产γ-氨基丁酸益生菌的研究
2015-03-18邹建勋毛茂张丽宁李慧丹楼徐栋陈蔚青
邹建勋,毛茂,张丽宁,李慧丹,楼徐栋,陈蔚青
(浙江树人大学生物与环境工程学院,浙江杭州 310015)
产γ-氨基丁酸益生菌的研究
邹建勋,毛茂,张丽宁,李慧丹,楼徐栋,陈蔚青
(浙江树人大学生物与环境工程学院,浙江杭州 310015)
益生菌是对人体健康具有促进作用的活性微生物,具有食用安全性。从酸乳制品中筛选出一株具有产γ-氨基丁酸能力较高的菌株,经形态学观察、生理生化检验和16S rDNA序列分析,初步鉴定为短乳杆菌(Lactobacillus brevis),属益生菌范畴。经培养基和发酵条件优化,发酵液中γ-氨基丁酸的含量可达2.03 g/L。以该菌株为出发菌株进行紫外诱变处理,得到一株突变菌株,γ-氨基丁酸产量达2.31 g/L,与出发菌株相比提高了11.4%。经过多次传代,稳定性较好。研究结果有助于拓宽益生菌产品应用领域。
益生菌;短乳杆菌;γ-氨基丁酸;鉴定;优化;紫外诱变
γ-氨基丁酸(γ-aminobutyric acid,GABA)是由谷氨酸脱羧生成的非蛋白质组成型氨基酸。它参与多种代谢活动,具有很高的生理活性,是哺乳动物中枢神经系统一种主要的抑制性神经递质,在中枢神经系统中作为抑制性神经递质起作用[1-3]。GABA具有降血压、调节心律失常、促进精神安定、调节激素分泌、增进脑活力、营养神经细胞、改善更年期综合症等功效,在医疗保健方面有着重要作用[4-9]。
益生菌(probiotics)是指来源于宿主并对宿主健康有一定促进作用的活性微生物。目前,研究最多且被大量报道的益生菌主要是乳酸菌,包括双歧杆菌(Bifidobacterium)、乳酸杆菌(Lactobacillus)和一些球菌(Lactococcus,Streptococcus thermophilus)等。益生菌的主要生理功能有调节肠道菌群平衡、缓解乳糖不耐症、促进营养物质的吸收利用、调控胆固醇、抗击高血压、抗衰老和免疫激活、抗肿瘤等。由于益生菌产品对人体机能有这些调节作用,因此成为功能性食品研究的热点,其安全性亦得到公认,各种益生菌产品得到广泛认可与应用[10-11]。近年来有研究证明某些乳酸菌具有一定产GABA能力[12-15]。
