回想、熟悉性与启动在编码过程的认知神经机制*
2015-03-01叶晓红陈幼贞孟迎芳
叶晓红 陈幼贞 孟迎芳
(1三明学院教育与音乐学院, 福建 三明 365004) (2福建师范大学心理系, 福州 350007)
1 前言
在过去30年里, 内隐记忆(implicit memory)一直是认知领域的研究热点之一, 它反映的是一种自动的、不需要意识参与的记忆, 这种记忆与传统的、需要有意识回忆的外显记忆是功能性分离的。但以往对内隐和外显记忆的描述及区分多是针对个体在信息提取时的心理经验, 而与此相关但更为关键的问题是:形成记忆时, 大脑经历了怎样的加工过程?
研究编码神经机制的典型范式是相继记忆范式(subsequent memory performance paradigm), 即首先通过神经成像技术记录被试学习过程中的神经活动, 然后根据随后的记忆测验结果(如再认正确与错误)对学习项目进行分类(如随后记住与随后遗忘), 并探讨不同类别在编码过程中的大脑活动差异, 从而间接探讨记忆形成时大脑的认知加工机制。采用事件相关电位(event-related potentials, ERP)的研究业已证实, 随后外显提取的项目在学习过程中记录到的波幅比随后未提取出的项目更为正偏斜, 这种差异也称为“Dm (Differences based on subsequent memory performance)效应”, 它是记忆成功形成时大脑活动的特异性反映。但在内隐记忆背景下关于Dm效应的研究不多。
早期研究大多通过内隐测验来探讨内隐记忆Dm效应, 并与外显测验(如回忆、再认)中的Dm效应进行比较, 但所得结果并不一致(Paller, Kutas, &Maye, 1987; Paller, 1990; Friedman, Ritter, & Snodgrass,1996)。究其原因, 主要在于内隐测验中获得的效应是否完全反映了内隐记忆如启动的“纯加工”过程。实际上, 任何一个记忆测验中, 多种记忆过程都会同时对个体的行为产生作用(Voss & Paller, 2008)。因此研究者已不提倡单单根据测验模式来定义内隐和外显记忆, 而应把之区分为两种不同的加工过程。外显与内隐记忆加工会同时存在于任何一个测验中, 重要的是, 如何在测验中分离出外显记忆以及不带提取意识的内隐记忆加工。
已有一些研究曾尝试在单一测验中同时获得内隐和外显记忆的神经测量, 并探讨二者之间的关系。Schott, Richardson-Klavehn, Heinze和 Düzel(2002)首创词干补笔与再认相结合的两阶段测验方式, 在一个测验中同时获得外显记忆及不带再认的知觉启动。被试学习一系列视觉词后进行词干补笔测验, 要求先用学过的词来完成词干, 如果不行,就用想到的第一个词来完成词干, 补笔完成后回答该词是否来自学习阶段。根据补笔及随后的回答对学习阶段的项目进行分类。用学过的词完成了补笔,但随后却回答该词未学过, 该词干所对应的学习词为内隐记住项目; 用学过的词补笔, 同时回答该词学过的为外显记住项目; 用未学过的词完成补笔,同时也判断该词未学过的, 其词干对应的学习词为忘记项目。结果发现, 外显记忆表现为600~800 ms中央区以及900~1200 ms右额区正走向的Dm效应,即外显记住项目诱发的 ERP比忘记项目更为正走向, 而内隐记忆表现为200~450 ms中央顶区负走向的Dm效应, 即内隐记住项目诱发的ERP比忘记项目更为负走向, 表现出与外显记忆完全不同的神经反应。其fMRI结果表明, 预测随后外显记忆的神经关联表现在双内侧颞叶和左前额皮层的激活增强,而预测随后知觉启动的神经关联表现在双侧纹外皮层, 左梭状回和双侧下前额皮层的反应减少(Schott et al., 2006)。Wimber, Heinze和 Richardson-Klavehn(2010)采用类似的实验程序进一步证实, 编码过程中不同的额顶皮层网络预测着随后的内隐和外显记忆, 外显记忆主要表现在背侧后顶区和腹外侧前额皮层的激活, 而内隐记忆则表现在腹侧后顶皮层以及背外侧前额皮层与内侧额区的激活。
为了进一步验证内隐与外显记忆在编码阶段的分离现象, 孟迎芳近期(2012, 2013)采用与Schott等类似的两阶段测验程序及相继记忆范式, 比较了内隐与外显记忆在编码过程中的Dm效应差异。被试学习系列词或面孔之后进行二选一的迫选再认测验,如果两个都是未曾见过的, 也必须从中猜测出一个,之后回答刚才所选刺激是否为学习阶段出现过的,根据两次答案对学习阶段的刺激进行分类, 随后被选择并告知为见过的定义为“相继记住”; 随后被选择但告知是猜测的定义为“相继启动”; 随后未被选择, 即测验中选择了新刺激且告知为猜测的定义为“相继忘记”。结果发现, 在词实验中, 内隐记忆主要与颞区及中央区早期负走向的 Dm效应相联系, 而外显记忆主要与前额区正走向的 Dm效应相联系;在面孔实验中, 内隐记忆表现为400~500 ms额中央区负走向的Dm效应, 而外显记忆表现为400 ms开始顶区正走向的Dm效应。综合上述在编码上的这些研究, 虽然上述结果存在一些差异, 但我们发现,内隐记忆均表现为一种负走向的 Dm效应, 与之前研究中较多发现的正走向Dm效应存在着明显的差异, 且内隐记忆负走向 Dm 效应都出现得较早, 而外显记忆正走向 Dm效应都出现得较晚。由此, 我们不禁产生一种疑问, 负走向的Dm效应是否为内隐记忆在记忆形成过程中所特有的一种表现呢?这种负走向的Dm效应与外显记忆正走向Dm效应存在着什么样的关系?它们分别反映着怎样的认知加工机制?二者在记忆形成过程中分别扮演着怎样的角色呢?这一系列的问题引发了我们对内隐记忆形成过程中的加工机制的进一步探讨。
