泰勒虫重组抗原Tams1-spag 基因原核表达与纯化
2015-02-27米晓云马文戈郭会玲苗书魁汪丽群魏玉荣新疆畜牧科学院兽医研究所新疆乌鲁木齐830000新疆维吾尔自治区动物卫生监督所新疆乌鲁木齐830063新疆农业大学动物医学学院新疆乌鲁木齐83005
任 方,易 忠,米晓云,魏 婕,马文戈,郭会玲,苗书魁,汪丽群,王 延,薛 英,黄 炯,魏玉荣*(.新疆畜牧科学院兽医研究所,新疆乌鲁木齐830000;.新疆维吾尔自治区动物卫生监督所,新疆乌鲁木齐830063;3.新疆农业大学动物医学学院,新疆乌鲁木齐83005)
泰勒虫重组抗原Tams1-spag 基因原核表达与纯化
任方1,3,易忠1,米晓云1,魏婕1,马文戈1,郭会玲1,苗书魁1,汪丽群2,王延1,薛英1,黄炯1,魏玉荣1*
(1.新疆畜牧科学院兽医研究所,新疆乌鲁木齐830000;2.新疆维吾尔自治区动物卫生监督所,新疆乌鲁木齐830063;3.新疆农业大学动物医学学院,新疆乌鲁木齐830052)
摘 要:为了在低成本的情况下,得到高浓度、高纯度的环形泰勒虫表面重组抗原的融合蛋白,设定表达过程中不同的OD600nm、IPTG浓度与诱导时间,探索最佳表达条件。通过KCl染色法切胶后分别经透析袋电洗脱法与快速离心法回收目的蛋白。对上述试验的蛋白样品进行SDS-PAGE和Western blot检测。结果表明,在OD600nm=1时加入浓度为1mmol/L IPTG,过夜诱导时,重组蛋白的表达量最大。在KCl染色法切胶回收目的条带的基础上,电洗脱法与快速离心法均能得到同等浓度的高纯度目的蛋白,经Western blot检测重组蛋白的生物活性没有改变,仍具有良好的反应原性。
关键词:环形泰勒虫;重组蛋白;原核表达;蛋白纯化
环形泰勒虫病是由寄生于动物红细胞、淋巴细胞及巨噬细胞内的环形泰勒虫(Theileria annulata)引发的一种蜱传急性、热性血液原虫病。此病呈地方流行性,表现为急性经过。不仅发病率与病死率较高,当病畜耐过此病或治愈后,体内仍存在低水平的虫体,导致抵抗力下降,或其他疾病暴发时再次诱发此病[1]。环形泰勒虫病对畜牧业危害很大,被世界动物卫生组织(OIE)列为必须报告的动物疾病,被我国列为二类疫病[2]。裂殖子表面抗原(merozoite surface antigen,Tams1)与子孢子表面抗原(sporozoite antigen,SPAG1)已被确认具有较好的免疫原性,可作为诊断与预防环形泰勒虫病的理想候选疫苗[3-5],但由于环形泰勒虫存在表面抗原的多态性,使得单一的表面抗原不能完全免疫预防此病[6],研究串联表面优势抗原蛋白为筛选理想的候选抗原提供一条新的思路。
鉴于在原核表达中,诱导时菌液OD600nm值、诱导剂异丙基代β-D-半乳糖苷(IPTG)的浓度以及诱导时间等因素均会影响外源基因的高效表达[7],而原核表达中包涵体不可避免的产生影响蛋白的纯化。探索合适的诱导条件与包涵体纯化回收的方法可以降低生产成本,提高试验效率。
