和睦湿地浮游植物群落结构特征及环境因子分析
2015-03-20林施泉茆传奇邵晓阳
姜 丹,林施泉,刘 忱,茆传奇,邵晓阳
(1.杭州师范大学生命与环境科学学院,生态系统保护与恢复杭州市重点实验室,浙江 杭州310036;2.国家海洋局第二海洋研究所,国家海洋局海洋生态系统与生物地球化学重点实验室,浙江 杭州310012)
浮游植物是水生态系统中的重要初级生产者,地球上一半左右的初级生产力都归功于浮游植物[1-2];其群落组成对全球生物地球化学循环具有重要的作用,继而反馈性影响全球环境变化[3-4].近年来,浮游植物已成为生物监测以及评价水质和水体营养状况的重要生物指标,在国内外已被广泛采用[5-6].我国许多学者利用浮游植物来监测和评价水生态环境.邓建明等对太湖流域主要河道浮游植物的群落结构进行了对比分析,发现太湖流域浮游植物物种多样性存在季节差异,且与环境因子显著相关[7].林施泉等利用浮游植物群落结构和生物多样性指数对福建木兰溪流域进行了水质评价,发现流域内水体呈中营养化污染并向富营养化污染转变[8].陆欣鑫等研究了浮游植物功能分组与环境因子的关系,发现电导率、总磷和水温是驱动呼兰河湿地浮游植物功能组演替的主要环境因素[9].
和睦湿地,紧邻杭州西溪湿地,占地总面积约10 km2,是杭州城西湿地的重要组成部分.和睦湿地地处余杭、五常、闲林、仓前等四大片区的中心地带,水网、河道、水塘等星罗密布,平均水深约2.78 m,植被繁多,以枫杨、垂柳、早竹、芦苇、凤眼莲、喜旱莲子草等为主,是浙江省内比较罕见的尚未经历现代开发的所谓“原生态”湿地[10].然而,由于近年来城市化进程的加快,特别是湿地内部人类活动的频繁介入,在相当程度上对湿地生态环境造成了严重破坏.本文通过对和睦湿地浮游植物的监测和调查,对浮游植物多样性与环境因子进行分析,探讨和睦湿地浮游植物群落季节变化的特点以及环境驱动因素,以期为和睦湿地水环境的恢复与保护提供科学依据和生物学资料.
1 材料与方法
1.1 采样点设置
根据和睦湿地生态特点,在和睦水乡湿地内共设置15个采样点(图1).其中,1为出水口,14、15号点分别为东西入水口,6~11、13为和睦水乡湿地的核心区.分别于2011年7月、10月和2012年1月、4月中旬进行浮游植物和水环境样品的采集,共计4次.
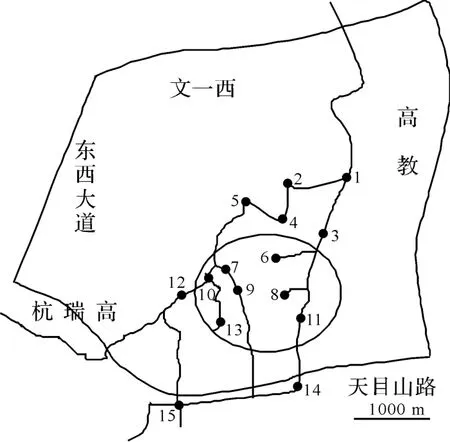
图1 和睦水乡湿地采样点分布Fig.1 Distribution of sampling sites in Hemu wetland
1.2 实验方法
1.2.1 样品采集、鉴定与指标测定
现场采用YSI6600测定表层0.5 m 水深处水温、浊度、溶解氧(DO)、p H 值和叶绿素a(Chla)等指标.将自水面0.5 m 深处采集的水样酸化后带回实验室,进行总氮(TN)、总磷(TP)、5日生化需氧量(BOD5)、化学需氧量(COD)等理化指标的测定;样品的采集、保存与测定详细步骤参考《水和废水监测分析方法》.