然而, 针对上述结果我们也发现了一些矛盾之处, 例如, 在Schott等(2002)研究中, 早期(200~450 ms)的负走向Dm效应在内隐记忆和外显记忆中有着一定的类似性, 虽然Schott等人对此并未做任何解释;孟迎芳(2013)的研究中也发现 200~300 ms中央区及600 ms开始的顶区负走向的Dm效应为两种记忆类型所共有。此外, 一般与外显记忆关联的内侧颞叶激活(Schott et al., 2002)在 Wimber等研究(2010)中却发现是与知觉启动相关联的。这些矛盾使得对研究结果的解释存在着不确定性, 究其原因,这些研究并未获得一致的内隐记忆Dm效应可能与熟悉性的混淆有关。根据记忆的双加工理论, 外显记忆可以划分为两个不同的加工过程:回想(recollection) 和熟悉性(familiarity)。回想反映的是对学习过的项目的特定信息的提取, 一般在再认测验中表现为“记得”反应, 熟悉性反映的是对学习过的项目和测验项目整体相似性的评估过程(Yonelinas,2002), 它是在缺乏来源信息的条件下产生的“知道”感觉。如Wimber等在对实验结果进行解释时曾提出, 启动项目可能易受到熟悉性或低自信度再认记忆的污染, 因为被试很可能只对那些能够回想起学习情节的项目或确定记得的项目做出“见过”的反应。按照这种假设, 那么预测随后启动的Dm效应可能同时包含了启动和熟悉性效应。因此, 在单一测验中进一步区分回想、熟悉性与启动是非常必要的。
基于此, 我们将通过两阶段的迫选再认测验与Dm 范式的结合, 在一个测验中同时获得回想、熟悉性及与不带提取意识的内隐记忆(知觉启动或概念启动)的Dm效应。为了能够同时探讨三者的Dm效应, 在本实验中, 我们将修改前人研究中的实验范式, 把迫选再认后的新/旧判断改为记住/知道/猜测判断:记住, 即选择是基于对学习阶段特定信息的提取; 知道, 即选择只是基于有点熟悉性的感觉,无法提取特定的信息; 猜测, 即完全没有记忆, 以至于该刺激“根本不是学习过的”, 只是迫于任务的要求进行了选择, 从而把学习项目区分为相继记住、相继知道、相继启动和相继忘记项目。记住和知道都意味着被试有主动的外显记忆, 而猜测则意味着外显记忆是完全缺失的。因此基于知觉或概念流畅性的无意识提取而产生的“正确猜测”行为提供了启动的行为测量, 而基于知觉或概念流畅性的有意识提取而产生的“知道感”提供了熟悉性的行为测量, 从而为进一步比较回想、熟悉性与启动之间的认知加工差异提供支持。
另外, Voss和Paller (2008)曾指出, 在任何一个内隐或外显测验中, 神经测量能够同时反映内隐和外显记忆加工, 因此如何利用实验参数选择性地获取不同记忆成分的操作是必要的, 否则无法确定与某种记忆加工关联的神经成像结果。因此我们认为,要想确定与三种记忆类型关联的神经分离现象, 需要提供能够在一个测验中区分不同记忆加工的相关行为测量。实验心理学已提供了好几个在行为上能够产生内隐和外显记忆分离的实验变量。例如,编码时的加工水平设置不会影响到随后的知觉启动, 但外显记忆行为会表现出加工水平差异; 编码时的干扰设置会减少随后的外显记忆行为, 但不会减少, 或只是稍微影响到随后的内隐记忆效应。基于此, 我们在编码阶段设计干扰变量, 以期通过编码阶段不同干扰变量的设置来证实单一测验中获得了多种不同的记忆加工类型。因而我们预测, 编码干扰会破坏回想和熟悉性Dm效应, 但不影响启动Dm效应。同时, 本研究可为进一步确定不同记忆类型的编码机制提供参考。
2 研究方法
2.1 实验设计
本项目包括两个实验, 实验 1为无干扰实验,实验2为编码干扰实验。
2.2 被试
实验 1选取了 15名大学生被试, 其中男生 8人, 女生7人, 实验2另外选取了15名大学生被试,其中男生9人, 女生6人, 所有被试均为某某大学学生, 年龄在20~25岁之间, 平均年龄22.1岁, 所有被试均为右利手, 视力或矫正视力正常, 无严重疾病史。实验结束后付给被试一定报酬。
2.3 刺激材料
(1)双字词
从北京语言学院教学研究所编著的《现代汉语频率词典》(1986年出版)选取低频汉语双字词480个, 词频为 2.3~12.2/百万, 平均词频 3.654/百万,所有词的意义均趋于中性化。把480个双字词随机分成4组, 每组120个词, 其中实验1与实验2使用相同的词语。在每个实验中, 2组双字词在学习阶段呈现, 剩余 2组作为新词用于测验阶段, 与学习阶段呈现过的旧词配对呈现。为了排除首因和近因效应, 每个实验都另外选取了 16个双字词作为填充词, 在学习阶段每组双字词呈现之前和之后出现,但结果不计入分析。
(2)干扰刺激
实验2中的干扰刺激由朝向不同的箭头符号构成朝向一致和不一致两类(见图 1), 干扰图片大小为3.18 cm × 2.91 cm, 在学习阶段与双字词同时呈现。

图1 干扰刺激样例
2.4 实验程序
2.4.1 实验1
实验过程中要求被试坐在隔音室内的沙发里,并注视计算机屏幕中央的注视点。计算机显示器的背景为黑色, 距离被试80 cm。要求被试用两手的食指分别按压游戏手柄的左键和右键进行相应的按键反应。
实验总共包括4个学习—测验组, 每组包括三个阶段:
(1)学习阶段:屏幕中央呈现一个 12.01 cm ×7.19 cm的白色方框, 单个双字词呈现在方框中央,字体大小为60, 每组内一半的双字词红色呈现, 另一半蓝色呈现, 要求被试对词的颜色进行判断, 红色按左键, 蓝色按右键。每组内的刺激随机呈现, 每个刺激呈现800 ms, 刺激间隔为(ISI) 1400 ± 200 ms。
(2)分心作业阶段:屏幕中央呈现358–2, 要求被试做一分钟的连续倒减2运算, 并大声的报告出来。
(3)迫选再认阶段:在测验阶段, 屏幕中央呈现一个13.36 cm × 6.27 cm的白色方框, 旧词与新词配对同时呈现在白色方框的左右两边, 旧词与新词的位置在被试内进行平衡。要求被试从中选择一个刚才见过的词语, 如果两个都没有见过, 也必须从中猜测出一个, 选择左边按左键, 选择右边按右键,词的字体大小为60, 所有的字体都为黑色, 两个词语的中间间隔大约 1 cm, 每个词配对呈现时间为2000 ms, 刺激间隔为(ISI) 1400 ± 200 ms。结束后屏幕再次出现按键的提示, 即对刚才选择出来的刺激表明自己的元记忆等级, 记住(百分之百的确定这个词语在学习阶段出现过, 并且能够非常清晰的回忆出所有的细节), 知道(仅仅只是觉得选择出来的词语很熟悉, 不能回忆出更多的细节信息), 猜测(对选择出来的词语没有任何感觉, 仅是为了完成任务, 迫不得已选择的)。三个不同元记忆等级通过游戏手柄的左右手按键及2键来控制, 记住用左手食指按左键, 知道用右手食指按右键, 猜测用右手拇指按2键。按键提示呈现时间为1400 ms, 要求被试在提示出现后尽可能地进行相应的选择。按键提示与下一个刺激之间的间隔时间为1400 ± 200 ms。
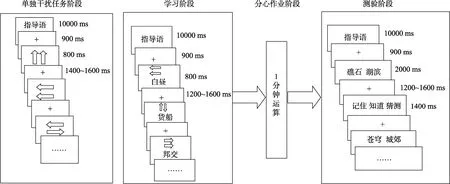
图2 编码干扰实验流程图
2.4.2 实验2
编码干扰实验分为两个阶段进行, 第一阶段进行单独干扰任务, 第二阶段干扰任务与记忆测验同时进行。(1)单独干扰任务阶段:要求被试对箭头的朝向进行判断, 箭头朝向一致用右手拇指按 2键,箭头朝向不一致不按键。每组内的刺激随机呈现, 每个刺激呈现800 ms, 刺激间隔为(ISI) 1500 ± 100 ms。具体实验程序图见图2。(2)干扰任务与记忆测验同时进行阶段:记忆实验2的程序与实验1类似, 不同之处仅在于学习阶段设置了干扰任务。在学习阶段, 干扰刺激与双字词同时出现, 呈现在双字词之上。要求被试既要进行双字词的颜色判断, 同时还要完成干扰任务。按键采用3键反应, 其中双字词颜色判断的按键方式与无干扰实验相同, 干扰任务采用“Go-no go”范式, 用右手拇指按2键对箭头朝向一致进行反应, 而箭头朝向不一致不反应。朝向一致与不一致的干扰刺激各占一半。在整个学习的过程中, 要求被试尽可能对两个刺激同时又快又准确的反应。每组内的刺激随机呈现, 每个刺激呈现800 ms, 刺激间隔为(ISI) 1400 ± 200 ms。具体实验流程图见图2。
在正式实验之前, 被试进行相应的练习, 熟悉整个程序, 练习的项目不进入正式实验。整个实验过程要求被试尽可能又快又准确的进行按键反应。实验程序使用 Presentation 0.71软件编制, 实验中使用的电脑为DELL Dimension 8200, 其显示器是15寸CRT, 分辨率为800×600, 刷新频率为75 Hz。
2.5 ERPs记录
采用 Neuroscan-64导脑电采集分析系统和Ag/AgCl电极帽, 连续记录学习阶段的EEG。电极位置采用 10-20扩展电极系统, 单级导联, 以头顶作为参考电极采集数据, 接地点在Fpz和Fz连线的中点, 另外有4个电极记录眼电:左眼框上、下侧2个电极记录垂直眼电, 两眼外侧的 2个电极记录水平眼电。滤波带通为0.05~100 Hz, A/D采样率为1000 Hz, 整个脑电采集过程中所有电极与头皮接触电阻均小于5 kΩ。
2.6 数据分析
对于实验所获得的数据, 均根据测验阶段两次选择的结果对学习阶段的刺激进行分类(图 3), 被试选择旧词, 再次表明元记忆等级为记住的为记住项目(记住); 选择旧词, 且随后表明元记忆为知道的为知道项目(知道); 选择了旧词但最后表明元记忆为猜测的为启动项目(启动); 被试选择新词, 且随后表明元记忆为猜测的, 与其相对应的旧词为忘记项目(忘记); 选择了新词, 且随后表明元记忆为记住或者知道的这两部分, 因为与本研究的目的无关, 不计入分析。

图3 数据分类示意图
对学习阶段的脑电记录进行脱机叠加平均处理, 分析窗口为–200~1200 ms, 用–200~0 ms作为基线进行矫正。伴有眨眼、眼动、肌电等伪迹的数据均被排除, 排除标准为±75 μV。对每种条件下的词所诱发的 ERPs进行分类叠加, 根据头皮分布与电极位置之间的关系, 同时结合波形、地形图等,共选取 15个代表电极进行统计分析, 包括前额区(FP1、FPz、FP2)、额区(F3、Fz、F4)、中央区(C3、Cz、C4)、顶区(P3、Pz、P4)和枕区(O1、Oz、O2)。所有的数据分析使用SPSS 11.5软件包进行, 同时使用Greenhouse-Geisser矫正法。
3 研究结果
我们分别对学习阶段各类项目的反应进行叠加, 总共获得学习阶段4类项目的ERPs:记住、知道、启动和忘记。为了具体了解各种项目之间的差异, 根据前人的研究, 我们把记住、知道、启动项目分别与忘记项目进行比较, 以此来确定回想、熟悉性和启动的Dm效应。
