5-HTR1A基因rs6295多态性与父母教养行为对青少年早期抑郁的交互作用:不同易感性模型的验证*
2015-02-06王美萍张文新陈欣银
王美萍 张文新 陈欣银
(1山东师范大学心理学院,济南 250014) (2宾夕法尼亚教育研究所,美国费城 19104-6216)
1 问题提出
抑郁是个体主要和常见的心理健康问题之一,在世界范围内也是造成残疾和疾病负担的五大原因之一(Caspi et al.,2003)。它通常用来指一系列范围较广的情绪问题,根据严重程度可分为三种类型:抑郁情绪(depression mood),主要特征是悲伤、苦恼或烦躁等;抑郁综合征(depressive syndromes),除表现出消极情绪外,还伴随着退缩、注意力涣散等行为特征;抑郁障碍(depressive disorder)或者重性抑郁(major depression),除上述特征外,同时伴有明显的躯体症状,如失眠、厌食等(Compas,Ey,&Grant,1993)。常态群体中儿童青少年的抑郁主要表现为抑郁情绪和抑郁综合征,而精神病学领域主要考察的是重性抑郁或者抑郁障碍。
青少年早期是抑郁发生的关键时期。横断和追踪研究均表明,与童年期相比,青少年早期的抑郁情绪发生率迅速增加(Brendgen,Wanner,Morin,&Vitaro,2005),此后,趋于稳定并接近成人总体水平(Compas et al.,1993)。文献显示,我国中学生的抑郁综合征发生率在20%~44%之间(冯正直,2002;王卫,2000),而且青少年期的抑郁会导致该时期的个体心理失衡,并与成年后的继发抑郁显著相关(Pine,Cohen,Cohen,&Brook,1999;Lewinsohn,Solomon,Seeley,&Zeiss,2000)。因而,考察青少年期抑郁,特别是青少年早期抑郁的影响因素与发生机制是抑郁科学研究中的重要基础性课题。
随着分子遗传学的兴起与发展,探索与抑郁相关的遗传基因及其与环境因素的交互作用方式成为当前该研究领域的前沿课题之一,并获得了一些有价值的发现。例如,Caspi等人(2003)刊发于《Science》上的一篇研究报告显示,5羟色胺转运体(serotonin-transporter-linked promoter region,5-HTTLPR)基因多态性对压力性生活事件与抑郁的关系具有显著调节作用,与携带 LL纯合型基因的个体相比,那些短等位基因携带者(携带SL和SS基因型的个体)在面临压力性生活事件时,更容易出现抑郁症状。斯坦福大学的 Thompson,Parker,Hallmayer,Waugh和Gotlib (2011)近期的研究表明,催产素受体(oxytocin receptor,OXTR)基因rs2254298多态性与消极父母教养环境对青少年的抑郁具有显著交互作用,那些携带杂合型基因(AG)且成长于高消极环境下的青少年的抑郁水平显著高于其他组。国内新近的一项研究(Chen,Li,&McGue,2013)亦表明,脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)基因Val66Met多态性与压力性生活事件对青少年抑郁具有显著交互作用,那些携带Val等位基因的个体更容易受到压力性生活事件的影响而表现出较高的抑郁水平。
通览相关文献可以发现,既有关于抑郁的基因-环境交互作用研究的主要理论基础是“素质?压力模型” (diathesis-stress model)。该模型认为,当个体处于应激或高压状态时,具有某种不良遗传素质的个体更容易发生心理与行为问题。由“素质?压力模型”出发,已有研究通常采用消极环境指标(例如童年期的受虐经历、母亲的抑郁、父母离异、压力性生活事件等)来考察基因?环境交互作用(Aslund et al.,2009;Caspi et al.,2003;Beach et al.,2010;Thompson et al.,2011)。显然,这类研究所考察的问题是“携带某种‘风险’基因型的个体是否更容易受到某种消极环境的影响?”然而,一个重要的基本事实是消极成长环境的存在并非完全等同于积极成长环境的缺失。基因?环境交互作用既可能表现为某些基因型的个体比其他个体更容易受到消极成长环境的影响而出现问题或障碍,也可能表现为某些基因型的个体比其他个体更容易受到积极成长环境的影响而表现良好或优秀。