浮游植物的采样和观察方法按常规浮游生物调查方法[11]进行,定性标本采样用25号浮游生物网采集,用4%甲醛溶液现场固定;定量标本采集用1 L采水器于水下0.5 m 处采样,将水样置于1.5 L采水瓶中,加入鲁戈氏液现场固定,经48 h沉淀浓缩至500 m L,再静置24 h后,抽取上清液,定容至60 m L.藻类计数采用迅数S300 型Algacount藻类计数仪进行.每个样品重复计数3片,误差超过15%则进行第4片计数后取其中结果相近的3片平均值.物种鉴定主要依据《中国淡水藻类——系统、分类及生态》[12]、《藻类名词及名称》[13]、《淡水微型生物图谱》[14]和《浙江省主要常见淡水藻类图集》[15].
1.2.2 生物多样性分析方法
本文运用Origin 8.6 处理数据和作图,主成分分析和聚类分析主要采用大型多元分析软件PRIMER V6.0,多样性指数的计算使用其子模块Diversity,包括Margalef丰富度指数(d)、Shannon-Wiener多样性指数(H′)、Pielou 均匀度指数(J′)和Simpson生态优势度指数(D),计算公式分别为:
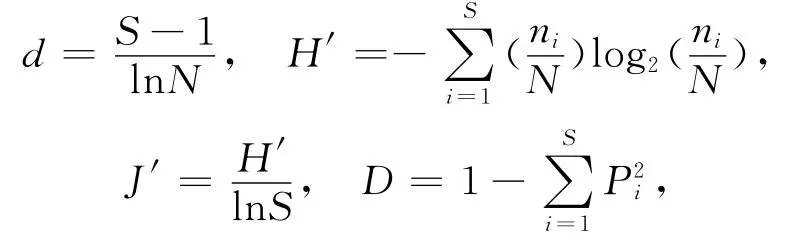
式中:S为生物的种类数;N为群落的个体总数;ni为i种的个体数;Pi为第i属的个体数(ni)占总个体数(N)的比值.
优势度的计算方法按照Mcnaughton 优势度指数公式[16]:
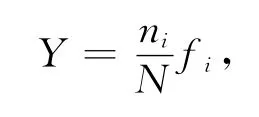
式中:ni为第i种的总个体数;N为所有物种的总个体数;fi为第i种在各站位出现的频率;Y>0.02定为优势种.
2 结果
2.1 和睦湿地水环境因子变化
由表1可知:和睦湿地四季水温有明显的变化,变幅为8.02~29.05℃,全年平均为20.17℃;浊度为8.28~10.01,平均为8.92;DO 为5.36~9.49 mg/L,平均为6.58 mg/L;水体p H 值呈弱碱性,为7.62~8.17,平均为7.82;水体TN 为2.76~6.75 mg/L,平均为5.12 mg/L,夏季和冬季高于春季和秋季;TP为0.18~5.62 mg/L,平均为2.34 mg/L,春季和冬季高于夏、秋季;COD 为21.64~63.32 mg/L,平均为44.36 mg/L;BOD5为4.47~13.03 mg/L,平均为8.40 mg/L;水体Chla含量为3.72~27.78μg/L,平均为9.77μg/L,春季Chla含量最高.