本文对环形泰勒虫表面优势抗原蛋白串联后,在大肠埃希菌中经诱导剂IPTG诱导表达,通过探索诱导表达时的最优条件,获得大量的目的蛋白。并运用KCl染色法切胶回收含有目的蛋白的胶条,对回收胶条分别使用透析袋电洗脱法与快速离心法回收目的蛋白。对此两种纯化后回收蛋白的方法进行比较,以期得到高浓度与纯度的重组融合蛋白,为今后进一步研究蛋白其他特性奠定基础。
1 材料与方法
1.1材料
1.1.1菌株、质粒及细胞 pMD-Tams1-TaSPSPAG1、PET-28a质粒与BL21(DE3)表达宿主菌由新疆畜牧科学院兽医研究所传染病研究室保存。
1.1.2主要试剂 凝胶配制试剂盒、4×SDS双色加样缓冲液为武汉博士德生物公司产品;透析袋为华美生物工程公司产品;Amicon Ultra-15超滤管为MILLIPORE公司产品;PEG8000为WOLSEN公司产品;BCA蛋白浓度测定试剂盒为Thermo公司产品。
1.2方法
1.2.1重组质粒的诱导表达 挑取鉴定正确的阳性转化菌落接种于3mL LB液体培养基(kan+)中,37℃220r/min振荡培养过夜,次日以1∶100将菌液接种于LB液体培养基(kan+)中继续振荡培养至OD600nm=0.4~0.8,加入IPTG至终浓度1 mmol/L,诱导表达4h。同时以诱导前的菌液与诱导后转化的PET-28a空载体作为对照。将培养好的菌液5 000r/min离心5min收集菌体,洗涤后加入灭菌的PBS缓冲液(pH7.4),悬浮菌体后充分的超声破碎菌体,待菌体澄清透明后加入上样缓冲液煮沸10 min,取10μL进行120 mL/L的SDSPAGE电泳检测分析,确认诱导表达出目的蛋白。
1.2.2不同条件时重组质粒的诱导表达
1.2.2.1不同OD600nm的诱导表达 挑取鉴定正确的阳性转化菌落接种于3mL LB液体培养基(kan+)中37℃、220r/min振荡培养过夜,次日以1∶100的体积将菌液接种于LB液体培养基(kan+)中继续振荡培养至OD600nm分别0.3、0.5、0.7、1时,加入IPTG至终浓度1mmol/L,诱导表达4h。取样品处理后进行120mL/L的SDS-PAGE电泳检测分析。
1.2.2.2不同IPTG浓度的诱导表达 挑取鉴定正确的阳性转化菌落接种于3mL LB液体培养基(kan+)中37℃、220r/min振荡培养过夜,次日以1 ∶100的体积将菌液接种于LB液体培养基(kan+)中继续振荡培养至OD600nm=1时分别加入IPTG,使其至终浓度为0.5、0.7、1、1.3、1.5、1.7、2 mmol/L,诱导表达4h。取样品处理后进行120 mL/L的SDS-PAGE电泳检测分析。
1.2.2.3不同时间的诱导表达 挑取鉴定正确的阳性转化菌落接种于3mL LB液体培养基(kan+)中37℃220r/min振荡培养过夜,次日以1∶100的体积将菌液接种于LB液体培养基(kan+)中继续振荡培养至OD600nm=1时加入IPTG至终浓度为1.3mmol/L,分别诱导表达2、3、4、5、6、7h并且进行过夜诱导,取样品处理后进行120mL/L的SDSPAGE电泳检测分析。
1.2.3重组蛋白的纯化与回收
