in an immortal sea turtle cell line[J].MarEnvironRes,2004(58):347-351.
[3] GYLLENHAMMAR I,ERIKSSON H,SODERQVIST A,et al.Clotrimazole exposure modulates aromatase activity ingonads brain during gonadal differentiation in Xenopus tropicalis frogs[J].AquaticToxicol,2009,91(2):102-109.
[4] CHEN S A,BESMAN M J,SPARKES R S,et a1.Human aromatase:cDNA cloning,Southern blot analysis,and assignment to the gene to chromosomel 5[J].DNA,1988,7(1):27-38.
[5] MAHENDROO M S,MENDELSON C R,SIMPSON E R.Tissue-specific and hormonally-controlled alternative promoters regulate aromatase cytochrome P450 gene expression in human adipose tissue[J].BiolChem,1993(268):19463-19470.
[6] SEBASTIAN S,TAKAYAMA K,SHOZU M,et al.Cloning and characterization of a novel endothelial promoter of the human CYP19(Aromatase P450) gene that is up-regulated in breast cancer tissue[J].MolEndocrinol, 2002,16(10):2243-2254.
[7] BULUN S E,SEBASTIAN S,TAKAYAMA K,et al.The human CYP19(aromatase P450) gene:update on physiologic roles and genomic organization of promoters[J].SteroidBiochemMolBiol,2003,86(3-5):219-224.
[8] HINSHELWOOD M M,LIU Z,CONLEY A J,et al.Demonstration of tissue-specific promoters in nonprimate species that express aromatase P450 in placentae[J].BiolReprod,1995,53(5):1151-1159.
[9] FüRBASS R,KALBE C,VANSELOW J.Tissue-specific expression of the bovine aromatase encoding gene uses multiple transcriptional start sites and alternative first exons[J].Endocrinology,1997,138(7):2813-2819.
[10] VANSELOW J,ZSOLNAI A,FESüS L,et al.Placenta-specific transcripts of the aromatase encoding gene include different untranslated first exons in sheep and cattle[J].EurJBiochem,1999(265):318-324.
[11] 王金玲,王 永,郑玉才,等.草地藏系绵羊CYP19基因第9内含子和启动子2多态性分析[J].绵阳师范学院学报,2009,28(5):64-66. WANG J L,WANG Y,ZHENG Y C,et al.Polymorphism of the intron 9 and promoter 2 ofCYP19 gene in Tibetan sheep[J].JournalofMianyangNormalUniversity(NaturalScienceEdition),2009,28(5):64-66.(in Chinese)
[12] SIMPSON E R,ZHAO Y,AGARWAL V L,et al.Aromatase expression in health and disease[J].RecProgHormRes,1997(52):185-214.
[13] SIMPSON E R,CLYNE C,RUBIN G,et al.Aromatase——a brief overview[J].AnnuRevPhysiol,2002(64):93-127.
[14] MEANS G D,KILGORE M W,MAHENDROO M S,et al.Tissue-specific promoters regulate aromatase cytochrome P450 gene expression in human ovary and fetal tissues[J].MolEndocrinol,1991(5):2005-2013.
[15] KATSUMI T,SATOSHI N,YUTAKAS S.Identification and characterization of transcriptional regulatory elements of the human aromatase cytochrome P450 gene[J].SteroidBiochemMolBiol, 1996,56(1):151-159.
[16] STOCCO C.Invivoandinvitroinhibition of cyp19 gene expression by prostagland in F2a in murine luteal cells:implication of GATA-4[J].Endocrinology,2004,145(11):4957-4966.
[17] LENZ S,P HLAND R,BECKER F,et al.Expression of the bovine aromatase cytochrome P450 gene(CYP19) is primarily regulated by promoter 2 in bovine follicles and by promoter 1.1 in corpora lutea[J].MolReprod,2004,67(4):406-413.
[18] MONGA R,GHAI S,DATTA T K,et al.Tissue-specific promoter methylation and histone modification regulateCYP19 gene expression during folliculogenesis and luteinization in buffalo ovary[J].GenCompEndocrinol,2011,173(1):205-215.
[19] ZHOU J,GURATES B,YANG S,et al.Malignant breast epithelial cells stimulate aromatase expression via promoterⅡin human adipose fibroblasts:an epithelial-stromal interaction in breast tumors mediated by CCAAT/Enhancer binding proteinbeta[J].CancerRes, 2001,61(5):2328-2334.
[20] PANNETIER M,FABRE S,BATISTA F,et al.FOXL2 activates P450 aromatase gene transcription:towards a better characterization of the early steps of mammalian ovarian development[J].MolEndocrinol, 2006,36(3):399-413.
