CBF-β 辅助SIVagm Vif 蛋白诱导限制因子APOBEC3G 的降解
2015-03-09朱丹彤艾有为林跃智马建章王晓钧侯志军张明海
朱丹彤,艾有为,林跃智,马建章,王晓钧*,侯志军*,张明海*
(1.东北林业大学 野生动物资源学院,黑龙江 哈尔滨 150040;2.中国农业科学院哈尔滨兽医研究所 兽医生物技术国家重点实验室,黑龙江 哈尔滨 150001)
APOBEC3G(A3G)是近年来发现的一种重要的天然免疫慢病毒限制因子,通过诱导病毒cDNA 超突变,降低病毒感染性,发挥重要的抗病毒作用[1]。Vif(Virion infectivity factor)是除了马传染性贫血病毒(EIAV)以外,其他慢病毒均编码的一种病毒辅助蛋白,分子量为23 ku,是慢病毒在某些细胞中复制所必需的辅助蛋白。Vif 通过招募基于Cul5、Elongin B、Elongin C 和Rb 组成的Cullin5-ring ligase(CRL5)E3 泛素连接酶复合物与A3G 结合,诱导多种APOBEC3 蛋白的泛素化及降解,解除APOBEC3介导的抗病毒作用[2-3]。HIV-1 的研究发现,Vif 对APOBEC3G 降解作用还需要另外一种因子CBF-β 的参与。CBF-β 是RUNX 家族转录辅助因子,可与RUNX 家族的蛋白形成异二聚体,该复合物对于细胞分化具有重要功能。在HIV-1 感染过程中,CBF-β 的缺失会导致Vif 失去对A3G/F 降解作用[4-5]。CBF-β 广泛表达于各种细胞,其存在可以提高Vif的稳定性和可溶性,抑制Vif 的寡聚化。未寡聚化的Vif 更利于E3 泛素连接酶复合物的形成。同时,CBF-β 的表达可以提高Vif 与Cul5 的结合,从而有效的招募E3 泛素连接酶复合体并与之稳定结合,有利于Vif 对A3G 的降解作用。其中aa15-aa126 为CBF-β 发挥该功能的关键区域[6-7]。由此,CBF-β 在HIV-1 编码的VIf 对A3G 的降解作用是至关重要的。除HIV-1 之外,最近研究发现SIVmac(恒河猴)编码的Vif 蛋白对A3G 的降解作用也同样需要CBF-β 的辅助作用[8]。
在HIV-1 及SIVmac 的相关研究显示CBF-β 的重要作用[8]。但这一结果是否适用其他来源的慢病毒还不清楚。本研究通过CBF-β 对不同属的灵长类慢病毒来源的Vif 生化特性的改变,来探索CBF-β促进灵长类慢病毒Vif 的原因和机制,证明SIVagm编码的Vif 对A3G 的降解同样需要CBF-β 的辅助作用,并且其第38 位色氨酸对于该蛋白的活性具有关键作用。
1 材料和方法
1.1 重组质粒及细胞 pcDNA-HIV-1-Vif-HA、pcDNASIVagm-Vif、pcDNA-HIV-1-Vif W38S-HA、pcDNASIV-Vif W38S-HA、pVR-CBF-β-myc-flag、293T 细胞及293T siRNACBF-β细胞系均由中国农业科学院哈尔滨兽医研究所马传染病与慢病毒创新团队提供;人A3G(pcDNA-A3G-V5)以及非洲绿猴A3G(pcDNAA3Gagm-V5)表达重组质粒由中国农业科学院哈尔滨兽医研究所基础免疫创新团队郑永辉研究员提供。
1.2 主要试剂 Plasmid Mini Kit I 购自OMEGA 公司;HiPure Plasmid Maxiprep Kit 购自Invitrogen 公司;鼠抗HA、鼠抗beta-actin、Flag 抗体、V5 抗体和放线菌酮(CHX)均购自Sigma 公司;DyLightTM羊抗鼠IgG 购自ROCK-LAND 公司。