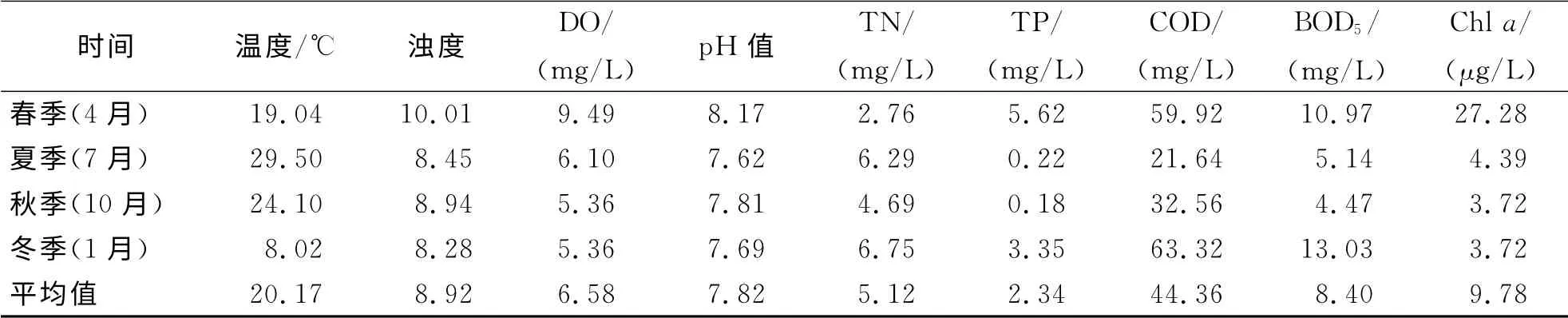
表1 和睦湿地水环境因子Tab.1 The environmental variables in Hemu wetland
2.2 和睦湿地浮游植物种类组成及优势种
通过对和睦湿地4次采样的浮游植物标本进行观察,共鉴定浮游植物101种,隶属于8门20目36科61属.其中绿藻门Chlorophyta种类居首位,有26属35种,占34.65%;其次为硅藻门Bacillariophyta 13属23种,占22.77%;裸藻门Euglenophyta 6 属22 种,占21.78%;蓝藻门Cyanophyta 9 属13 种,占12.87%;除此之外,隐藻门Cryptophyta有2属3种;金藻门Chrysophyta 2属2种;甲藻门Pyrrophyta 2属2种;黄藻门Xanthophyta 1属1种.本次调查区域内浮游植物种类数量表现出季节性差异,春季和夏季浮游植物种类数较高,分别达到68种和75种,秋季和冬季种类数相同,均为57种.浮游植物优势种主要有四尾栅藻Scenedesmusquadricauda、梅尼小环藻Cyclotellameneghiniana、卵形隐藻Cryptomonsovata、啮蚀隐藻Cryptomonserosa、绿色裸藻Euglenaviridis和小型色球藻Chroococcusminor等.
2.3 和睦湿地浮游植物丰度分布
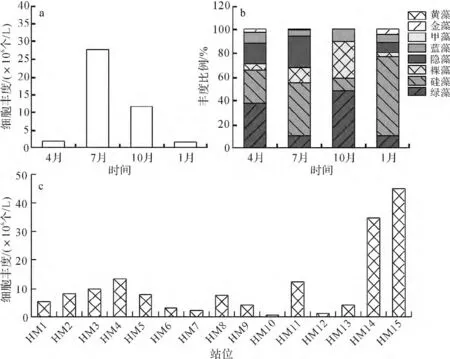
图2 和睦湿地浮游植物细胞丰度Fig.2 Phytoplankton abundance in Hemu wetland
和睦湿地浮游植物细胞丰度在调查期间季节变化明显,细胞丰度高峰区在夏季和秋季(图2a).浮游植物细胞丰度在调查周年内变化范围为17.47×106~41.55×106个/L,全年细胞丰度平均值为27.99×106个/L,其中:硅藻占总丰度的比例最高,达到37.46%;其次为绿藻,占26.50%;裸藻和隐藻占总丰度的比例相近,为13%左右.不同季节浮游植物丰度分布不同,春季绿藻和硅藻所占比例较高,夏季以硅藻和隐藻占优势,秋季以绿藻和裸藻为主,冬季硅藻占显著优势(图2b).从站位来看(图2c),最高值出现在HM15站位,细胞丰度平均为44.98×106个/L,其次是HM14站位,细胞平均丰度为34.86×106个/L,最低值出现在HM10站位,平均值为0.77×106个/L.
2.4 和睦湿地浮游植物群落多样性
群落物种多样性是群落组织独特的生物学特征,它反映了群落特有的物种组成和个体丰度特征.由图3显示,不同季节浮游植物各多样性指数略有不同,其中Margalef指数在0.60~1.68 之间,平均值为1.25,夏季和秋季高于冬季和春季.Pielou指数在0.59~0.81之间,平均值为0.64,春季和秋季高于夏季和冬季.Shannon-Weaver指数在在1.21~2.51之间,平均值为2.00;Simpson指数在0.62~0.86之间,平均值为0.74;这两个指数均表现出春季和夏季高于秋、冬季.