本研究从酸乳等益生菌制品中分离、筛选具有产GABA能力的菌株,对菌株进行初步鉴定和培养条件优化;在此基础上,通过紫外诱变育种进一步提高菌株的GABA产量,以期拓宽益生菌产品应用新领域,使益生菌产品在促进人类健康方面发挥更好的作用。
1 材料与方法
1.1 材料
1.1.1 样品与试剂
各种酸乳制品、酸菜等益生菌食品,购自超市。γ-氨基丁酸(GABA)与谷氨酸(Glu)标准品购自上海生工生物技术有限公司(纯度99.9%以上);其他试剂均为国产分析纯。16S rDNA PCR引物由上海生工生物技术有限公司合成,正向引物F:5'AGA GTT TGA TCC TGG CTC AG 3',反向引物R:5'ACG GCT ACC TTG TTA CGA CTT 3'。
1.1.2 培养基
TYG固体培养基,GYP种子培养基,GYP发酵培养基,LB培养基[16-17]。
1.2 方法
1.2.1 GABA的检测方法
GABA的检测采用文献[18]所述改良纸层析法并经进一步优化。将发酵液8000 r/min离心15 min,取上清液用滤纸过滤,所得滤液为待测样品。层析展开剂组成为正丁醇∶冰醋酸∶水=4∶1∶3(v/v)。取2~10 μL待测样品点样,用γ-氨基丁酸标准品以及谷氨酸钠标准品作参比,0.1%~0.5%的茚三酮作为显色剂,层析结束后85°C干燥箱烘干15 min,用洗涤剂(0.1%硫酸铜∶75%乙醇=2∶38)洗脱,725N紫外分光光度计520 nm下测定吸光度,由标准曲线确定GABA含量。实验中研究了不同茚三酮浓度和样品点样量对标准曲线精度的影响,以确定优化的显色剂使用浓度和点样体积。
1.2.2 乳酸菌的分离及培养方法
1)菌株的分离与纯化:取1 mL或1 g样品,用无菌水梯度稀释。将稀释至10-4、10-5和10-6的样品稀释液各取0.1 mL,平板涂布培养2~3 d,再挑取单个菌落,以划线法在TYG培养基上纯化,转接至斜面培养基,37°C培养24 h。分离出来的菌株每2周转接1次,4°C冰箱低温保藏。
2)种子培养:试管中装入5 mL GYP液体培养基,挑取1环菌苔接入其中,37°C静置培养24 h。
3)深层发酵培养:取培养24 h的种子培养液,按3%接种量接种到发酵培养基,用保鲜膜封口,37°C静置培养。定时取发酵液进行相关分析。