总体来看, 这4类项目的ERP差异大约从刺激后200 ms开始, 因此我们从200 ms开始分析, 每100 ms为一分析时段。具体分析过程中首先对原始数据进行标准化处理(McCarthy & Wood, 1985), 然后对不同实验结果进行分时段分脑区对两类项目的 ERP平均波幅进行 2(项目类别:忘记 vs记住/知道/启动)×3(电极)的重复测量方差分析。
3.1 无干扰条件下回想、熟悉性和启动的Dm效应分析
(1)回想的Dm效应
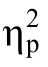
(2)熟悉性的Dm效应
方差分析结果表明, 知道和忘记项目的 ERP差异主要表现在 300~400 ms的前额区[F
(1,14) =4.46,p
= 0.053, η= 0.242]及 500~600 ms的顶区位置[F
(1,14) = 7.13,p
= 0.018, η= 0.337], 所有的差异均表现为知道诱发的 ERP比忘记更为正走向,即表现为正走向Dm效应。而在枕区, 200~300 ms发现了项目类别主效应[F
(1,14) = 6.06,p
= 0.027,η= 0.302], 但与前额区的差异趋势不同, 表现为知道诱发的 ERP比忘记更为负走向。具体差异波形图见图4。(3)启动的Dm效应

图4 无干扰条件下不同项目类别在编码阶段的ERPs比较
3.2 干扰条件下回想、熟悉性和启动的Dm效应分析
(1)回想的Dm效应
方差分析结果表明, 在不同的脑区及所有的时间段均未发现记住和忘记项目的 ERP差异。具体波形差异详见图5。
(2)熟悉性的Dm效应
(3)启动的Dm效应
方差分析结果表明, 在中央区600~800 ms发现项目类别的主效应[600~700:F
(1,14) = 4.03,p
= 0.064,η= 0.224; 700~800:F
(1,14) = 7.15,p
= 0.018, η=0.338], 启动诱发的ERP比忘记更为负走向。具体波形差异详见图5。综上所述, 我们可以发现, 在无干扰条件下,回想的Dm效应主要表现为从400 ms开始的右前额区正走向Dm效应以及800 ms开始的枕区负走向Dm效应, 而熟悉性主要表现为300~400 ms的前额区及500~600 ms的顶区正走向的Dm效应, 启动表现为700 ms开始的中央区负走向Dm效应。而在编码干扰条件的作用下, 这些Dm效应有着不同的变化, 首先我们没有发现与回想关联的明显 Dm效应, 而与熟悉性关联的正走向 Dm效应表现在600~800 ms的右额区, 与启动关联的负走向Dm效应表现在600~800 ms的中央区。
为了进一步探讨不同条件下发现的Dm效应是否具有类似性, 我们采用标准化的地形图分析方法(McCarthy & Wood, 1985; Picton et al., 2000)对熟悉性和启动的Dm效应分别进行2(实验条件:无干扰vs干扰)×62(电极)的混合方差分析。以往研究表明,标准化的地形图分析方法有助于判断不同条件下的地形图差异是否源于实验条件的不同所造成的神经信号激活程度的差异。如果不同实验条件所记录的大脑神经信号的激活是相同的, 那么所对应的地形图趋势应是相同的, 相反, 如果实验条件和电极存在着交互作用, 则可表明不同的干扰条件产生了不同的地形图差异(Alain, Achim, & Woods,1999)。结果发现, 对于熟悉性和启动的 Dm 效应,我们均未在相应时间段发现实验条件的差异, 表明两种条件下获得的熟悉性或启动Dm效应应该是类似的。

图5 干扰条件下不同项目类别在编码阶段的ERPs比较
4 讨论与分析
通过两阶段的迫选再认测验与Dm范式的结合,我们在一个测验中同时获得回想、熟悉性及启动的Dm 效应。结果发现, 这三种记忆类型关联的 Dm效应存在着差异, 更为重要的是, 在编码过程中设置的干扰变量对三种 Dm效应产生了不同的影响,即编码干扰破坏了回想的Dm效应, 但对熟悉性和启动的Dm效应影响相对较小。
4.1 与启动关联的负走向Dm效应
与之前研究结果类似(Schott et al., 2002; 孟迎芳, 2012, 2013), 本研究获得的启动Dm效应也表现为一种负走向的趋势, 即启动诱发的 ERP比忘记更为负走向, 这种趋势从刺激后700 ms开始, 直到记录结束, 在中央区表现的最为明显。重要的是,中央区这种负走向的Dm效应并未受到编码干扰的影响, 表现出对编码干扰的“免疫性”特征。可见,中央区这种负走向的Dm效应可能是与启动效应有着较多关联的一种神经表现。Schott等(2002)认为,这种负走向的Dm效应可能反映了在知觉和词汇层面上对词进行的有效加工, 这种加工有助于随后的知觉启动。Wimber等(2010)提出, 这种负走向的Dm效应反映了被试在编码阶段对信息的定向性注意, 虽然其无法保证随后的再认记忆, 却能有效地促进随后的知觉辨认。我们的结果进一步支持了这些解释。中央区负走向的Dm效应可能代表着刺激驱动的知觉加工(Whittlesea, 1993; Whittlesea,Jacoby, & Girard, 1990; Conroy, Hopkins, & Squire,2005; Meng, Ye, & Gonsalves, 2014), 这种知觉加工可以产生对该刺激的知觉流畅性, 虽然这种知觉流畅性无法支持随后的有意识提取, 但在迫选再认测验中, 它会提高被试选择旧项目的倾向(Voss &Paller, 2008, 2009; Voss, Baym, & Paller, 2008), 因为这些旧项目所引起的知觉流畅性要系统地高于新项目, 从而产生不带提取意识的内隐记忆效应。并且这种由刺激驱动的知觉流畅性不容易受到干扰的影响, 这与以往研究中一致发现的内隐记忆“抗干扰”性也是一致的(Mulligan, 1998; Benjamin Clarke & Bulter, 2008; Lozito & Mulligan, 2010), 因此我们认为, 中央区负走向的Dm效应可能是内隐记忆特有的一种ERP关联成分。