新近兴起的理论模型——“不同易感性模型”(differential susceptibility model)明确提出并证明,同一基因型能够令个体变得“更好或更坏” (Belsky&Pluess,2009;Ellis,Boyce,Belsky,Bakermans-Kranenburg,&van Ijzendoorn,2011)。因此,现有以“素质?压力模型”为理论基础的研究未能揭示基因?环境交互作用的多种可能方式,深入地揭示遗传基因与环境因素对青少年早期抑郁的作用机制,需要同时考察青少年成长的积极环境和消极环境的影响。家庭是儿童青少年成长的重要微观环境变量(Bronfenbrenner,1979),其中父母的教养行为对青少年的发展具有重要影响。根据性质父母教养行为可以分为积极教养行为与消极教养行为。既有研究表明父母支持(Hayden et al.,2010)、温情(Propper,Willoughby,Halpern,Carbone,&Cox,2007),以及敌对与消极的父母教养(England &Sim,2009;McGinn,Cukor,&Sanderson,2005)均与抑郁存在显著关联。因此,本研究将采用不同性质父母教养行为作为环境指标,对不同易感性模型的观点进行验证。
基因指标则选取 5-羟色胺 1A受体(serotonin receptor 1A,5-HTR1A)基因rs6295多态性。已有分子遗传学研究显示(Kishi et al.,2009;Wu et al.,2008),5-HTR1A基因是抑郁的重要候选基因。人类5-HTR1A基因位于第 5号染色体 q12.3区,编码5-HTR1A。5-HTR1A的激活可抑制中缝核神经元放电,导致5-羟色胺的合成与释放减少。精神药理学和神经行为研究结果表明,中枢 5-羟色胺(5-HT)功能异常与抑郁症的发病相关(薛坤喜,2010),抑郁症患者和抑郁自杀死亡者脑内的 5-HTR1A表达或活动过度(Stockmeier et al.,1998)。5-HTR1A基因有Gly22Ser (rs1799920),Ile28Val (rs1799921),C-1019G (rs6295)和 Arg219Leu (rs180004)等多个多态性位点(薛坤喜,2010;Drago,Ronchi,&Serretti,2008),其中最受研究者关注的是转录调控区的rs6295位点。该位点的多态性能够通过调节双重抑制因子核DEAF-1相关蛋白和Hes5的表达,从而影响突触前、后膜5-HTR1A的表达(Szewczyk et al.,2009;Le François,Czesak,Steubl,&Albert,2008;陈爱敏,2011)。有资料表明,rs6295基因多态性与经前焦虑障碍显著相关,C等位基因携带者的焦虑水平显著高于 GG基因型携带者(Dhingra et al.,2007)。Wu等人(2008)的研究进一步显示,重性抑郁患者中CC基因型携带者显著多于健康控制组。然而,也有研究表明(陈爱敏,2011),G等位基因可能增加抑郁的发生率。研究结果的分歧可能与上述研究均没有涉及环境因素的效应有关。根据不同易感性模型的观点,同一基因型能够令个体变得“更好或更坏”,因而,当不同的研究中被试所经历的环境水平不一样时,研究者就有可能获得截然相反的结论。鉴于此,本研究中我们将采用 5-HTR1A基因rs6295多态性为遗传指标,父母积极和消极教养行为作为环境指标,对不同易感性模型观点进行验证,即携带某种基因型的个体是否在消极成长环境下更容易患抑郁,而在积极成长环境下则表现出更低的抑郁水平。
遗传基因与环境因素对抑郁的交互作用是否存在性别差异的问题是抑郁的分子遗传学研究领域重要且亟待研究的内容。迄今仅有少数几项研究对这一问题进行了考察,而且研究结果显示不同基因与环境对抑郁的交互作用的性别差异模式不一致。例如,Sjöberg等人(2006)发现,携带5-HTTLPR短等位基因(S)的女青少年更容易受环境压力的影响而患抑郁,但该等位基因对男青少年具有保护作用。Nyman等人(2011)的研究结果则表明儿茶酚胺氧位甲基转移酶(catechol-O-methyltransferase,COMT)基因rs4680多态性与环境的交互效应仅存在于男性群体中,携带 G等位基因的男性在经历了环境压力后抑郁水平较高。然而关于5-HTR1A基因rs6295多态性与环境因素的交互作用是否存在性别差异,如果存在,其具体的差异模式是怎样的问题仍有待探讨。
综上所述,本研究拟以不同易感性模型为理论基础,以青少年早期个体为被试,考察5-HTR1A基因rs6295多态性与父母积极和消极教养行为对青少年早期抑郁的交互作用模式及其性别差异,一方面考证其交互作用模式是否符合不同易感性模型的观点,另一面揭示该交互作用模式是否具有性别一致性。
2 研究方法
2.1 样本