1.2.3 基因组DNA提取与16S rDNA扩增与序列分析
取1.5 mL菌液,12000 r/min离心5 min,弃上清液。用适量TE溶液重悬,100°C水浴10 min 及0°C冰浴5 min后12000 r/min离心10 min,取上清液,获得基因组DNA,以此为模板进行16S rDNA扩增。
PCR反应条件:94°C预变性10 min;94°C变性1 min,56°C退火45 s,72°C延伸90 s,共30次循环;循环结束后72°C终延伸10 min。PCR产物经纯化后,送上海生工生物技术有限公司进行序列测定,获得菌株16S rDNA基因序列。
将菌株的16S rDNA序列与GenBank数据库中序列进行BLAST同源性检索,再利用MEGA 5.0软件采用邻接法(Neighbor-Joining)构建系统进化树,自展数(Bootstrap)为1000。
1.2.4 紫外诱变育种
将菌种活化后,取稀释后的菌悬液置于距离预热30 min的紫外灯30 cm处进行诱变,分别在30 s,60 s,90 s,120 s,150 s时,取0.1 mL的菌悬液稀释不同梯度涂布LB平板,立即用锡箔纸包裹避光,以防止光复活。倒置于30°C恒温箱培养48 h后统计平板上的菌落个数,以未经紫外照射的平板为对照,计算致死率。挑选平板上长出的菌落与出发菌株同时进行摇瓶发酵,经初筛、复筛,筛选出GABA产量较高的菌株。
2 结果与分析
2.1 改良纸层析法测定GABA含量相关参数
研究了不同显色剂浓度对实验结果的影响。将0.1%~0.5%的茚三酮溶解于展开剂中,不同浓度的茚三酮显色、洗脱后比色结果显示,0.4%的浓度较合适。不同点样量实验结果表明,当GABA标准溶液点样量在4 μL时,标准曲线R2值最大(R2=0.9957)。GABA标准曲线如图1所示。

图1 GABA样品的标准曲线
2.2 产GABA乳酸菌筛选
初筛时,参照GABA标准品层析显色结果,观察平行位置上样品显色状况,从而初步确定菌株是否具有产GABA能力。从多种酸乳制品、酸菜等样品中分离了100余株菌株,根据纸层析结果初步判定13株菌株具有一定产GABA能力。13株产GABA菌株的定量分析结果见表1,可见53号菌株产量最高,故选择作为出发菌株。该菌株筛选自市售酸乳制品。