有意思的是, 在中央区我们也有发现与熟悉性关联的较微弱的Dm效应, 与启动Dm效应的时间窗口相似[700~800 ms:F
(1,14) = 3.80,p
= 0.072, η= 0.213], 但表现为一种正走向的Dm效应, 即知道诱发的 ERP比忘记更为正走向。这一现象为我们实验设计的有效性和意义提供了进一步的支持证据。正如我们在前言中所述, 以往研究中对内隐记忆Dm效应的探讨可能“参杂”着熟悉性的成分, 因此很可能与熟悉性关联的正走向Dm效应减弱或抵消了与启动关联的负走向Dm效应(Friedman et al.,1996)。而在本实验中, 我们进一步区分了熟悉性和启动效应, 并获得了与各自关联, 但趋势上完全相反的Dm效应, 这也进一步证实了熟悉性与启动在编码阶段的神经关联应该是存在差异的。此外, 在实验中也发现早期枕区负走向Dm效应(200~300 ms), 并且这一效应在三种记忆类型中都有发现。Wiggs和Martin (1998)曾提出, 早期与内隐记忆相关的负走向的 ERP新/旧效应主要与视觉皮质皮层相关(Slotnick & Schacter, 2006), 且这种早期效应被称为视觉感觉启动效应(Henson,Rylands, Ross, Vuilleumeir, & Rugg, 2004; Slotnick& Schacter, 2010), 反映了被试对刺激具体信息的知觉流畅性加工, 随后研究者也进一步证实, 枕区的波形差异仅仅代表视觉辨认的差异, 而没有涉及任何记忆的成分(Barense, Henson, Lee, & Graham,2010; O'Neil, Cate, & Kohler, 2009; Squire & Wixed,2011)。因此我们认为, 早期枕区负走向的Dm效应不太可能反映被试在编码过程中对词汇的加工差异, 可能更多反映了在编码过程中被试对词给予的一种视觉辨认。
4.2 回想与熟悉性的Dm效应存在分离
以往研究中探讨熟悉性和回想在编码加工上的差异, 主要采用相继记忆范式与“记得/知道”再认测验相结合, 以此来比较回想和熟悉性的Dm效应差异。例如Mangelsa, Pictonb和Craik (2001)发现, 相继记得刺激要比相继知道或相继忘记刺激引发一个更为正走向的ERP, 但相继知道与忘记之间并未发现明显的 ERP差异。Voss和 Paller (2009)结合自信度判断发现, 记住与高自信度知道都表现出明显的Dm效应, 且二者存在差异, 记住Dm效应比高自信度知道 Dm 效应有着更大的顶-枕区分布, 但对中自信度知道未发现Dm效应。可见在这些研究中, 对于熟悉性并未获得与之关联的可靠的Dm效应。对此, Mangelsa等(2001)曾将之归结为内隐记忆对熟悉性的“污染”, 也就是“知道”反应中可能还包含着基于知觉流畅性的“有效猜测(lucky guess)”成分。故此, 在本实验中我们加入了“猜测”判断, 将基于知觉流畅性的无意识提取而产生的“正确猜测”部分从熟悉性中分离出来。在此基础上,我们发现, 与熟悉性关联的 Dm效应主要表现在300~400 ms的前额区及500~600 ms的顶区。而回想的Dm效应主要表现在从400 ms开始的右侧前额区正走向Dm效应, 以及800 ms之后的枕区, 但趋势上为负走向的Dm效应。但更为重要的是, 在编码干扰的作用下, 与熟悉性关联的正走向Dm效应仍有存在, 主要表现在600~800 ms的右侧额区,但未发现任何与回想关联的有效Dm效应。由此推测, 熟悉性和回想在编码阶段的认知加工应该是存在着差异的。
关于熟悉性和回想之间的分离, 以往研究也进行了大量的探讨(Yonelinas, 2002; Woodruff, Hayama,& Rugg, 2006; Libby, Yonelinas, Ranganath, &Ragland, 2012; Moulin, Souchay, & Morrisb, 2013;Aly, Ranganath, & Yonelinas, 2014), 但大多针对提取时的认知加工, 发现二者有着不同的神经关联,回想通常与顶区500~800 ms (LPC)的新/旧效应关联, 而熟悉性则表现为额区300~500 ms (FN400)的新/旧效应。并且研究还发现, 在编码阶段的注意分散会减少回想顶区新/旧效应, 而对熟悉性额区新/旧效应不产生影响(Curran, 2004)。与提取阶段的这些研究结果类似, 我们在编码阶段也发现了同样的分离现象, 与熟悉性关联的Dm效应主要表现在额区, 且不受编码干扰的影响, 而与熟悉性相比, 回想还表现在从800 ms开始的枕区负走向Dm效应,但该效应易受编码干扰的影响。由此可见, 熟悉性和回想之间的加工差异在记忆形成的过程中就已经发生了。