选取山东省济南市11所小学的5年级2715名学生及其母亲作为被试。连续追踪测评4年。其中提取基因被试1555名(筛选过程见研究程序)。本研究采用了第一个时间点和最后一个时间点的数据,剔除因被试流失而缺失的数据后共获有效被试1323名,其中男生683人,女生640人。初次测评时学生被试平均年龄为11.31岁(SD
=0.49),母亲的平均年龄为38.03±2.39。母亲受教育水平为初中及其以下者占 12.8%,高中或中专毕业者 25.7%,大专或大学本科毕业者56.10%,研究生毕业者5.40%。家庭月收入1000元以下的占3.30%,1000~3000元的占27.60%,3000~6000元的占46.80%,6000元以上的占22.20%。2.2 研究工具、仪器与材料
2.2.1 青少年抑郁量表
采用国内学者俞大维和李旭修订的由美国心理学家 Kovacs编制(Children’s Depression Inventory,CDI;Kovacs,1992)的儿童抑郁量表。该量表包括27个项目,内容涉及抑郁的一系列症状(例如,“不高兴”、“社交减少”、“感到疲倦”、“睡眠障碍”等),适用于年龄在 7~17岁之间的儿童,由青少年自我报告。采用 3 点记分——0(偶尔)、1(经常)、2(总是),以所有项目的总分代表抑郁状况,得分越高,抑郁水平越高。两次测量中,量表的克隆巴赫系数分别为0.88和0.89。
2.2.2 父母教养行为问卷
采用项目组自行编制问卷,该问卷在对中国儿童青少年研究中已得到广泛使用,并且相关研究表明该问卷具有良好的信效度(Chen,Bian,Xin,Wang,&Silbereisen,2010;Chen,Chen,Wang,&Liu,2002;Chen,Liu,&Li,2000;Chuang &Su,2009)。问卷由母亲报告,其中温情(例如:“当我的孩子感到不安或害怕时,我会给他/她安慰和理解”)、引导(例如:“当我的孩子做错什么事情时,我会耐心地和他/她交谈”)、鼓励独立(例如:“在很多事情上,我让我的孩子自己做决定”) 3个维度测查的是积极教养行为,共14个项目;拒绝(例如“如果我的孩子不来烦我,我就不会理睬她”)与高控制/惩罚(例如:“我不允许我的孩子对我发脾气”,“我认为斥责和批评会促进孩子成长”) 2个维度测查的是消极教养行为,共10个项目。验证性因素分析显示,一阶二因子模型结构拟合较好(c=1928.34,df
=251,RMSEA=0.05,CFI=0.86,TLI=0.85)。问卷采用5点记分(0? “完全不符合”,4 ? “完全符合”),平均得分越高,则该种教养行为越多。积极和消极教养行为内部一致性系数分别为0.85和0.72。2.2.3 DNA样本采集、提取、分型仪器与试剂
仪器 离心机(Eppendorf公司)、D-37520台式离心机(Thermo公司)、PCR仪(GeneAmp PCR System 9700,Applied Biosystems公司)、MassArray TM Nanodispenser (SAMSUNG 公司)、MassARRAY compact System (SEQUENOM 公司)、G384+10 Spectrochip TM (SEQUENOM 公司)、移液枪(Eppendorf公司)。
试剂 HotStarTaq DNA Polymerase (1000 U)(包括 4×250 units HotStarTaq DNA Polymerase、10×PCR Buffer、25 mM MgCl2,Qiagen 公司)、iPLEX TM Reagent Kit(包括 10×SAP Buffer、1 U/uL SAP enzyme、10×iPlex Buffer、iPlex Termination mix、iPlex enzyme,SEQUENOM 公司)、Clean Resin(SEQUENOM 公司)、dNTP Mixture (TaKaRa 公司)。
2.3 研究程序
2.3.1 问卷调查程序
以学校为单位,选取山东省济南市的 11所小学的5年级学生作为测查对象,并向所测查学生及其监护人发放参与本项目研究的同意信。在取得他们同意的前提下,以班级为单位,采用问卷法进行第一次(2008年)抑郁量表测查,共获得有效被试2715名。在与各学校领导、班主任老师协商好的时间段内,以班级为单位组织母亲到学校完成父母教养行为问卷的测查。之后连续追踪测评 3年(2009~2011年),每年测评一次。主试均由经过严格培训的心理学专业教师、研究生和大四本科生担任。
2.3.2 DNA样本采集与分型程序