表1 产GABA菌株的定量分析结果
2.353 号菌株的形态观察与生理生化鉴定
53号菌株的菌落形态特征为:菌落较小,乳白色,不透明,中间隆起,边缘整齐,表面湿润光滑。显微镜观察为杆菌,革兰氏染色阳性。其生理生化特征为:产乳酸;过氧化氢酶反应不产气体,阴性反应;对葡萄糖、蔗糖、麦芽糖、果糖、半乳糖5种糖类消化实验均呈阳性反应。根据形态和生理生化特征,此菌株符合乳酸杆菌特点。
2.453 号菌株的分子生物学鉴定
以53号菌株的基因组DNA为模板,其16S rDNA PCR产物琼脂糖凝胶电泳检测见图2。

图2 53号菌株的16S rDNA PCR产物琼脂糖凝胶电泳图
PCR样品纯化后送检测序,测序结果经BLAST比对结果发现与GenBank登记多株短乳杆菌(Lactobacillus brevis)的16S rDNA序列同源性为98%以上,系统进化树构建结果如图3。结合生理生化鉴定结果,可初步判断53号菌株为短乳杆菌,属于益生菌范畴,将其命名为L.Brevis 53。
2.5 培养基与发酵条件优化
2.5.1 不同碳源对GABA发酵的影响
以GYP发酵培养基为基础,分别采用葡萄糖、乳糖、蔗糖作为发酵培养基的初始碳源,研究对产GABA的影响,结果见图4,乳糖和蔗糖作为碳源发酵效果最好,葡萄糖作为碳源的发酵效果最差,从工业化生产和原料成本的角度出发可选择蔗糖作为GABA发酵的碳源。
2.5.2 不同氮源对GABA发酵的影响
以GYP发酵培养基为基础,分别采用胰蛋白胨、蛋白胨、牛肉膏、酵母膏作为发酵培养基的初始氮源,研究对GABA发酵的影响,结果见图5,蛋白胨作为氮源发酵效果最好。
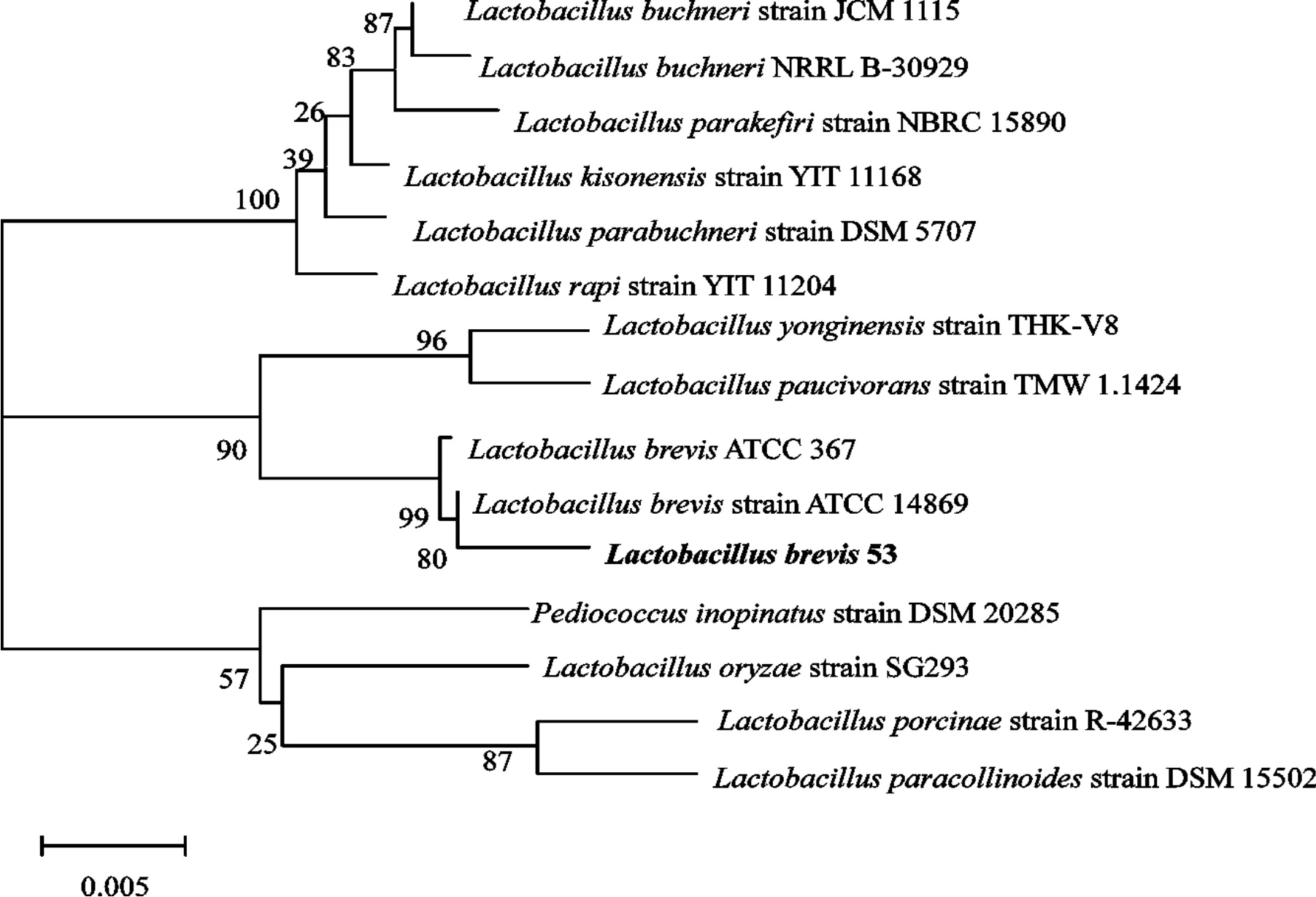
图3 53号菌株的系统进化树

图4 碳源对GABA产量的影响

图5 不同氮源对菌株产GABA的影响
2.5.3 L-谷氨酸浓度对GABA发酵的影响
分别取1 g/L,4 g/L,6 g/L,8 g/L,10 g/L,12 g/L 的L-谷氨酸钠在三角瓶中进行发酵,比较不同浓度的L-谷氨酸对GABA发酵的影响。由图6可知,随着L-谷氨酸浓度的提高,GABA的产量也稳步提高;当L-谷氨酸的浓度大于6 g/L时,GABA产量增加不明显,故选择6 g/L为L-谷氨酸钠合适的添加浓度。

