松材线虫与拟松材线虫杂交及后代繁殖力1)
2015-03-07刘侃诚曹东霞田兴一李警保韩正敏
刘侃诚 曹东霞 田兴一 李警保 韩正敏
(南京林业大学,南京,210037)
责任编辑:程 红。
松萎蔫病是我国林业乃至世界林业的最主要病害。该病害是以松材线虫(Bursaphelenchus xylophilus)为主导,综合伴生细菌、媒介昆虫以及寄主松树的复杂病害系统[1]。和松材线虫同属于一个属的拟松材线虫(B.mucronatus),在各地松林内普遍存在,它们有时混生,有时单独出现[2]。近些年的研究发现,拟松材线虫在一些地区也表现出较强的致病性[3],尤其是法国拟松材线虫群体以及“M”型松材线虫群体,无论在致病性上还是形态上都表现为中间态[4]。用分子生物学的方法进行检测时,也发现了分子特征介于两者之间的线虫[5]。松材线虫与拟松材线虫作为2 个种的界限愈加模糊。由于这2 个种亲缘关系很近,又经常在病木中同时存在,2 个种是否能够杂交一直是研究的热点[6-10]。所以深入研究这一问题,不但能在伞滑刃属线虫的分类上,而且在松萎蔫病的流行和检疫等方面都有重要的价值。
本研究在收集的不同地理来源的松材线虫株系和拟松材线虫株系的基础上,在实验室内进行种间生物学杂交和回交试验,观察其杂交的可能性及杂交后代的繁殖能力,以确定松材线虫和拟松材线虫在自然界的杂交潜力,这对松萎蔫病的流行规律研究及病害的防治等方面都有重要意义。
1 材料与方法
1.1 供试线虫来源
虫株来源及编号见表1。各虫株都经过形态学和特异引物扩增详细鉴定,在分类上确认无误。

表1 供试线虫株系的编号、采集地和来源
1.2 未交配雌线虫的培养
将在灰葡萄孢(Botrytis cinerea)培养基上培养的线虫用无菌水洗出,在10×体视镜下用移液枪挑取单条大龄幼虫。显微镜下进一步镜检,确定其没有生殖器分化,放入装有2 mL 无菌水的无菌微型培养皿中。挑入1~2 根灰葡萄孢菌丝(饲喂线虫),25℃下培养2~3 d,待线虫长成成虫备用[11]。
1.3 线虫杂交方法
将培养好的单条未交配雌虫成虫和3 条异种雄成虫挑入长满灰葡萄孢的试管中,让其在灰葡萄孢斜面上交配并繁殖。同时做反交交配组合,每个组合重复20 次。25 ℃恒温暗培养,7 d 后,用无菌水洗下试管中的所有线虫。统计杂交后代的数量、杂交率、性比和死亡率[11]。
1.4 杂交后代繁殖力的测定
取F1 代培养好的未交配雌虫和1 条活跃雄虫继续在长有灰葡萄孢的试管内培养,做正交和反交组合,并以同种内交配产生的F1 代作为对照,每个组合重复20 次。25 ℃恒温暗培养。7 d 后,洗出各试管的F2 代,统计杂交后代的数量、杂交率、性比、畸形率和死亡率。以后杂交出的每一代(每隔7 d)都按此方法进行繁育[11]。
2 结果与分析
2.1 松材线虫和拟松材线虫杂交和种内交配
分别对来自浙江、云南、美国的松材线虫株系和来自中国、法国的拟松材线虫株系进行种间杂交和种内交配,培养7 d 后观察其F1 代的繁殖情况(表2)。结果显示,中国浙江松材线虫株系(ZJBX)和法国拟松材线虫株系(FrBm)杂交组合均有后代产生,且后代数量很多。和种内交配比较,该杂交组合表现出一定的杂交优势。ZJBX 和中国拟松材线虫株系(CBm)、中国云南松材线虫株系(YnBX)和FrBm这2 个杂交组合也均有后代产生,但后代数量都偏少。美国松材线虫株系(USBX)和FrBm 正反交产生的后代数量极少,死亡率高,杂交率低;反交后代无雄虫产生,因此认为USBX 和FrBm 不能顺利杂交。种内交配的所有组合,其后代繁殖都比较稳定,平均F1 代数量为17~34 条·管-1。
2.2 杂交后代的继代培养