在征得学生被试及其监护人同意的前提下,以班级为单位采集学生被试的唾液样本,每人采集2~5 ml,最终共获取有效样本1555个。而后将唾液样本交付上海某生物科技有限公司,由该公司专业技术人员进行DNA的提取、纯化与分型。DNA分型利用美国Sequenom公司的MassARRAY系统完成。5-HTR1A基因引物为F:ACGTTGGATGTTGTTTTCTTCATTGGGCCG,R:ACGTTGGATGGCTTGACATCATTGGCTGAC。PCR 反应条件:94°C 15min;94°C 20s,56°C 30s,72°C 1min,共 45 个循环;最终 72°C 3min。PCR 扩增后,剩余的 dNTP将被去磷酸消化掉,反应体系包括1.53 μL水、0.17 μL SAP缓冲液、0.3单位碱性磷酸酶(Sequenom)。该反应在37°C进行40 min,然后85°C 5min使酶失活。碱性磷酸酶处理后,针对SNP的单碱基延伸引物在下列反应体系中进行:0.755 μL水、0.2μL 10×iPLEX 缓冲液、0.2μL 终止混合物、0.041μL iPLEX酶(Sequenom),0.804 μL 10uM的延伸引物。单碱基延伸反应在下列条件下进行:94°C 30s;94°C 5s,52°C 5s,80°C 5s 5 个循环,共 40 个循环;最后72°C 3min。在终止反应物中加入6 mg阳离子交换树脂(Sequenom)脱盐,混合后加入25 μL水悬浮。使用MassARRAY Nanodispenser (Sequenom)将最终的分型产物点样到一块 384孔的 spectroCHIP(Sequenom)上,并用基质辅助激光解吸电离飞行时间质谱进行分析。最终结果由MassARRAY RT软件系统(版本号 3.0.0.4)实时读取,并由 MassARRAY Typer软件系统(版本号3.4)完成基因分型分析。
2.4 数据处理与分析
采用 Foxbase 2.0进行数据录入,应用 SPSS 19.0进行数据处理与分析。主要统计方法为分层回归分析。基于横断数据在解释变量间关系方面的局限性,本研究的父母教养采用的是第一个测量时间点(2008年)的数据,抑郁采用的是最后一个时间点(2011年)的数据,同时控制了基线水平(2008年)的抑郁。另外,由于rs6295位点GG型基因携带者较少(CC=719人,约54%;CG=497人,约37.67%;GG=107人,约8.1%),所以参照同类研究的常规做法(Barry,Kochanska,&Philibert,2008;Gu et al.,2013;Lee et al.,2005),在后续的分析中我们将CG与GG基因型携带者进行了合并,统称为G等位基因携带者。此外,初步的统计分析显示,提取基因与未提取基因被试的抑郁得分(2008:t
=1.15,df
=2713;2011:t
=0.45,df
=1876;ps
>0.05)与父母教养得分(积极教养:t
=0.32,df
=2192;消极教养:t
=1.72,df
=2192;p
s >0.05)均无显著差异。此外,为了便于对 rs6295多态性与父母教养行为的交互作用进行简单效应分析,我们对父母教养行为进行了标准化处理,高于平均分 1个标准差的为高分组,低于平均分1个标准差的为低分组,各组的人数分布具体如下:高积极教养组249人(CC型:137人,CG/GG型:112人);低积极教养组253人(CC型:143人,CG/GG型:110人);高消极教养组237人(CC型:120人,CG/GG型117人);低分组223人(CC型:122人,CG/GG型101人)。3 研究结果
3.1 5-HTR1A基因rs6295位点的基因型分布
rs6295位点基因型分布(见表1)不存在显著的性别差异(χ=0.76,df
=1,p
>0.05);其观测值与期望值吻合良好(χ=2.58,df
=1,p
>0.05),符合Hardy-Weinberg平衡定律。
表1 rs6295位点基因型在男女青少年中的分布
3.2 变量的描述统计量与相关分析结果
如表2所示,男女青少年的父母积极教养行为和消极教养行为均存在显著差异,具体表现为父母对男青少年的积极教养行为显著少于对女青少年的,但消极教养行为显著多于对女青少年的。第一个测量时间点(5年级)的抑郁得分也存在显著性别差异,男青少年的抑郁得分高于女青少年(2008:M
=5.55,SD
=6.12,M
=4.30,SD
=5.30),在第二个时间点上男青少年抑郁得分也表现出高于女青少年的趋势,但差异并不显著(2011:M
=7.65,SD
=7.21,M
=6.94,SD