1.3 CBF-β 对SIVagm Vif 功能的影响 按照磷酸钙沉淀转染法,设立对照组和实验组2 个转染组,对照组将3 μg A3G、1 μg A3G 和1 μg pcDNAHIV-1-Vif-HA、1 μg A3G 和1 μg pcDNA-HIV-1-Vif-HA 及1 μg CBF-β 分别转染293T siRNACBF-β;实 验组将3 μg A3Gagm、1 μg A3Gagm 和1 μg pcDNASIV-Vif-HA、1 μg A3Gagm 和1 μg pcDNA-SIV-Vif-HA 及1 μg CBF-β 分别转染293T siRNACBF-β,同时用空载体pcDNA3.1 补平转染质粒总量差异。转染48 h后收集细胞并裂解,进行SDS-PAGE,将转印蛋白的PVDF 膜用5 %的脱脂乳封闭,分别以HA 抗体(1:10 000)、V5 抗体(1∶5 000)、Flag 抗体(1∶5 000)、actin 抗体(1∶10 000)为一抗,以DyLightTM羊抗鼠IgG 为二抗(1∶10 000),通过红外荧光扫描成像系统(YQ09-0052)检测各个实验组的蛋白表达情况。
1.4 CHX 对Vif 蛋白稳定性检测 通过磷酸钙转染法,设立两个实验组,对照组将3 μg pcDNA-HIV-1-Vif-HA 与CBF-β 质粒或者相应量的pcDNA3.1 质粒共转染293T 细胞;实验组将3 μg pcDNA-SIVVif-HA 与CBF-β 质粒或者相应量的pcDNA3.1 质粒共转染进293T 细胞。转染24 h 之后换成含有终浓度100 μg/mL CHX(放线菌酮)的完全培养液,分别在加入CHX 后的0 h、1 h、2 h、6 h 和12 h 收 获细胞,并将细胞裂解。进行SDS-PAGE,将转印蛋白的PVDF 膜用5 %的脱脂乳封闭,分别以HA 抗体(1∶10 000)、Flag 抗体(1∶5 000)、actin 抗体(1∶10 000)及DyLightTM羊抗鼠IgG 为二抗(1:10 000)进行孵育,对蛋白表达进行检测。
1.5 SIVagm Vif 功能试验 通过点突变技术把HIV-1 Vif 和SIVagm Vif 的38 位色氨酸(W)突变成丝氨酸(S)并且以Bam HⅠ和NotⅠ为酶切位点加入HA 标签,构建重组质粒pcDNA-HIV-1 Vif W38SHA 和pcDNA-SIV Vif W38S-HA 进行相应的实验。通过磷酸钙沉淀法,分别将3 μg pcDNA-A3G 与pcDNA-HIV-1 Vif W38S-HA、3 μg pcDNA-A3Gagm与pcDNA-SIVagm-Vif W38S-HA 转染293T 细胞。转染48 h 后收集细胞并裂解,进行SDS-PAGE,将转印蛋白的PVDF 膜用5 %的脱脂乳封闭,分别以HA抗体(1∶10 000)、V5 抗体(1∶5 000)、actin 抗体(1∶10 000)及DyLightTM羊抗鼠IgG 为二抗(1∶10 000)进行孵育,对蛋白表达进行检测。
2 结果