图6 L-谷氨酸浓度对GABA发酵的影响
2.5.4 pH对发酵产GABA的影响
将菌种接入不同初始pH的发酵培养基后,37°C培养24 h后测定发酵液中GABA的产量。图7显示在初始pH为5.1时,发酵液中GABA的含量最高。
2.5.5 温度对发酵产GABA的影响
不同温度下发酵培养24 h后测定发酵液中GABA的含量。图8可见培养温度为37°C时发酵产GABA的量最高,说明菌体在37°C时体内的谷氨酸脱羧酶具有较高酶活,温度偏低或偏高对菌体的生长及酶活都不利。

图7 pH对发酵产GABA的影响
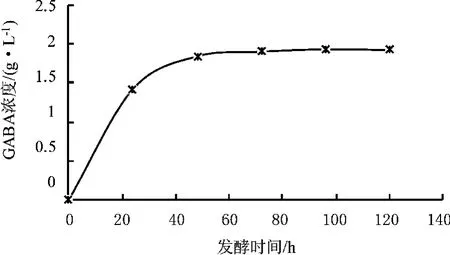
图10 pH 7.0下培养时间对GABA产量的影响
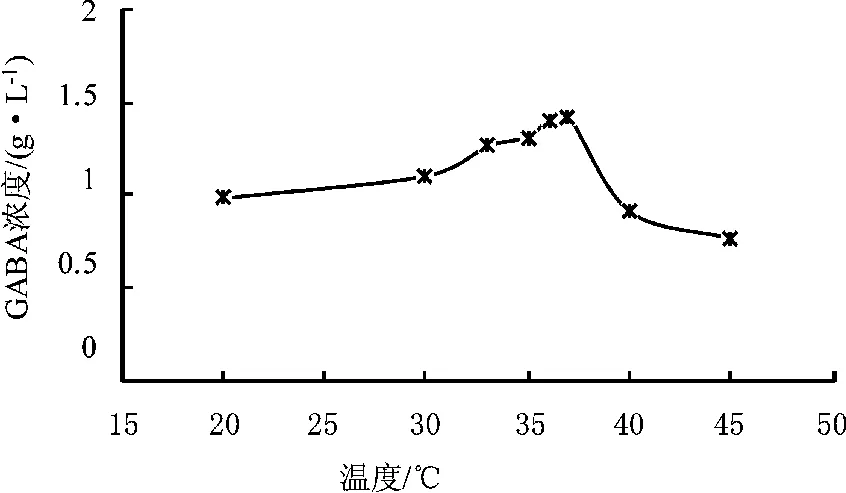
图8 温度对发酵产GABA的影响
2.5.6 培养时间对细胞生长与GABA产量的影响
在不同时间测定不同pH发酵液中细胞生长的OD值以及pH 7.0菌液的GABA含量。图9和图10表明随着发酵时间的延长,发酵液中GABA的产量和细胞数量不断增加,在48 h前增加幅度较大;48 h后继续培养GABA的增加幅度较小。考虑到发酵时间延长,生产周期也延长,成本增加,因此确定48 h作为生产GABA的发酵周期。

图9 培养时间对不同pH下细胞生长的影响
2.5.7 优化后的培养结果
通过以上实验对培养基碳源、氮源进行了优化,获得了优化的培养条件。经优化的培养基为(g/L):蛋白胨10.0,酵母粉10.0,蔗糖10.0,醋酸钠2.0,硫酸镁0.1,硫酸锰0.2,硫酸亚铁0.1,氯化钠0.1。优化的发酵条件为:培养基初始pH 5.1,培养温度为37°C,培养时间为48 h。利用优化后条件对L.Brevis 53进行培养,平行样品数为3,经检测样品GABA平均含量为2.03 g/L,与优化前相比提高了43.0%。
2.6 L.Brevis 53的紫外诱变结果
2.6.1 紫外诱变对菌株致死效应的影响
从图11可知,随着处理时间的延长,致死率增加,紫外照射时间为150 s时几乎全部死亡,表明该乳酸菌对紫外线敏感,致死效应明显。
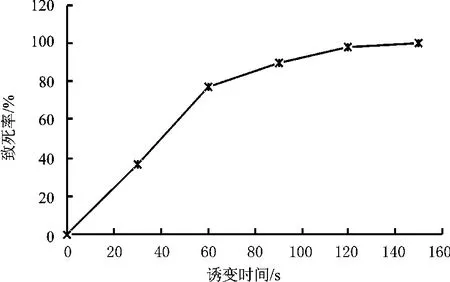