=6.39)。Rs6295 多态性与积极和消极教养行为相关不显著,说明可以排除基因-环境相关的可能性;rs6295多态性与性别、抑郁相关亦不显著。积极教养行为与消极教养行为呈显著负相关,因而为避免多元共线性的问题,在后续的分析中分别对积极和消极教养行为建构模型进行相应的统计分析。积极教养行为与抑郁呈显著负相关,消极教养行为则与抑郁呈显著正相关。两次抑郁得分之间存在显著正相关,因此在考察rs6295多态性与父母教养行为,以及二者的交互作用对抑郁(2011)的预测作用时,我们对基线水平的抑郁(2008)进行了控制。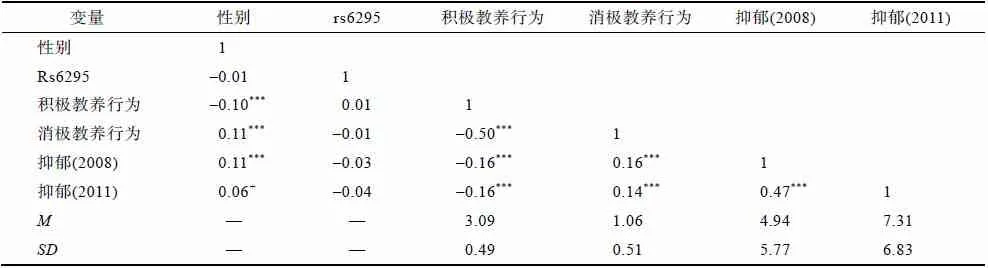
表2 rs6295多态性、性别、父母教养行为与抑郁的描述统计量及相关分析结果
3.3 5-HTR1A基因rs6295多态性与父母教养行为对男女青少年早期抑郁的影响
以 2011年时间点的抑郁为因变量,基线水平的抑郁(2008)为控制变量,rs6295多态性、积极教养行为或者消极教养行为,以及二因素的交互项为预测变量,分别在男女青少年群体中进行分层回归分析,结果显示(见表3),rs6295多态性对男女青少年早期抑郁均无显著主效应;父母积极教养行为则能够显著负向预测男女青少年的抑郁,消极教养行为能够显著正向预测男青少年抑郁,但对女青少年抑郁无显著预测作用;rs6295多态性与父母积极教养行为对女青少年的抑郁具有显著交互作用,但与父母消极教养行为的交互作用不显著,此外,rs6295多态性与父母积极或消极教养行为对男青少年的抑郁均无显著交互作用。

表3 抑郁对男女青少年的5-HTR1A基因rs6295多态性与父母教养行为的分层回归分析
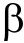
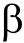
鉴于 rs6295多态性与父母消极教养行为的检验结果在校正前呈边缘显著,而且本研究中女青少年消极教养行为的较低得分(见表2)可能会限制其与rs6295多态性交互作用的效应,我们进一步探查了二者的交互作用图。由图1和2可以看出rs6295多态性与父母消极教养行为对女青少年抑郁亦呈交互作用趋势,即二者的效应并非完全相互独立。

图1 rs6295多态性与父母教养行为对女青少年早期抑郁的交互作用

图2 rs6295多态性与父母教养行为对女青少年早期抑郁的交互作用
4 讨论
本研究结果显示5-HTR1A基因rs6295多态性与父母教养行为对青少年早期抑郁的交互作用显著,并且该交互作用存在性别差异。具体表现为,在女青少年群体中,rs6295多态性与父母积极教养行为对抑郁存在显著交互作用,然而,在男青少年群体中,rs6295多态性与父母教养行为对抑郁无任何显著交互作用。
对女青少年群体中基因-环境交互效应的进一步分析显示,当父母积极教养行为水平较低时,携带不同基因型的女青少年的抑郁水平无显著差异,然而当父母积极教养行为水平较高时,CC型基因携带者的抑郁水平显著低于G等位基因携带者。另一方向的分析同时表明,父母的积极教养行为可以显著预测CC基因型携带者的抑郁,但对G等位基因携带者的抑郁无显著预测作用。上述研究结果表明,rs6295位点CC型基因可能是对外界环境更为敏感的“可塑基因” (plasticity gene),而非风险基因(vulnerability gene)。该研究结果得到了精神病药理学领域研究发现的支持。例如,有关rs6295多态性与抗抑郁药物反应的研究显示,CC基因型携带者更容易受到药物的影响而表现出更好的治疗效果(Hong,Chen,Yu,&Tsai,2006;Yu et al.,2006)。另外,由于在常态群体中抑郁的得分通常呈正偏态分布,本研究中抑郁水平也呈中等程度的正偏态分布,所以我们也尝试了对抑郁得分进行平方根转换后重新进行统计分析,结果无实质性改变,而且本研究所呈现的是经过Benjamini–Hochberg程序(1995)校正后的结果。这表明了上述研究结果的稳定性。
令人遗憾的是,目前尚未有资料能够揭示与阐明CC基因型对环境影响更为敏感的内在机理,但相关文献显示,这可能有两个方面的原因:其一,与C等位基因和G等位基因的不同神经生物功能有关。譬如,Pernhorst等人(2013)的研究表明,CC型颞叶癫痫病人携带者的海马区域的 5-HTR1A的mRNA的表达显著高于 GG型基因携带者;Lindstedt等人(2012)的研究则进一步显示,G等位基因能够削弱5-羟色胺神经元信号,降低突触后受体的表达,导致个体对热疼痛刺激的反应不敏感。其二,可能与rs6295基因多态性和附近其他基因多态性间的相互作用有关(Hong et al.,2006)。例如,有动物模型研究显示(Li,Wichems,Heils,Lesch,&Murphy,2000),5-HTT基因敲除鼠的5-HTR1A基因mRNA的表达和浓度均显著下降。