2.1 CBF-β 辅助SIVagm Vif 降解A3Gagm 的检测结果 分别采用CBF-β/SIVagm Vif/A3Gagm 3 种重组质粒和SIVagm Vif/A3Gagm 2 种重组质粒共转染293T 细胞并采用A3G 单转染及相关HIV 重组质粒组合作为对照组进行对比性分析。并采用western blot 检测目的蛋白的表达。结果显示,同CBF-β 对HIV-1 Vif 和A3G 的作用相同,在SIVagm 中,CBF-β 的表达组中Vif 蛋白的表达显著高于其他2组(图1A,图1B 第4 列),同 时Vif/SIVagm Vif 对A3G/A3Gagm 蛋白降解作用显著高于未加入CBF-β的对照组(图1A,图1B)。上述结果表明,SIVagm Vif 对A3Gagm 的降解也需要CBF-β 的辅助。
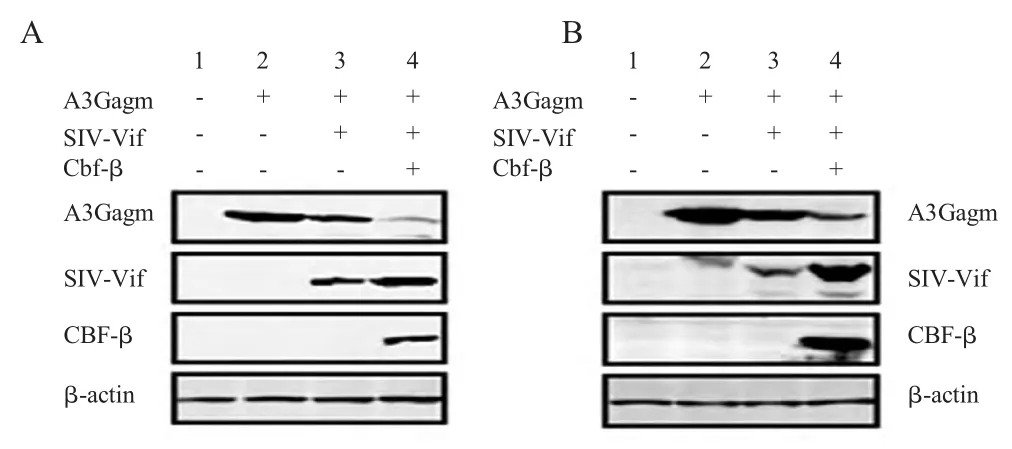
图1 在293T 细胞中共转染48 h 后检测Vif 介导的A3G 降解需要CBF-βFig.1 The degradation of Vif mediated A3G with the expression of CBF-β in 293T cells at 48 hrs post cotransfections
2.2 CBF-β 对SIVagm Vif 细胞内稳定性的检测结果 CHX 是一种在真核生物中对蛋白质生物合成过程有抑制效应的化合物。结果2.1 显示,加入CBF-β 的实验组SIVagm Vif 的表达显著高于其他2组。为进一步探索CBF-β 提高Vif 表达的机制,采用 将CBF-β 和Vif 表达质粒pcDNA-HIV-1-Vif-HA、pcDNA-SIV-Vif-HA 共转染293T 细胞后,通过CHX进行蛋白稳定性试验。由于CHX 能够抑制细胞内蛋白合成,但对蛋白降解没有影响。通过CHX 的处理后,在与单独表达SIVagm Vif 组相比,加入CBF-β 的实验组SIVagm Vif 的表达水平显著升高,同时与CHX 的作用时间呈负相关(图2)。表明,CBF-β 不仅促进HIV-1 Vif 在细胞内的稳定性,也能促进SIVagm Vif 在细胞内的稳定性。

图2 293T siRNACBF-β 细胞转染24 h 后 经CHX 处理后Vif 蛋白表达水平的检测Fig.2 The stabilization of Vif in presence of CHX in 293T siRNACBF-β from 24 hrs post the transfections
2.3 Vif W38S 点突变对CBF-β 的敏感性的检测结果 有研究表明HIV-1 Vif 中第38 位色氨酸(W)对HIV-1 Vif 与CBF-β 的相互作用具有关键作用。将HIV-1 Vif 的第38 位色氨酸(W)突变成丝氨酸(S)之后,HIV-1 Vif 丧失与CBF-β 之间的相互作用,CBF-β不再促进其细胞内稳定性以及Vif 介导的A3G 降解的功能。为了检测在SIVamg Vif 中与CBF-β 的关键结合部位是否也是第38 位色氨酸,将pcDNASIVagm-Vif W38S-HA 和CBF-β 共转染293T 细胞,对比pcDNA-SIVagm Vif-W38S-HA 单独转染的CHX-蛋白稳定性试验,发现CBF-β 对SIVagm W38S 突变体的稳定性同对HIV-1 Vif 同样没有促进作用。此外,CBF-β 也丧失了促进SIVagm W38S 细胞内稳定性的能力(图3A、B)。上述结果表明第38 位色氨酸对HIV-1 Vif、SIVagm Vif 与CBF-β 之间的相互结合作用至关重要。