图11 紫外诱变菌株致死率曲线
2.6.2 紫外诱变处理后的效果
出发菌株经紫外照射诱变后,首先初筛出20个菌株,再经摇瓶复筛,选出发酵正向突变的1株菌株53-10,经紫外诱变处理后的γ-氨基丁酸产量达2.31 g/L,与出发菌株相比提高了11.4%。诱变后菌株发酵液GABA相对含量见图12(将出发菌株的GABA设定为100%)。
2.6.3 遗传稳定性试验结果
将筛选出来的突变菌种连续传代4次,分别检测GABA产量,结果(表5)表明菌株GABA产量波动不大,遗传性状稳定。
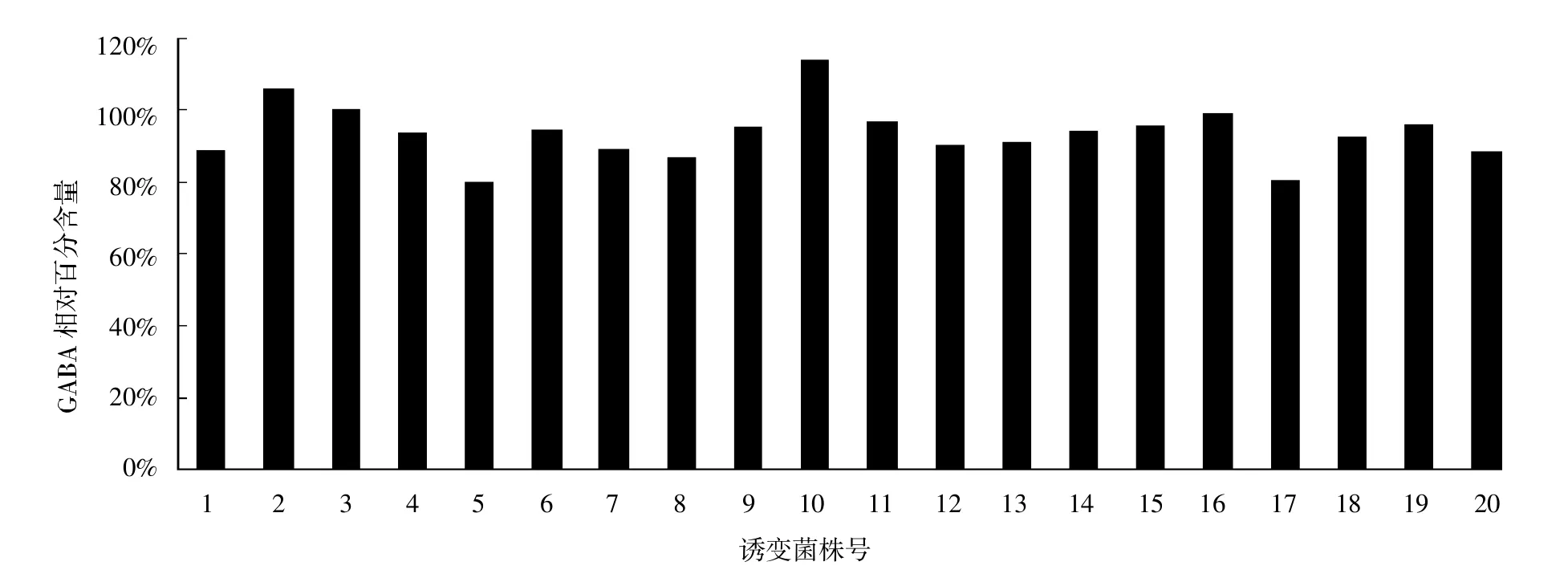
图12 诱变后菌株发酵液GABA相对含量柱状图
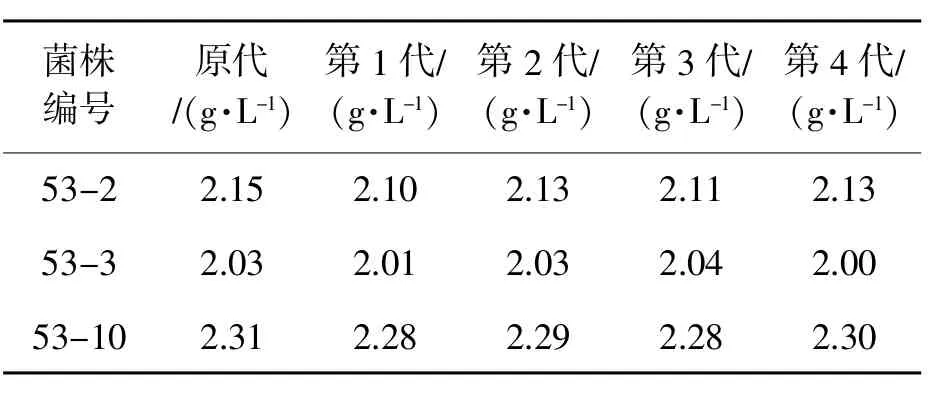
表5 诱变后GABA产生菌的遗传稳定性
3 讨论
本实验从酸乳制品中筛选出一株具有较高产γ-氨基丁酸能力的菌株,经形态学观察、生理生化检验和16S rDNA序列分析,初步鉴定为短乳杆菌(Lactobacillus brevis),属益生菌范畴。经培养基优化和发酵条件研究,发酵液中γ-氨基丁酸的含量可达2.03 g/L。以该菌株为出发菌株进行紫外诱变处理,得到一株突变菌株,γ-氨基丁酸产量达2.31 g/L,与出发菌株相比提高了11.4%。经过多次传代稳定性较好。益生菌是对人体健康具有促进作用的活性微生物,本文研究结果有助于拓宽益生菌产品应用领域,使益生菌产品在促进人类健康方面发挥更好作用。
[1]Benke D,Zemoura K,Maier P J.Modulation of cell surface GABA(B)receptors by desensitization,trafficking and regulated degradation[J].World Journal of Biological Chemistry,2012,3 (4)∶61-72.
[2]冯志彬,程显好,陈笑,等.γ-氨基丁酸产生菌分离筛选及发酵培养基初步优化[J].中国酿造,2008,27(21):51-53.
[3]韩雪,梁金钟.产γ-氨基丁酸乳酸菌菌株的分离鉴定[J].食品工业科技,2012,33(20):180-183.