为进一步了解各杂交后代的繁殖能力,对可进行种间杂交的ZJBX×FrBm、ZJBX×CBm 组合进行严格的继代培养,测定每代的繁殖数量、性比、死亡率等,以ZJBX、FrBm 的种内交配为对照(表3)。结果显示,ZJBX×FrBm 杂交组合,无论正交还是反交,同种内交配后代繁殖数量相比,其后代都具有较强的繁殖能力。ZJBX×CBm 组合后代的繁殖数量、性比(♀:♂)、死亡率较对照组ZJBX 及FrBm(种内交配)后代均具有一定的优势。ZJBX×FrBm、ZJBX×CBm 这2 个杂交组合同松材线虫和拟松材线虫的种内交配相比,继代培养具有更强的繁殖优势。
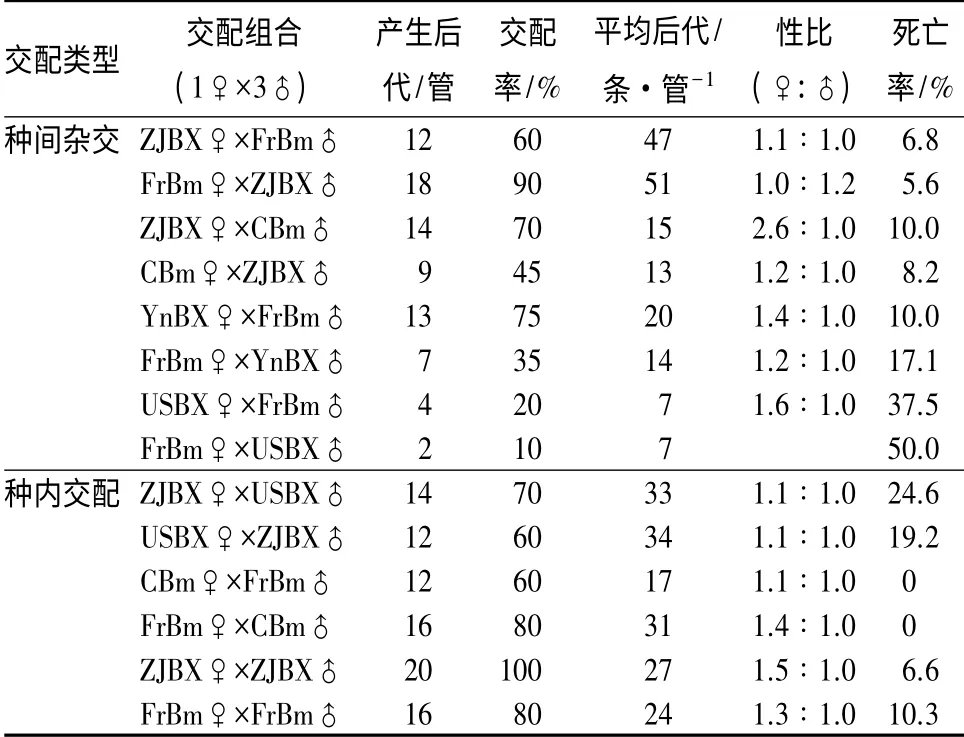
表2 松材线虫和拟松材线虫种间杂交和种内交配的F1 代

表3 松材线虫和拟松材线虫种间杂交后代继代培养结果
2.3 杂交后代同亲本的回交能力
ZJBX×FrBm、ZJBX×CBm 组合的正、反交杂交后代(F1、F3)与亲本进行严格的回交,对回交后繁殖数量、性比、死亡率等进行测定(表4)。结果显示,ZJBX×FrBm 正、反交组合F1 代更易于同亲本回交,但F3 代同亲本回交产生的后代数量高于F1 代,F3代更易同FrBm 亲本回交并产生大量的后代。ZJBX♀×CBm♂组合回交结果显示,F1 代同亲本的回交率均为50%,F1×ZJBX♂回交后代数量较少,且雄虫的数量比雌虫多近3 倍,死亡率较高。因此,回交后代继续繁殖下去的可能性不大。F1×CBm♂回交后代数量较多,雌虫的数量是雄虫的2 倍,无死亡率,具有很大的繁殖优势。F3 代同ZJBX 亲本不能回交。F3 代同CBm 亲本回交产生的线虫数量是F1代回交后代的2 倍。CBm♀×ZJBX♂组合中与拟松材线虫回交的组合的交配率和后代数量均略占优势。与2.1 中松材线虫和拟松材线虫的杂交和种内交配相比,ZJBX×FrBm、ZJBX×CBm 组合杂交后代同亲本回交具有一定的繁殖优势,并且更趋于同拟松材线虫回交。

表4 松材线虫和拟松材线虫种间杂交后代与亲本回交结果
3 讨论
关于松材线虫和拟松材线虫是否能够杂交一直在争论之中。持否定意见的是Mamiya[6]的研究,他们用日本松材线虫和日本拟松材线虫进行杂交,得出的结论是不能产生F2 代,F1 代也不能与其亲本进行回交。刘伟等[5]的研究虽然获得了F2 代和F3代,但后代数量少,他的结论也认为自然界存在杂交种的可能性不大。季宏铁[11]的研究结果表明,绝大多数的松材线虫和拟松材线虫杂交产生的后代在一代或多代之后就不再可育繁殖,只有法国拟松材线虫和北美松材线虫杂交才可以产生可育的后代。而Riga et al.[9]研究发现,西伯利亚拟松材线虫株系与北美松材线虫株系、德国拟松材线虫株系与日本松材线虫株系、日本松材线虫株系与法国拟松材线虫株系杂交,都获得了大量的杂交后代,认为这几个杂交组合都具有很高的杂交潜力。本研究中多个杂交组合均能杂交或回交成功,且有大量的后代产生,有些杂交组合还具有一定的杂交优势(如中国浙江松材线虫株系和法国拟松材线虫株系的杂交组合)。但美国松材线虫株系和法国拟松材线虫株系正反交产生的后代数量极少,死亡率高。说明不同的线虫株系组合,杂交和繁殖能力是不同的。
近年来,中国各地有很多地区拟松材线虫致使成片松林枯死的现象[12-15]。如2005年中国云南省华坪县发生大面积松树死亡现象,枯死松树达2.5万余株,经研究发现松树枯死主要原因为拟松材线虫所致[14]。1995年江西省德兴市马尾松大量死亡,经江西农业大学等专家实地调查,并结合症状观察和分离培养鉴定最终认定其为拟松材线虫为害所致[15]。本研究中,来自我国的松材线虫株系和我国的拟松材线虫株系之间是能够杂交、回交并产生可繁育的后代,说明这2 个株系之间有一定的杂交潜力。结合以上拟松材线虫致病现象,推测在我国松林内很可能有松材线虫和拟松材线虫的杂交种存在。