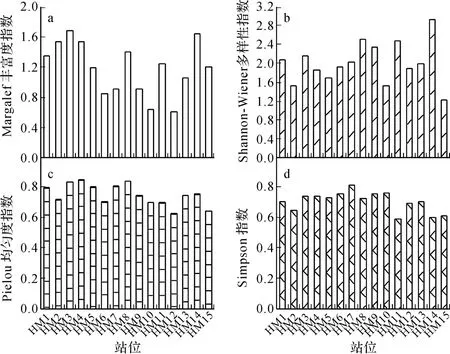
图3 和睦湿地浮游植物群落多样性变化Fig.3 Phytoplankton diversity in Hemu wetland
2.5 浮游植物与环境因子的关系
水体环境因子与浮游植物丰度之间的相关分析见表2.溶解氧与其他环境因子存在极显著正相关,而温度与浮游植物呈现极显著正相关外,与其他大部分环境因子为负相关.除此之外,水体TN 浓度升高,可使浮游植物丰度显著增大,而TP可使水体Chla浓度升高.结果还显示,浮游植物丰度与Chla浓度不存在明显相关性.原始数据经Kaiser法标准化后,采用最大方差正交旋转法进行因子旋转,提取得到4个主因子.由表3得知:第一主因子的贡献率为39.02%,强正相关的因子变量为TN、BOD 和COD;第二主因子的贡献率为17.99%,正相关因子变量为Chla和p H;第三主因子与温度呈现明显的负相关关系,第四主因子与浊度呈显著正相关.
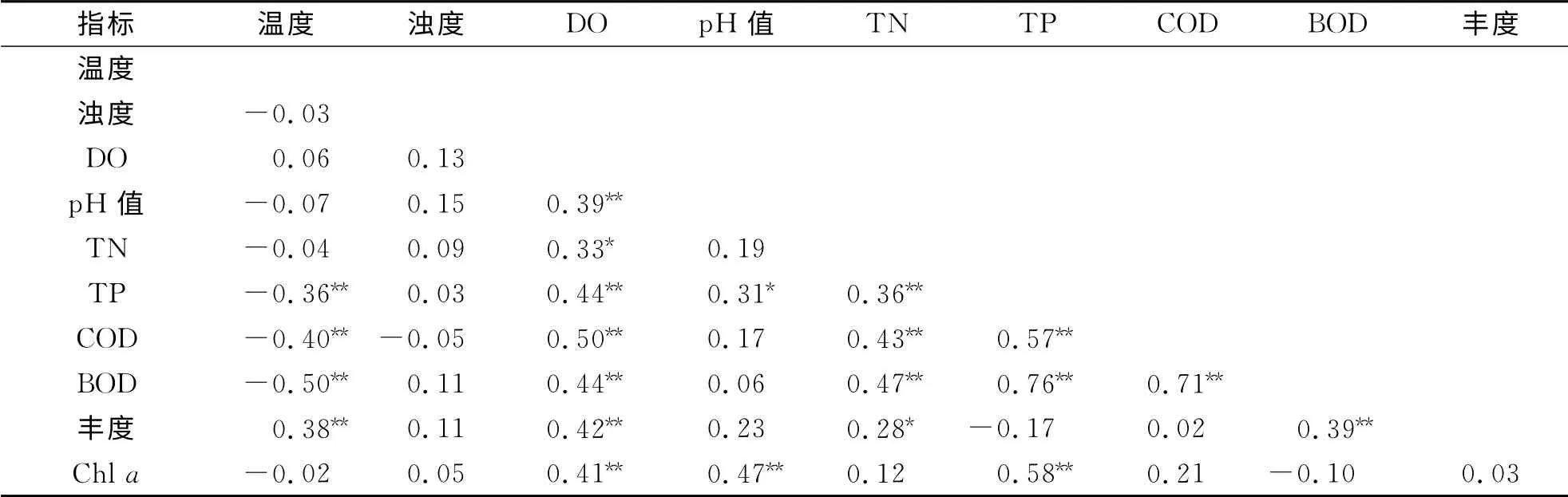