1.2.3.1重组蛋白的纯化 将750μL超声破碎后的包涵体提取液处理后不插样品梳直接上样,按常规方法进行SDS-PAGE。电泳结束后用250mmol/L KCl染色10min,将染成银白色含目的蛋白的胶切割回收,用无菌PBS(pH 7.2)漂洗3次至胶条白色消失。
1.2.3.2电洗脱法回收重组蛋白 将含有目的蛋白的胶条放入处理好的透析袋中,封口后放入含Tris-甘氨酸缓冲液的水平电泳槽中70V2h进行电洗脱回收蛋白,之后将取出胶条的透析袋放入PBS(pH7.2)缓冲液中4℃放置12h~24h。用聚乙二醇8000(PEG8000)对含有目的蛋白的透析袋进行浓缩,将浓缩后的产物进行SDS-PAGE电泳检测分析后BCA浓度测定。
1.2.3.3快速离心法回收目的蛋白 将多次切割收集的含有目的蛋白的胶条研碎后收集到离心管中,加入PBS(pH7.2)缓冲液放于-20℃反复冻融3次,8 000r/min离心5min,取上清液后再加入PBS (pH7.2)缓冲液放于4℃过夜透析,8 000r/min离心5min取上清液,之后再放于4℃过夜透析,经过两次重复4℃过夜透析离心回收后,将此4次收集的上清液分别进行SDS-PAGE电泳检测分析。将收集的全部上清液在Amicon Ultra-15超滤管中6 000 r/min离心30min进行浓缩。将浓缩后的产物进行SDS-PAGE电泳检测分析后BCA浓度测定。
1.2.4回收后重组蛋白Western blot的检测 纯化后的重组蛋白转至NC膜后,用含50g/L脱脂牛奶的PBST封闭4℃过夜,以环形泰勒虫阳性血清(1∶100)为一抗,孵育2h,PBST缓冲液洗膜后,以HPR标记的兔抗牛IgG为二抗,孵育1h,用PBST缓冲液洗涤后进行DAB显色后拍照保存。
2 结果
2.1表达产物不同条件的SDS-PAGE电泳
取不同条件重组质粒诱导表达的样品进行SDSPAGE,经过考马斯亮蓝染色观察。结果表明,25ku ~35ku处存在一条特异性的蛋白条带,与预期大小一致,而对照组未出现此表达条带。发现当OD600 nm=1时加入IPTG至终浓度为1mmol/L,过夜诱导时,蛋白表达量最高(图1~图4)。

图1 重组蛋白的SDS-PAGE电泳结果Fig.1 SDS-PAGE analysis of the recombinant proteins

图2 不同OD600nm对重组蛋白表达量的影响Fig.2 The effect of different OD600nm on expression levels of recombinant proteins

图3 不同浓度IPTG对重组蛋白表达量的影响Fig.3 The effect of defferent IPTG concentrations on expression levels of recombinant proteins
2.2重组蛋白纯化与回收的SDS-PAGE电泳