Moscovitch (1992)曾提出的编码加工两阶段神经生理学模型, 认为在形成记忆的过程中, 额叶皮层首先对输入的信息进行精加工, 之后内侧颞叶-海马系统自动登记呈现给它的信息, 进行信息融合,以形成长期记忆(也见Buckner, Koutstaal, Schacter,& Rosen, 2000)。而我们实验中发现的与回想关联的持续枕区负走向Dm效应在以往研究中也有重复,如Mangelsa等人(2001)发现, 随后记住的词诱发的ERP要比随后知道或忘记的词更为负走向, 这种效应表现在下颞位置, 且持续时间较长(500~1500 ms), 研究者认为, 与回想关联的大脑后部持续的负走向Dm效应可能反映了对刺激细节表征的持续加工, 以支持随后对这些特定细节信息的有意识提取。综合这些结果, 我们认为, 在形成记忆的过程中, 额叶的加工能够支持随后的熟悉性反应。这样的加工对于随后的回忆反应可能是必要的, 但并不足以支持回忆反应, 因为回忆反应需要提取编码时的一些特定细节, 而这些可能反映在大脑后部持续负走向Dm效应上。
综上所述, 我们通过使用单一的迫选再认测验及Dm范式, 有效地获得了回想、熟悉性和启动在编码阶段的神经分离现象。启动主要与中央区相关联, 并且这种负走向 Dm 效应是内隐记忆特有的;熟悉性主要表现在额区正走向的Dm效应; 而回想则需要前额区正走向Dm效应和后部枕区持续性负波共同完成。
Alain, C., Achim, A., & Woods, D. L. (1999). Separate memory-related processing for auditory frequency and patterns.Psychophysiology, 36
, 737–744.Aly, M., Ranganath, C., & Yonelinas, A. P. (2014). Neural correlates of state-and strength-based perception.Journal of Cognitive Neuroscience, 26
, 792–809.Barense, M. D., Henson, R. N. A., Lee, A. C. H., & Graham, K.S. (2010). Medial temporal lobe activity during complex discrimination of faces, objects, and scenes: Effects of viewpoint.Hippocampus, 20
, 389–401.Buckner, R. L., Koutstaal, W., Schacter, A. L., & Rosen, B. R.(2000). Functional MRI evidence for a role of frontal and inferior temporal cortex in amodal components of priming.Brain, 123
, 620–640.Benjamin Clarke, A. J., & Bulter, L. T. (2008). Dissociating word stem completion and cued recall as a function of divided attention at retrieval.Memory, 16
(7),7
63–772.Curran, T. (2004). Effects of attention and confidence on the hypothesized ERP correlates of recollection and familiarity.Neuropsychologia, 42
, 1088–1106.Conroy, M. A., Hopkins, R. O., & Squire, L. R. (2005). On the contribution of perceptual fluency and priming to recognition memory.Cognitive, Affective, & Behavioral Neuroscience, 5
, 14–20.Friedman, D., Ritter, W., & Snodgrass, J. G. (1996). ERPs during study as a function of subsequent direct and indirect memory testing in young and old adults.Cognitive Brain Research, 4
, 1–16.Henson, R. N., Rylands, A., Ross, E., Vuilleumeir, P., & Rugg,M. D. (2004). The effect of repetition lag on electrophysiological and haemodynamic correlates of visual object priming.NeuroImage, 21
, 1674–1689.Lozito, J. P., & Mulligan, N. W. (2010). Exploring the role of attention during implicit memory retrieval.Journal of Memory and Language, 63
, 387–339.Libby, L. A., Yonelinas, A. P., Ranganath, C., & Ragland, J. D.(2012). Recollection and familiarity in schizophrenia: A quantitative review.Biological Psychiatry, 73