表2 和睦湿地浮游植物丰度与水环境因子之间的相关关系Tab.2 The relationship between the abundance of phytoplankton and the environmental variables in Hemu wetland
3 讨论
3.1 浮游植物群落季节变化与环境因子的关系
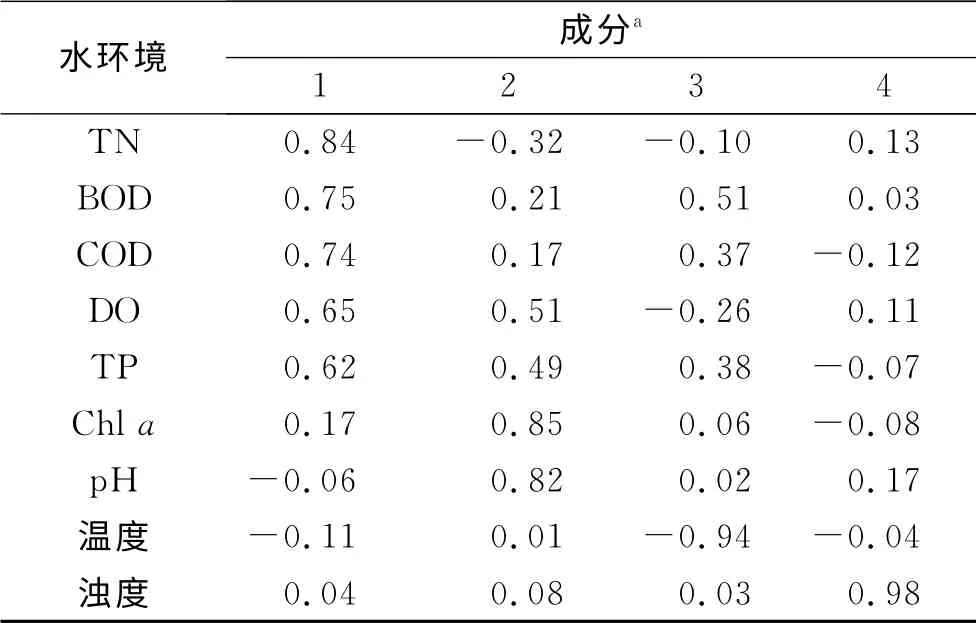
表3 和睦湿地水环境因子最大方差旋转因子矩阵Tab.3 Varimax rotated factor matrix for the environmental data in Hemu wetland
浮游植物的群落结构是水质生物学评价的重要参数[1,17].由于存在地域和水体类型差异,不同河流生态系统浮游植物群落的组成和动态存在较大的差异[18-19].和睦湿地调查显示,硅藻、绿藻和裸藻在种类和数量上都占优势,这个结果与西溪湿地封闭水塘的结果大致相同.贾兴焕等在2009年对西溪湿地31个封闭池塘水体浮游植物进行研究,发现物种数上绿藻最多,蓝藻和隐藻的物种数虽少,但数量上却占优势[3].和省内其他水库相比较,物种种类数和汤浦水库浮游植物(105种)接近[20],略低于紧水滩水库(139种)[21],明显低于横山水库(246种)[22].