2.2.1快速离心法回收目的蛋白的SDS-PAGE电泳 取反复冻融后的胶条,快速离心回收的样品、回收后经Amicon ultra-15超滤管浓缩的样品与3次过夜后离心收集的样品进行SDS-PAGE检验(图5)。表明反复冻融后快速离心吸取上清液并不能完全回收蛋白,经过4℃透析仍能析出少部分蛋白,在重复至第3次时析出的蛋白经考马斯亮蓝已经鉴定不出,可以忽略不计。
2.2.2电洗脱法与快速离心法回收重组蛋白的SDS-PAGE电泳 取透析袋电洗脱法与快速离心法浓缩后回收的样品进行SDS-PAGE,经过考马斯亮蓝染色观察试验结果,并对两种方法回收的蛋白浓度进行BCA浓度测定(图6)。结果表明,在运用KCl染色法切胶回收目的条带的基础上,使用电洗脱法与快速离心法均可回收得到高纯度的目的蛋白,且两种方法回收的蛋白浓度不相上下,电洗脱法与快速离心浓缩法得到目的蛋白浓度分别为2.06 mg/mL和2.01mg/mL。
2.3回收后重组蛋白Western blot的检测
用牛环形泰勒虫阳性血清抗体进行免疫印迹试验,与SDS-PAGE电泳胶相对应的位置出现特异性杂交条带,而对照组的表达产物则无此条带(图7),可以初步确定纯化回收后的重组蛋白的生物活性没有改变,仍具有良好的反应原性。

图4 不同诱导时间对重组蛋白表达量的影响Fig.4 The effect of time on expression levels of recombinant proteins
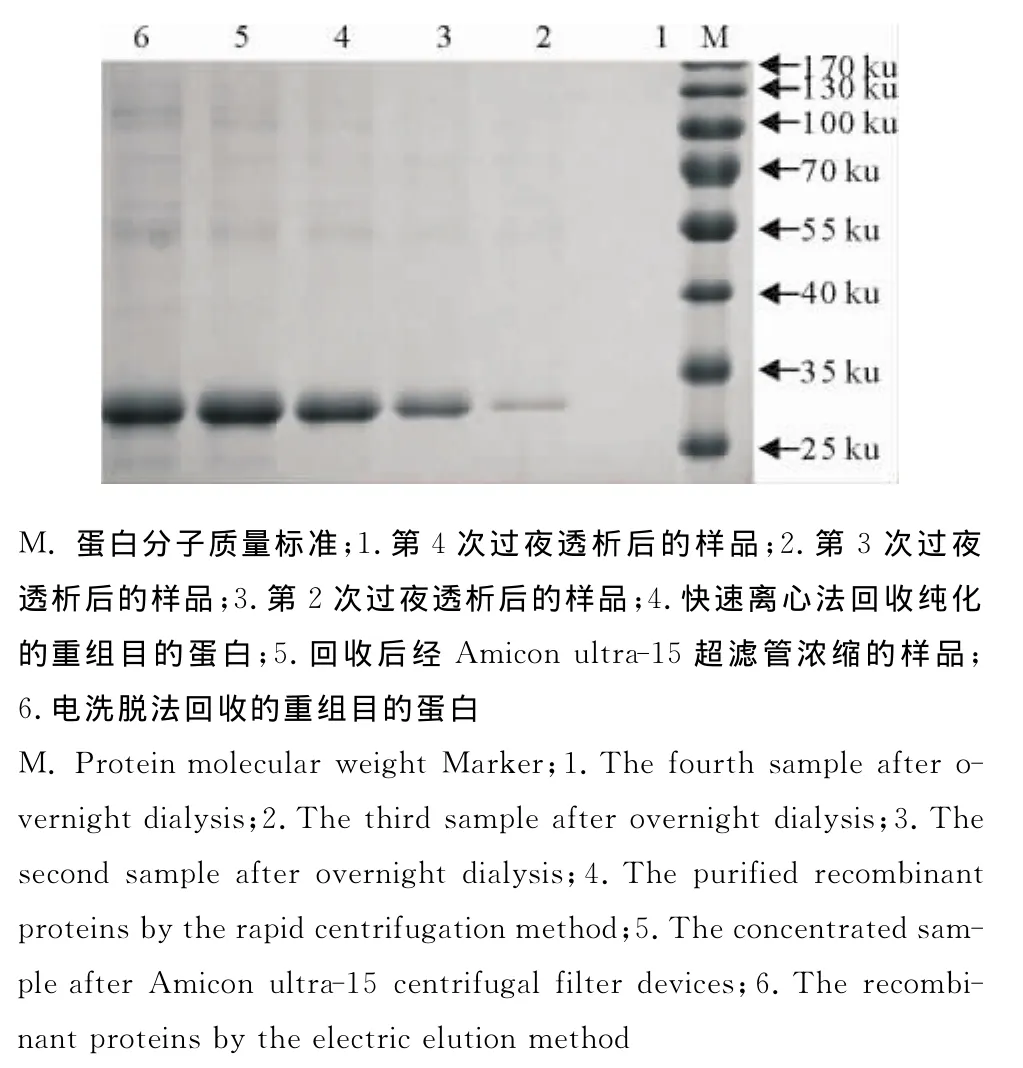
图5 重组表达蛋白的纯化Fig.5 The purification of recombinant proteins

图6 不同方法纯化的重组蛋白Fig.6 The purified recombinant proteins by different methods

图7 纯化的重组蛋白Western blot检测结果Fig.7 Western blot analysis of purified recombinant proteins
3 讨论
裂殖子表面抗原(Tams1)是环形泰勒虫感染红细胞时表面分泌的一种膜内蛋白,子孢子表面抗原(sporozoite antigen,SPAG1)是虫体发育至子孢子阶段时分泌的一种膜内蛋白,这两种表面抗原均具有良好的免疫原性且已被验证并在实际诊断中应用。但目前,以单一抗原为基础的诊断方法还存在着一些问题,例如抗原多态性的存在,各分离株间变异性的出现,不完全保护的产生等。而对环形泰勒虫不同发育阶段的串联表面优势抗原的研究则解决了这些问题,为筛选理想的候选抗原提供了一条新的思路。魏玉荣等[8-9]将环形泰勒虫表面抗原Tams1、Spag1、Tasp的高免疫原性区片段串联后用真核系统进行表达,并以此为基础,成功建立了以环形泰勒虫重组蛋白Tasp-Tams1-Spag为包被抗原的间接ELISA检测方法,验证了串联抗原重组蛋白的可行性。但串联蛋白低成本、高效率的制备仍需要进一步的探索。