2.4 Vif W38S 点突变对A3G 降解能力的检测结果将A3G 与pcDNA-HIV-1-Vif W38S-HA、A3Gagm 与pcDNA-SIVagm Vif W38S-HA 共转染293T 细胞后,收集样品进行western blot 检测。结果显示,野生型HIV-1/SIVagm Vif 蛋白能够有效地下调细胞内A3G/A3Gagm 的水平(图4A、图4B),而Vif W38S 突变组对A3G 的降解作用显著下降(图4A,图4B)。表明W38S 的突变不但改变了CBF-β 和Vif 的结合,同时也造成了Vif 诱导相应物种A3G 降解的能力。
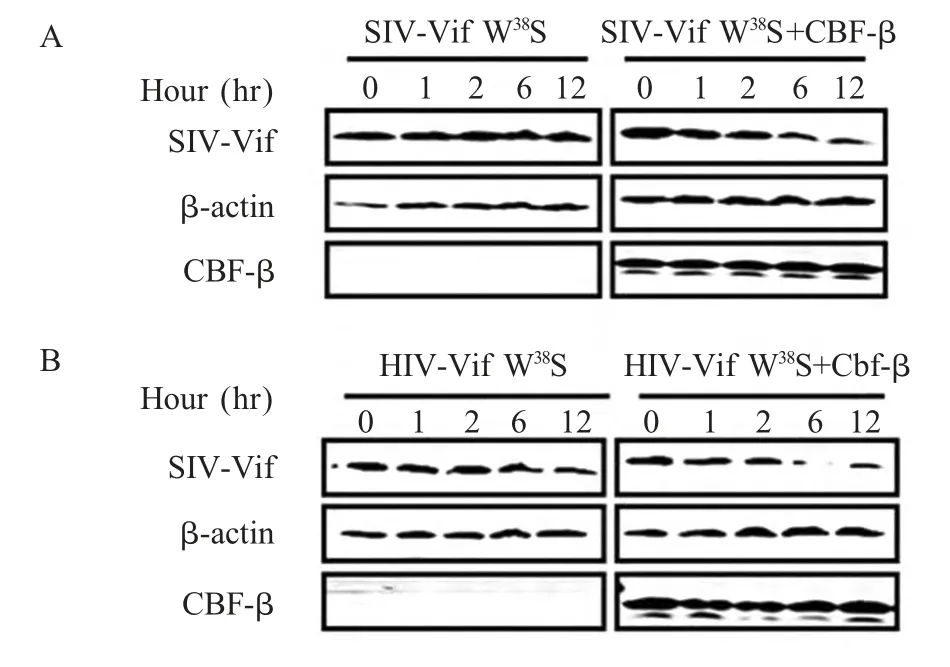
图3 Vif 突变体对CBF-β 结合活性的检测Fig.3 Detection of the binding activity of Vif mutant to CBF-β
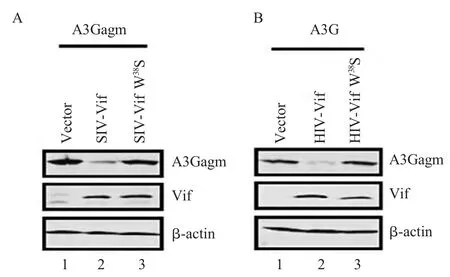
图4 Vif W38S 突变体对A3G 降解能力的检测Fig.4 Detection of A3G degradation activity with Vif W38S mutant
3 讨论
本研究显示,CBF-β 能够直接和Vif 结合影响其稳定性,减少其寡聚化水平。从而利于Vif 招募相应的因子,发挥对A3G 的降解作用。根据晶体结构显示,Vif 的蛋白结构包括一大一小2 个结构域[9]。CBF-β 能够与Vif 大的结构域发生紧密结合。而CBF-β 的羧基端则在Vif 的两个结构域间形成三明治结构。另一个结构域负责与CUL5-ELOC 结合。Vif 是处于复合体的核心位置。同时,结构显示与CBF-β-RUNX1 结合的区域为Vif 的α/β 区域,这一区域的关键氨基酸为第22 位色氨酸以及第38 位色氨酸[10]。这2 个氨基酸的破坏会打破CBF-β-Vif 及RUNX-CBF-β 的连接,从而影响复合物的形 成[11]。本研究也证实了这点。将HIV-1 Vif 与CBF-β 相互作用的第38 位色氨酸突变成丝氨酸之后,CBF-β 不再促进HIV-1 Vif 在细胞内的稳定性,该HIV-1 Vif突变体随即丧失了诱导A3G 降解的功能。由于不同种间的差异,SIVagm Vif 没有第22 位色氨酸,所以将SIVagm Vif 相应的第38 位色氨酸突变为丝氨酸之后,观察到CBF-β 不再促进SIVagm Vif 在细胞内的稳定性,该突变体丧失了诱导A3Gagm 降解的功能。恒河猴和非洲绿猴慢病毒编码的Vif 基因和CBF-β 作用的关键位点较为保守。因此,该位点将成为一个潜在的设计抗HIV-1 及SIV 药物的靶点。
[1]Wang Xiao-jun,Dolan P T.Biochemical differentiation of APOBEC3F and APOBEC3G proteins associated with HIV-1 life cycle[J].J Biolog Chem,2006,282(3):1585-1594.
[2]Fribourgh J L,Nguyen H C.Core binding factor beta plays a critical role by facilitating the assembly of the Vif-cullin 5 E3 ubiquitin ligase[J].J Virol,2014,88(6):3309-3319.