水体环境的变化,比如水温、营养盐以及水体扰动程度,都会影响浮游植物群落的演替[23].当周围环境发生变化,藻类能通过自身的群落演替进而维持生态平衡[24-25].水温通常是影响浮游植物发育和繁殖的限制因子,直接影响浮游植物的群落组成、演替方向以及部分生理特性[26].在本次调查结果中,水体温度与和睦湿地浮游植物丰度存在显著正相关(R2=0.380,P=0.003),而且随着水温的升高,Margalef等多样性指数明显提高.Ma等[27]在扎龙湿地的研究显示:水温能够显著影响浮游植物的群落结构,进而影响浮游植物的群落演替.我们的结果显示:和睦湿地浮游植物的群落结构也呈现明显的季节现象,夏、秋两季水温较高,使湿地内部耐受高温的藻类存活下来,在丰度上蓝藻和裸藻占优势;冬季温度较低,主要以硅藻存在;春季水温比较适宜,绿藻在这种环境下就显得较为活跃[28].这与Sommer等[29]提出的PEG(plankton ecology group)模型大致相同,其通过对大量温带湖泊浮游生物和理化因子数据的分析,认为浮游植物群落季节演替大概是:从冬春的隐藻和硅藻转变为夏季的绿藻,到夏末秋初则是蓝藻占优势,秋季时硅藻数量再次上升,这一模式主要反映中营养水平深水湖泊的情况.值得一提的是,和睦湿地在地理位置上属于亚热带,受亚热带季风气候影响,可能造成与PEG模型略有差别,但仍可以看出,水温对浮游植物群落结构季节变化起着至关重要的作用.
3.2 和睦湿地水环境评价
本次结果显示,在和睦湿地出现的优势种大部分为污染指示种,代表性藻种如四尾栅藻为β-ms(超富营养型)代表性种类,啮蚀隐藻为α-ms(富营养型)指示种,还有囊裸藻、鱼腥藻、小球藻、纤维藻和舟形藻等,也大都属于污染指示种.与其他湿地相比较[26,28],和睦湿地存在浮游植物多样性指数较低,且冬季低于其他季节的现象,说明冬季的水质最差,属于α-污染水体[20,22].同时,湿地水环境中的TN、TP、COD 和Chla等也均超出了湖泊富营养化临界值,特别是TN 和TP远远超过《地表水环境质量标准(GB3838-2002)》V 类水标准,其水质状况低于相邻的西溪湿地[30-31].综合来看,可以推测和睦湿地在调查期间内基本处于富营养化的状态.究其原因,主要有两点:一是和睦湿地大部分封闭水塘用以传统的鱼类养殖,投放过量的饲料和渔药,大大超过了环境容量和环境自净能力,导致营养不均衡,特别是N、P 等营养物质过剩;二是和睦湿地东西入水口(HM14和HM15站位),其地理位置距离闲林工业区较近,且周边有多个房地产开发在建项目,工业污水的大量排放对和睦湿地水环境造成严重的影响.因此,我们认为应该加强和睦湿地富营养化防治的工作,防止其水环境状况进一步恶化,这对杭州城西湿地的保护和可持续发展具有重要意义.
[1]Gaedke U.Functional and taxonomical properties of the phytoplankton community:interannual variability and response to re-oligotrophication[J].Archiv für Hydrobiologie,1998,53:119-141.
[2]Litchman E,Klausmeier C A,Schofield O M,etal.The role of functional traits and trade-offs in structuring phytoplankton communities:scaling from cellular to ecosystem level[J].Ecology Letters,2007,10(12):1170-1181.
[3]Boyer T,Polasky S.Valuing urban wetlands:a review of non-market valuation studies[J].Wetlands,2004,24(4):744-755.
[4]Reynolds C S.The ecology of phytoplankton[M].Cambridge:Cambridge University Press,2006.
[5]Tao X.Phytoplankton biodiversity survey and environmental evaluation in Jia Lize wetlands in Kunming City[J].Procedia Environmental Sciences,2011,10:2336-2341.
[6]VerasztóC,Kiss K T,Sipkay C,etal.Long-term dynamic patterns and diversity of phytoplankton communities in a large eutrophic river(the case of River Danube,Hungary)[J].Applied Ecology and Environmental Research,2010,8(4):329-349.
[7]邓建明,徐彩平,陈宇炜,等.太湖流域主要河道浮游植物类群对比研究[J].资源科学,2011,33(2):210-216.
[8]林施泉,邵晓阳,姜丹,等.福建省木兰溪流域浮游植物群落结构特征[J].湿地科学,2013,11(1):48-53.
[9]陆欣鑫,刘妍,范亚文.呼兰河湿地夏、秋两季浮游植物功能分组演替及其驱动因子分析[J].生态学报,2014,34(5):1264-1273.