本研究所揭示的上述基因-环境交互作用是一种不同于“素质?压力模型”观点的新的交互作用形式,这表明了仅着眼于考察消极环境因素与基因交互作用的“素质?压力模型”的不足。此外,通过对比图1的左右两图我们可以发现,携带不同基因型的青少年在低积极教养行为下的抑郁差异模式,与在高消极教养行为条件下的抑郁差异模式不完全一致,他们在高积极教养行为下的抑郁差异模式与在低消极教养行为下的也不完全一致。简言之,消极教养行为的缺失并非等同于积极教养行为的存在,反之亦然。这进一步表明了在探查抑郁的发生机制时,同时考察积极与消极环境因素的必要性。尽管如此,本研究结果只部分支持了不同易感性模型的观点(Belsky et al.,2009;Ellis et al.,2011),即只验证了其关于携带某种基因型的个体在积极成长环境下会表现出更低的抑郁水平的假设,而没有支持其携带该基因型个体在消极成长环境下也更容易患抑郁的观点。该结果一方面可能说明rs6295多态性与环境的交互作用模式既不符合素质?压力模型,也不符合不同易感性模型,而是符合新近提出的“优势敏感性” (vantage sensitivity)作用模式(Pluess &Belsky,2013),即CC型基因携带者更容易受到积极环境的影响,而对消极环境不敏感。另一种可能是 rs6295多态性与环境的交互作用模式实际上应该符合不同易感性模型,但是由于本研究的被试均为正常群体青少年,所报告的消极教养得分相对较低(M
=1.11,SD
=0.51 ;M
=0.99,SD
=0.51),其变异范围不足以揭示出消极教养与 rs6295多态性的交互作用。然而现有资料显示,rs6295多态性能够调节负性生活事件对重性抑郁的影响(Zhang et al.,2009),所以第一种推理应该不成立。此外,本研究中rs6295多态性与父母消极教养行为对女青少年抑郁的交互作用图(图1与图2)显示,二者的效应并非相互独立。因此,我们的第二种推理更为合理,未来研究可增加临床抑郁患者样本,对该结果继续进行考证。另外,本研究中所发现的基因-环境交互效应较小(仅为 1%)。这与既有关于抑郁的遗传与环境交互作用机制的研究发现是一致的。譬如,Klauke等人(2011)在考察 5-羟色胺转运体基因与儿童期创伤对抑郁的影响时发现,二者的交互效应的解释率仅为1.7%。Beach等人(2010)在考察儿童期虐待与 MAOA基因对抑郁的交互作用时,也仅发现了1.2%的效应量。Plomin也曾经指出,与动物研究相比,要鉴别人类行为的基因-环境交互作用更加困难(Plomin,DeFries,McClearn &McGuffin,2001/2008),人类的绝大多数表型并不像苯丙酮尿症等单基因遗传疾病那样有着清晰的遗传机制,其单基因位点的解释率往往不足1% (Plomin,Kennedy,&Craig,2006)。这或许也正是本研究发现 rs6295多态性对青少年抑郁无任何显著主效应的原因所在。神经递质之间的功能关系十分复杂,一种递质功能紊乱可能引起另外一种或几种递质的功能失衡,从而导致一定的病理生理现象。因而,未来研究有必要采用多基因-环境设计进一步探查 rs6295多态性对青少年早期抑郁的影响。本研究所揭示的基因-环境交互作用的性别差异模式,与既有抗抑郁药物的研究结果也相一致。例如,Yu等人(2006)的研究发现,携带CC基因型的个体比携带 G等位基因的个体对抗抑郁药物氟西汀(fluoxetine)的反应更好,并且该关联仅存在于女性群体中。尽管目前对5-HTR1A基因rs6295多态性与父母教养行为交互作用性别差异的内在机制尚不清楚,但根据曹衍淼等人(2013)的文献分析结果,这可能与男女对不同类型环境的敏感性的差异有关。譬如,携带5-HTTLPR S等位基因的女性更容易受到父母婚姻质量、亲子关系质量、伤害性家庭冲突等的影响,而携带5-HTTLPR L等位基因的男性更容易受到地区贫困水平、公共居住环境等宏观社会环境水平的影响。本研究所选择的父母教养行为,以及 Yu等人(2006)的药物治疗均属于微系统或者近端环境因素,因此,未来研究中有必要选择其他水平的环境指标,进一步探查 5-HTR1A基因 rs6295多态性与环境因素对抑郁交互作用的性别差异模式。