, 944–950.Meng, Y. F. (2012). ERP dissociation and connection between implicit and explicit memory at encoding.Acta Psychologica Sinica, 44
(1), 30–39.[孟迎芳. (2012). 内隐与外显记忆编码阶段脑机制的重叠与分离.心理学报, 44
(1), 30–39.]Meng, Y. F. (2013). Neural correlates at encoding of covert face recognition.Acta Psychologica Sinica, 45
(9), 935–943.[孟迎芳. (2013). 内隐面孔记忆的编码机制.心理学报,
45
(9), 935–943.]Meng, Y. F., Ye, X. H., & Gonsalves, B. D. (2014). Neural
processing of recollection, familiarity and priming at encoding: Evidence from a forced-choice recognition paradigm.Brain Research, 1585
, 72–82.Moulin, C. J., Souchay, C., & Morrisb, R. G. (2013). The cognitive neuropsychology of recollection.Cortex, 49
(6),1445–1451.McCarthy, G., & Wood, C. C. (1985). Scalp distributions of event-related potentials: An ambiguity associated with analysis of variance models.Electroencephalography and Clinical Neurophysiology, 62
, 203–208.Mulligan, N. W. (1998). The role of attention during encoding in implicit and explicit memory.Journal of Experimental Psychology: Learning, Memory, and Cognition, 24
, 27–47.Mangelsa, J. A., Pictonb, T. W., & Craik, F. I. M. (2001).Attention and successful episodic encoding: An eventrelated potential study.Cognitive Brain Research, 11
, 77–95.Moscovitch, M. (1992). Memory and working-with-memory:A component process model based on modules and central systems.Journal of Cognitive Neuroscience, 4
, 257–267.O'Neil, E. B., Cate, A. D., & Kohler, S. (2009). Perirhinal cortex contributes to accuracy in recognition memory and perceptual discriminations.Journal of Neuroscience, 29
,8329–8334.Paller, K. A. (1990). Recall and stem-completion priming have different electrophysiological correlates and are modified differentially by directed forgetting.Journal of Experimental Psychology: Learning, Memory and Cognition, 16
(5), 1021–1032.Paller, K. A., Kutas M., & Maye S. A. (1987). Neural correlates of encoding in an incidental learning paradigm.Electroencephalography and Clinical Neurophysiology, 67
,360–371.Picton, T. W., Bentin, S., Berg, P., Donchin, E., Hillyard, S. A.,Johnson, R. J. R., …Taylor, M. J.. (2000). Guidelines for using human event related potentials to study cognition:Recording standards and publication criteria.Psychophysiology, 37