[10]董鸣,王慧中,匡廷云,等.杭州城西湿地保护与利用战略概要[J].杭州师范大学学报:自然科学版,2013,12(5):385-390.
[11]金相灿,屠清瑛,章宗涉,等.湖泊富营养化调查规范[M].2版.北京:中国环境科学出版社,1990.
[12]胡鸿钧,魏印心.中国淡水藻类:系统、分类及生态[M].北京:科学出版社,2006.
[13]曾呈奎,毕列爵,陆保仁,等.藻类名词及名称[M].2版.北京:科学出版社,2005.
[14]周凤霞,陈剑虹.淡水微型生物图谱[M].北京:化学工业出版社,2010.
[15]陈茜,吴斌,邵卫伟,等.浙江省主要常见淡水藻类图集[M].北京:中国环境科学出版社,2010.
[16]Danilov R,Ekelund N G A.The efficiency of seven diversity and one similarity indices based on phytoplankton data for assessing the level of eutrophication in lakes in central Sweden[J].Science of the Total Environment,1999,234(1/2/3):15-23.
[17]Zhu W,Pan Y,Tao J,etal.Phytoplankton community and succession in a newly man-made shallow lake,Shanghai,China[J].Aquatic Ecology,2013,47(2):137-147.
[18]Hamilton P B,Lavoie I,Ley L M ,etal.Factors contributing to the spatial and temporal variability of phytoplankton communities in the Rideau River(Ontario,Canada)[J].River Systems,2011,19(3):189-205.
[19]Nirmal Kumar J I.Phytoplankton composition in relation to hydrochemical properties of tropical community wetland,Kanewal,Gujarat,India[J].Applied Ecology and Environmental Research,2011,9(3):279-292.
[20]施练东,竺维佳,张俊芳,等.亚热带水库浮游植物群落结构季节演替及其春季水华成因分析:以浙江汤浦水库为例[J].水生态学杂志,2013,34(2):32-38.
[21]张华,胡鸿钧,晁爱敏,等.浙江紧水滩水库浮游植物群落结构季节变化特征[J].生态学报,2013,33(3):944-956.
[22]杨亮杰,余鹏飞,竺俊全,等.浙江横山水库浮游植物群落结构特征及其影响因子[J].应用生态学,2014,25(2):569-576.
[23]Li Z,Wang S,Guo J S,etal.Responses of phytoplankton diversity to physical disturbance under manual operation in a large reservoir,China[J].Hydrobiologia,2012,684(1):45-56.
[24]Reynolds C S,Huszar V,Kruk C,etal.Towards a functional classification of the freshwater phytoplankton[J].Journal of Plankton Research,2002,24(5):417-428.
[25]Fornarelli R,Antenucci J P,Marti C L.Disturbance,diversity and phytoplankton production in a reservoir affected by inter-basin water transfers[J].Hydrobiologia,2013,705(1):9-26.
[26]Laskar H S,Gupta S.Phytoplankton community and limnology of Chatla floodplain wetland of Barak valley,Assam,North-East India[J].Knowledge and Management of Aquatic Ecosystems,2013,411(6):1-14.
[27]Ma Y,Li G B,Li J,etal.Seasonal succession of phytoplankton community and its relationship with environmental factors of North Temperate Zone water of the Zhalong Wetland,in China[J].Ecotoxicology,2014,23(4):618-625.
[28]Wang X,Wang Y,Liu L S,etal.Phytoplankton and eutrophication degree assessment of Baiyangdian Lake Wetland,China[J].Scientific World Journal,2013,2013:Article ID 436965.
[29]Sommer U,Gliwicz M Z,Lampert W ,etal.The PEG-Model of seasonal succession of planktonic events in freshwaters[J].Archives of Hydrobiology,1986,106(4):433-471.
[30]Li Y,Liu H,Hao J,etal.Trophic states of creeks and their relationship to changes in water level in Xixi National Wetland Park,China[J].Environmental Monitoring and Assessment,2012,184(4):2433-2441.
[31]史坚,廖欣峰,方晓波,等.西溪湿地四季水质时空变化及影响因子分析[J].环境污染与防治,2014,36(6):39-46.