此外,本研究结果显示,在第一个时间点上(5年级)男青少年的抑郁水平显著高于女青少年,然而在第二个时间点上(8年级)个体抑郁水平的性别差异并不显著。王美萍和张文新(2010)以初中生为被试的研究则发现,初一与初二(7、8年级)青少年的抑郁水平无显著性别差异,但初三(9年级)女青少年的抑郁得分显著高于男青少年。由此可以推断,从童年期至青少年期,个体抑郁水平的性别差异大致遵循着这样一种发展模式:小学时期男生抑郁得分显著高于女生,进入初中后女生抑郁水平逐渐增加,性别差异逐渐缩小,至初三时女生抑郁得分显著高于男生。即小学时期与中学时期抑郁的性别差异表现出不同的模式。国外有关抑郁的元分析的研究结果显示(Twenge &Nolen-Hoeksema,2002),8-12岁期间,男生的抑郁得分高于女生,但自13岁始,女生抑郁得分反而显著高于男生。这与上述假设基本一致,也从另一个侧面印证了本研究结果的可靠性。
5 研究结论
5-HTR1A基因rs6295多态性与父母教养行为的交互作用是否符合不同易感性模型的观点?其交互作用模式是否具有性别一致性?本研究对此进行了探索并发现,携带CC基因型的青少年在积极教养环境下表现出更低的抑郁水平,且该交互作用仅存在于女青少年群体中。这为不同易感性模型的观点提供了部分实证支持,同时也表明了rs6295多态性与父母教养行为的交互作用模式存在性别差异。
Aslund,C.,Leppert,J.,Comasco,E.,Nordquist,N.,Oreland,L.,&Nilsson,K.W.(2009).Impact of the interaction between the 5-HTTLPR polymorphism and maltreatment on adolescent depression.A population-based study.Behavior Genetics,39
(5),524–531.Barry,R.A.,Kochanska,G.,&Philibert,R.A.(2008).G× E interaction in the organization of attachment:Mothers’responsiveness as a moderator of children’s genotypes.Journal of Child Psychology and Psychiatry,49
(12),1313–1320.Beach,S.R.H.,Brody,G.H.,Gunter,T.D.,Packer,H.,Wernett,P.,&Philibert,R.A.(2010).Child maltreatment moderates the association of MAOA with symptoms of depression and antisocial personality disorder.Journal of Family Psychology,24
(1),12–20.Belsky,J.,&Pluess,M.(2009).Beyond diathesis stress:Differential susceptibility to environmental influences.Psychological Bulletin,135
(6),885–908.Benjamini,Y.,&Hochberg,Y.(1995).Controlling the false discovery rate:A practical and powerful approach to multiple testing.Journal of the Royal Statistical Society.Series B (Methodological),57
(1),289–300.Brendgen,M.,Wanner,B.,Morin,A.J.S.,&Vitaro,F.(2005).Relations with parents and with peers,temperament,and trajectories of depressed mood during early adolescence.Journal of Abnormal Child Psychology,33
(5),579–594.Bronfenbrenner,U.(1979).Contexts of child rearing:Problems and prospects.American Psychologist,34