在原核表达中,蛋白的表达量与纯化回收率对生产研究具有一定的影响,外源基因的高效表达会受到诱导时菌液OD600nm值、诱导剂IPTG的浓度以及诱导时间等因素的影响。探索合适的诱导条件可以降低生产成本,提高试验效率。在外源基因高效表达的基础上,对目的蛋白的纯化回收也大大影响到所需蛋白的生产效率。众所周知,外源基因在大肠埃希菌中高效表达,由于其表达部位的低电势与外源蛋白质分子的特殊结构,常常形成高密度、无活性的非折叠状的包涵体沉淀。目前,包涵体的提取方法大多是基于变性-复性过程的基础上,而影响包涵体复性的因素有许多,如蛋白的浓度与自身的理化性质,复性体系的离子强度、pH、温度等。不仅蛋白复性的条件难以把握,而且很多蛋白质在复性的过程中会难以控制的产生蛋白沉淀。一般蛋白质的复性效率只在20%左右。除此之外,变性-复性后目的蛋白的纯化也存在着杂蛋白干扰[10]。这些都严重影响目的蛋白的生产效率。为了解决包涵体蛋白的纯化问题,目前国内外学者应用切胶回收目的条带的方法纯化蛋白[11-12],学者们通过研究摒弃了会影响蛋白活性的考马斯亮蓝R-250染色切胶回收的方法,用KCl染色等负染法切胶回收含有目的蛋白的胶条[13]。对纯化后的蛋白一般采用电洗脱法、过夜透析高速离心法或反复冻融高速离心法回收目的蛋白[14-16]。
本文参照分子克隆与相关文章,探索在蛋白诱导表达过程中最合适的OD600nm、IPTG浓度与诱导时间。得知随着OD600nm的增大,诱导表达量明显增多,在OD600nm=1时加入IPTG,重组蛋白的表达量最大。在相同的诱导时间,当IPTG的浓度为1mmol/L后随着IPTG浓度的增加重组蛋白的表达量并不会增大,即1mmol/L的IPTG可以表达出大量蛋白。在保证其他诱导条件一定的情况下,发现重组蛋白的表达量会随着诱导时间的延长而增加,当过夜诱导时蛋白的表达量最高。对诱导表达出的大量蛋白经SDS-PAGE电泳后KCl染色切胶回收,可以完全去除杂蛋白的影响获得高纯度的目的蛋白。在回收过程中,电洗脱法与快速离心法回收的蛋白浓度不相上下,均可高效的回收蛋白。电洗脱法虽然可以将胶条中的蛋白完全析出,但浓缩时的效率不如离心法Amicon ultra-15超滤管。在快速离心法回收蛋白时,相同数量切胶回收的胶条分多次少量比大量冻融离心的回收率高,在第一次反复冻融提取含蛋白的上清后,经过两次过夜透析才可最大量的析出目的蛋白。实验室可以根据不同的情况而选用任意一种方法进行蛋白的回收。本试验为更快捷经济得到高浓度与纯度的环形泰勒虫重组融合蛋白提供了技术支持。
参考文献:
[1] 汪 明.兽医寄生虫学[M].北京:中国农业大学出版社,2008:332.
[2] 农业部畜牧兽医局.一、二、三类动物疫病释义[M].北京:中国农业出版社,2004:97.
[3] 蔺红玲,张继瑜,魏小娟,等.环形泰勒虫主要膜蛋白的研究进展[J].黑龙江畜牧兽医,2011,(05):22-24.
[4] Raiendran C,RavD D.Diagnosis of tropical bovine theileriosis by ELISA with recombinant merozoite surface protein of Theileria annulata(Tams1)[J].J Parasit Dis,2014,38(1):41-45.