[21] 于今非.CYP19和OSP-1卵巢特异表达启动子克隆与检测[D].南宁:广西大学,2010. YU J F.Cloning and expression assay of ovarian specific expressing promoter:OSP-1 and CYP19[D].Nanning:Guangxi University,2010.(in Chinese)
[22] 斯佩克特D L,戈德曼R D,苯因万德L A.细胞实验指南[M].黄培堂,译.北京:科学出版社,2003. SPECTOR D L,GOLDMAN R D,LEINWAND L A.Cells:A laboratory manual[M].HUANG P T,Translation.Beijing:Science Press,2003.(in Chinese)
[23] FITZPATRICK S L,RICHARDS J S.Regulation of cytochrome P450 aromatase messenger ribonucleic acid and activity by steroids and gonadotropins in rat granulosa cells[J].Endocrinology,1991,129(3):1452-1462.
[24] MICHAEL M D,MICHAEL L F,SIMPSON E R.A CRE-like sequence that binds CREB and contributes to cAMP-dependent regulation of the proximal promoter of the human aromatase P450(CYP19) gene[J].MolCellEndocrinol,1997,134(2):147-156.
[25] STOCCO C.Aromatase expression in the ovary:hormonal and molecular regulation[J].Steroids,2008,73(5):473-487.
[26] KUO F T,BENTSI-BARNES I K,BARLOW G M,et al.Mutant Forkhead L2(FOXL2) proteins associated with premature ovarian failure(POF) dimerize with wild-type FOXL2,leading to altered regulation of genes associated with granulosa cell differentiation[J].Endocrinology,2011,152(10):3917-3929.
[27] KUO FT,FAN K,BENTSI-BARNES I,et al.Mouse forkhead L2 maintains repression of FSH-dependent genes in the granulosa cell[J].Reproduction,2012,144(14):485-494.
[28] TU C H,LIU W P,HUANG L L.Cloning and transcriptional activity of a novel ovarian-specific promoter from rat retrovirus-like elements[J].ArchBiochemBiophys,2009,485(1):24-29.
[29] MEANS G D,MAHENDROO M.Structural analysis of the gene encoding human aromatase cytochrome P450,the enzyme responsible for estrogen biosynthesis[J].BiolChem,1989,264(32):19385-19391.
[30] HINSHELWOOD M M,MENDELSON C R.Tissue-specific expression of the human CYP19(aromatase) gene in ovary and adipose tissue of transgenic mice[J].JSteroidBiochemMolBiol,2001(79):193-201.
(编辑 程金华)
Cloning and Expression Assay of SheepCYPl9 Gene Ovarian Promoter
SUN Hong-xin1,2,ZHANG Ying-jie1*,LIU Yue-qin1,CHEN Xiao-yong2,DUN Wei-tao2
(1.CollegeofAnimalScienceandTechnology,AgriculturalUniversityofHebei,Baoding071000,China; 2.HebeiInstituteofAnimalScienceandVeterinaryMedicine,Baoding071000,China)
The research was conducted to clone the sheepCYP19 gene ovarian promoter fragments and detect the tissue-specific expression in cells level.According to the known sequence the specific prmiers were designed,1.1 and 0.5 kb of sheepCYP19 gene ovarian promoter fragment were amplified by PCR,then the sequences were analyzed by software with published sequences.After eukaryotic expression vector pCYP19-1.1-EGFP-N2and pCYP19-0.5-EGFP-N2were constructed by cloning promoter fragments into the pEGFP-N2vector without CMV,then the recombinant plasmids were transfected into sheep granular cells and fetal fibroblast cells by liposome LipofectamineTMLTX+PLUS.The EGFP fluorescence expression was observed under the microscope after transfection for 24,48 and 72 h.The sequenced results showed that sheepCYP19 gene promoter fragments which were cloned were 1.1 and 0.5 kb length and had highly homologous with the published sequences.The sequence analyzing with transcription factor binding sites prediction software indicated that the promoter fragment contained a core promoter cis-element smilar with TATA box and transcription factor binding sites.24 h after transferring with pCYP19-1.1-EGFP-N2,the EGFP-expressing positive granulosa cells could be observed,48 h after transferring,the EGFP-expressing positive granulosa was increased,72 h after transfection the EGFP-expressing positive granulosa increased to the most.But after transfection the EGFP which was expressed in sheep fetal fibroblast cells was little.The results also showed that no EGFP-expressing positive granulosa cells and fetal fibroblast cells were observed at 24,48 and 72 h after transfection with pCYP19-0.5-EGFP-N2.The sheepCYP19 promoter 1.1 kb can drive foreign gene expressing in sheep granulosa cells and it can be used for the functional studies of fecundity-related genes and transgenic animal research,but it is not ovarian specific expression promoter.
sheep;CYP19;ovarian promoter;expression
10.11843/j.issn.0366-6964.2015.09.008
2015-01-26
国家肉羊产业技术体系资助项目(CARS-39)
孙洪新(1978-),女,山东临清人,高级畜牧师,博士生,主要从事羊遗传繁育研究, E-mail:sdlqshx@126.com
*通信作者:张英杰,教授,博导,主要从事羊遗传育种及营养研究,E-mail :zhangyingjie66@126.com
S826.2
A
0366-6964(2015)09-1540-09