, 127–152.Slotnick, S. D., & Schacter, D. L. (2006). The nature of memory related activity in early visual areas.Neuropsychologia, 44
,2874–2886.Slotnick, S. D., & Schacter, D. L. (2010). Conscious and nonconscious memory effects are temporally dissociable.Cognitive Neuroscience, 1
(1), 8–15.Squire, L. R., & Wixted, J. T. (2011). The cognitive neuroscience of human memory since H.M.Annual Review of Neuroscience, 34
, 259–288.Schott, B. H., Richardson-Klavehn, A., Heinze, H. J., & Düzel,E. (2002). Perceptual priming versus explicit memory:Dissociable neural correlates at encoding.Journal of Cognitive Neuroscience, 14
(4), 578–592.Schott, B. H., Richardson-Klavehn, A., Henson, R. N., Becker C., Heinze, H. J., & Düzel, E. (2006). Neuroanatomical dissociation of encoding processes related to priming and explicit memory.The Journal of Neuroscience, 26
(3),792–800.Voss, J. L., Paller, K. A. (2008). Brain substrates of implicit and explicit memory: The importance of concurrently acquired neural signals of both memory types.Neuropsychologia,46
(13), 3021–3029.Voss, J. L., Baym, C. L., & Paller, K. A. (2008). Accurate forced-choice recognition without awareness of memory retrieval.Learning & Memory, 15
, 454–459.Voss, J. L., & Paller, K. A. (2009). An electrophysiological signature of unconscious recognition memory.Nature Neuroscience, 12
, 349–355.Wimber, M., Heinze, H. J., & Richardson-Klavehn, A. (2010).Distinct frontoparietal networks set the stage for later perceptual identification priming and episodic recognition memory.The Journal of Neuroscience, 30
(40), 13272–13280.Woodruff, C. C., Hayama, H. R., & Rugg, M. D. (2006).Electrophysiological dissociation of the neural correlates of recollection and familiarity.Brain Research, 1100
, 125–135.Whittlesea, B. W. A., Jacoby, L. L., & Girard, K. (1990).Illusions of immediate memory: Evidence of an attributional basis for feelings of familiarity and perceptual quality.Journal of Memory & Language, 29
, 716–732.Wiggs, C. L., & Martin. A. (1998). Properties and mechanisms of perceptual priming.Current Opinion in Neurobiology,8
(2), 227–233.Whittlesea, B. W. A. (1993). Illusions of familiarity.Journal ofExperimental Psychology: Learning, Memory, & Cognition,19
, 1235–1253.Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30 years of research.Journal of Memory and Language, 46
, 441–517.