[4]Xu J,Li C,Yin X H,et al.Additive neuroprotection of GABA A and GABA B receptor agonists in cerebral ischemic injury via PI-3K/Akt pathway inhibiting the ASK1-JNK cascade[J].Neuropharmacology,2010,54(7)∶1029-1040.
[5]秦江辉,周礼红,胡开成,等.高产γ-氨基丁酸的红曲霉菌株选育[J].江苏农业科学,2012,40(3):320-322.
[6]田小磊,吴晓岚,张蜀秋,等.γ-氨基丁酸在高等植物逆境反应中的作用[J].生命科学,2002,14(4):215-219.
[7]陈海军,林亲录,王婧,等.γ-氨基丁酸测定方法的研究[J].食品工业科技,2007,28(5):235-237.
[8]胡珊,王小波,戴甄,等.产γ-氨基丁酸红曲菌种的筛选及其发酵工艺条件研究[J].食品与发酵科技,2011,47(4):64-67.
[9]Binh T T T,Ju W T,Woo J J,et al.Optimization of γ-aminobutyric acid production in a newly isolated Lactobacillus brevis[J]. Biotechnology Letters,2014,36(1)∶93-98.
[10]刘刚,任文凯,熊霞,等.肠道与微生物相互作用体外研究模型进展与应用[J].动物营养学报,2013,25(8):1677-1682.
[11]周青,魏春,应向贤,等.产γ-氨基丁酸乳酸菌的筛选及发酵过程研究[J].食品与发酵工业,2011,37(5):26-31.
[12]仇婷,李海星,曹郁生.稀释率对短乳杆菌NCL912连续培养产γ-氨基丁酸的影响[J].食品与发酵工业,2010,36(12):68-71.
[13]Zhang L,Xiao Y X.Analytical research of cantonese guest of the government tea[J].Journal of Tea,2010,21(2)∶20-21.
[14]Takahashi T,Furukawa A,Hara S,et al.Isolation and characterization of sake yeast mutants deficient in γ-aminobutyric acid utilization in sake brewing[J].Journal of Bioscience and Bioengineering,2010,97(6)∶412-418.
[15]蒋冬花,后加衡.酸菜中高产γ-氨基丁酸乳酸菌的筛选[J].微生物学杂志,2007,27(1):35-39.
[16]朱晓立,邓毛程.产γ-氨基丁酸乳酸菌的分离筛选与微波诱变育种[J].中国酿造,2009,28(9):41-44.
[17]王超凯,刘绪,张磊,等.产γ-氨基丁酸乳酸菌的筛选及发酵条件初步优化[J].食品与发酵科技,2012,48(1):36-39.
[18]张晖,徐永,姚惠源.纸层析法定量测定米胚芽中的γ-氨基丁酸[J].无锡轻工大学学报,2004,23(2):101-103.
(责任编辑:朱小惠)
Study on γ-aminobutyric acid-producing probiotics
ZOU Jianxun,MAO Mao,ZHANG Lining,LI Huidan,LOU Xudong,CHEN Weiqing
(College of Biological and Environmental Engineering,Zhejiang Shuren University,Hangzhou 310015,China)
Probiotics are live micro-organisms with edible safety and beneficial to human health. A γ-aminobutyric acid(GABA)producing strain was isolated from yogurt samples.Based on 16S rDNA sequence,phylogenetic analysis,morphological and physiological characterization,the strain was identified as Lactobacillus brevi,which belongs to the category of probiotics.The yield of GABA reached 2.03 g/L after optimization of culture conditions and media components.Ultraviolet irradiation was used to treat the original strain.The accumulation of GABA was 2.31 g/L,which was increased 11.4%compared with that of original strain.The mutant GABA-producing strain was stable after several generations.These results showed that the application field of probiotic products could be broadened.
probiotics;Lactobacillus brevis;γ-aminobutyric acid(GABA);identification;optimization;ultraviolet irradiation mutagenesis
Q939.9
A
1674-2214(2015)01-0022-06
2014-11-20
邹建勋,(1992—),男,安徽亳州人,研究方向为微生物生物技术,E-mail:892380241@qq.com.通信作者:陈蔚青教授,E-mail:cwq135790@sina.com.