[1] 刘侃诚.松树萎蔫病生防细菌的研究[D].南京:南京林业大学,2013:6-10.
[2] 贲爱玲,过晔,韩正敏.松材线虫与拟松材线虫杂交及后代的致病性[J].林业科学,2010,46(8):114-119.
[3] 赵宇翔,徐正会,王峰,等.拟松材线虫发生区与未发生区云南松死亡研究[J].西南林学院学报,2003,23(1):62-66.
[4] 马承铸,钱振官,顾真荣,等.上海佘山发现黑松松材线虫(Bursaphelenchus xylophilus)M 型株系[J].上海农业学报,1996,12(1):56-60.
[5] 刘伟,杨宝君.松材线虫和拟松材线虫的杂交遗传差异研究[J].林业科学研究,1994,7(5):469-473.
[6] Mamiya Y.Interspecific hybridization between Bursaphelechus xylophilus and B.mucronatus(Aphelenchida:Aphelenchoididae)[J].Applied Entomology and Zoology,1986,21(1):159-163.
[7] Hajaukiewicz P T,Myers R T.Experimental crossing of selected isolates of Bursaphelenchus xylophilus and Bursaphelenchus mucronatus[J].Phytopathology,1988,78(11):1507-1508.
[8] De Guiran G,Bruguier N.Hybridization and phylogeny Nematologica of the pine wood nematode (Bursaphelenchus spp.)[J].Nematologica,1989,35:321-330.
[9] Riga E,Beckenbach K,Webster J M.Taxonomic relationships of Bursaphelenchus xylophilus and B.mucronatus based on interspeeific and intraspecific cross-hybridization and DNA analysis[J].Fundamental and Applied Nematology,1992,15(5):391-395.
[10] Bolla R I,Boschert M.Pine wood nematode species complex:interbreeding potential and chromosome number[J].Journal of Nematology,1993,25(2):227-238.
[11] 季宏铁.松材线虫(Bursaphelenchus xylophilus)与拟松材线虫(B.mucronatus)生物杂交学研究[D].南京:南京农业大学,2004:10-25.
[12] 姚晓华.江西省拟松材线虫分布简报[J].江西林业科技,1996(1):30,40.
[13] 胡楠,李茂军,唐春艳.双牌县拟松材线虫发生情况及防治方法[J].绿色科技,2013(9):146-147.
[14] 李桂香,李丹蕾,喻盛甫.华坪县松树萎蔫死亡主要原因初步研究[J].云南农业大学学报:自然科学版,2007,22(2):225-228.
[15] 方光明.拟松材线虫导致马尾松萎蔫[J].江西林业科技,1996(1):31,35.