[5] Darghouth M A,Boulter N R,Gharbi M,et al.Vaccination of calves with an attenuated cell line of Theileria annulataand the sporozoite antigen SPAG-1produces a synergistic effect[J]. Vet Parasitol,2006,142(1-2):54-62.
[6] Ghoneim A M,EI-Fayomy A O.Targeting tams-1gene results in underestimation of Theileria annulata infection in diseased cattle in Egypt[J].Acta Parasitol,2014,59(1):85-90.
[7] 郑进峰.环形泰勒虫TaSP基因的克隆、原核表达及PCR诊断方法的建立[D].河南郑州:河南农业大学,2013.
[8] 魏玉荣,易 忠,马文戈,等.泰勒焦虫表面抗原Tams1、Spag1 和Tasp基因的克隆与表达[J].新疆农业科学.2013,(11):2136-2142
[9] 魏玉荣,易 忠,马文戈,等.泰勒焦虫重组蛋白Tasp-Tams1-Spag1间接ELISA检测方法的建立[J].新疆农业大学学报. 2013,36(5):360-365.
[10] 冯延叶.变性蛋白复性装置研制及包涵体蛋白变性和复性技术研究[D].上海:华东理工大学,2013.
[11] 刘 镕,钟沁萍,蒋明森,等.一种获得高纯度包涵体蛋白的简便方法[J].中国寄生虫学与寄生虫病杂志.2011,29(5):399-401.
[12] Lee G H,Cooney D.The economics of inclusion body processing[J].Bioprocess Biosyst Eng,2006,29(2):73-90.
[13] 高慎阳,查恩辉,王 珅,等.一种“高性价比”切胶纯化原核表达蛋白的方法[J].中国农学通报,2010,26(22):24-26.
[14] 潘 辉.新疆卡拉库尔羊Fas基因的克隆、原核表达及抗原性分析[D].新疆塔里木:塔里木大学,2012.
[15] Bio-rad laboratories.Model 422electro-eluter instruction manual[M].US:Bio-rad Laboratories,2005:1-11.
[16] 李友娟,郭凤达,葛均青,等.鳗鲡疱疹病毒ORF51基因的原核表达及多克隆抗体的制备[J].福建农林大学学报,2014 (5):490-494.
Prokaryotic Expression and Purification of Theileria annulata Recombinant Antigen Tams1-spag Genes
REN Fang1,3,YI Zhong1,MI Xiao-yun1,WEI Jie1,MA Wen-ge1,GUO Hui-ling1,
MIAO Shu-kui1,WANG Li-qun2,WANG Yan1,XUE Ying1,HUANG Jiong1,WEI Yu-rong1
(1.Institute of Veterinary Medicine,Xinjiang Academy of Animal Science,Urumqi,Xinjiang,830000,China;2.Institute of Animal Health Supervision Station Inspection,Urumqi,Xinjiang,830063,China;
3.College of Veterinary Medicine,Xinjiang Agricultural University,Urumqi,Xinjiang,830052,China)
Abstract:To study the purification method of the of fusion protein of Theileria annulata surface recombinant antigens and to obtain high purity products at low cost,different parameters were set in order to obtain the optimum of expression.The experimental parameters included OD600nm,IPTG concentration induction time.Firstly,the target proteins were isolated by cutting the gel slices that contained the right bands which were stained by KCl solution.Secondly,the recycle protein were recovered by electrical elution and high centrifugation.Through SDS-PAGE and Western-blot,the experimental results indicated that the maximal quantity of expression of recombinant proteins was confirmed when IPTG concentration was 1mmol/Land OD600nm=1,the induced time was stay overnight.The two methods can obtain the same purified products and high purity.Western-blot assay indicated that the recombinant protein were not denaturated,and had good antigencity.
Key words:Theileria annulata;recombinant protein;prokaryotic expression;protein purification
作者简介:任 方(1989-),女,新疆乌鲁木齐人,硕士研究生,主要从事动物病毒病研究。*通讯作者
基金项目:国家自然科学基金项目(31160505);国家科技支撑计划项目(2013BAD12B04)
收稿日期:2014-12-02
中图分类号:S852.7
文献标识码:A
文章编号:1007-5038(2015)06-0043-05