[3]Salter J D,Lippa G M.Core-binding factor beta increases the affinity between human Cullin 5 and HIV-1 Vif within an E3 ligase complex[J].Biochem,2012,51(44):8702-8704.
[4]Ai You-wei,Zhu Dan-tong,Wang Cui-hui,et al.Core-binding factor subunit beta is not required for non-primate lentiviral vif-mediated APOBEC3 degradation[J].J Virol,2014,88(20):12112-12122.
[5]Matsui Y,Shindo K,Nagata K,et al.Defining HIV-1 Vif residues that interact with CBFbeta by site-directed mutagenesis[J].Virology,2014,449:82-87.
[6]Zhou Xiao-hong,Han Xue.Dispersed and conserved hydrophobic residues of HIV-1 Vif are essential for CBFbeta recruitment and A3G suppression[J].J Virol,2014,88(5):2555-2563.
[7]Han Xue,Liang Wei-zi.Evolutionarily conserved requirement for core binding factor beta in the assembly of the human immunodeficiency virus/simian immunodeficiency virus Vif-cullin 5-RING E3 ubiquitin ligase[J].J Virol,2014,88(6):3320-3328.
[8]Wang Xiao-dan,Wang Xiao-ying,Zhang Hai-hong,et al.Interactions between HIV-1 Vif and human ElonginB-ElonginC are important for CBF-beta binding to Vif[J].Retrovirology,2013,10(1):94-99.
[9]Guo Ying-ying,Dong Li-yong.Structural basis for hijacking CBF-β and CUL5 E3 ligase complex by HIV-1 Vif[J].Nature,2014,505(7482):229-233.
[10]Ai You-wei,Ma Jianz-hang.Multiple lysines combined in HIV-1 Vif determines the responsiveness to CBF-beta[J].Biochem Biophys Res Commun,2015,457:385-390.
[11]Zhang W,Du J,Evans S L,et al.T-cell differentiation factor CBF-beta regulates HIV-1 Vif-mediated evasion of host restriction[J].Nature,2012,481(7381):376-379.