(10),844–850.Cao,Y.M.,Wang,M.P.,Cao,C.,Chen,G.H.,&Zhang,W.X.(2013).Gender differences in the genetic underpinning of depression.Advances in Psychological Science,21
(9),1605–1616.[曹衍淼,王美萍,曹丛,陈光辉,张文新.(2013).抑郁遗传基础的性别差异.心理科学进展,21
(9),1605–1616.]Chen,J.,Li,X.Y.,&McGue,M.(2013).The interacting effect of the BDNF Val66Met polymorphism and stressful life events on adolescent depression is not an artifact of gene–environment correlation:Evidence from a longitudinal twin study.Journal of Child Psychology and Psychiatry,54
(10),1066–1073.Caspi,A.,Sugden K.,Moffitt T.E.,Taylor,A.,Craig,I.W.,Harrington,H.,...Poulton,R.(2003).Influence of life stress on depression:Moderation by a polymorphism in the 5-HTT gene.Science,301
(5631),386–389.Chen,A.M.(2011).Relationship between 5-HTR1AC(-1019)G and G-protein b3 subunit C825T polymorphisms and post-stroke depression
(Unpublished master’s thesis).Southern Medical University,Guangdong.[陈爱敏.(2011).5-羟色胺1A受体、G蛋白β3亚基基因多态性与卒中后抑郁的相关性研究
(硕士学位论文).南方医科大学,广东.]Chen,X.,Bian,Y.,Xin,T.,Wang,L.,&Silbereisen,R.K.(2010).Perceived social change and childrearing attitudes in China.European Psychologist,15
(4),260–270.Chen,X.Y.,Chen,H.C.,Wang,L.,&Liu,M.W.(2002).Noncompliance and child-rearing attitudes as predictors of aggressive behaviour:A longitudinal study in Chinese children.International Journal of Behavioral Development,26
(3),225–233.Chen,X.Y.,Liu,M.W.,&Li,D.(2000).Parental warmth,control,and indulgence and their relations to adjustment in Chinese children:A longitudinal study.Journal of Family Psychology,14
(3),401–419.Chuang,S.S.,&Su,Y.(2009).Do we see eye to eye? Chinese mothers’ and fathers’ parenting beliefs and values for toddlers in Canada and China.Journal of Family Psychology,23
(3),331–341.Compas,B.E.,Ey,S.,&Grant,K.E.(1993).Taxonomy,assessment,and diagnosis of depression during adolescence.Psychologieal Bulletin,114
(20),323–344.Drago,A.,Ronchi,D.D.,&Serretti,A.(2008).5-HT1A gene variants and psychiatric disorders:A review of current literature and selection of SNPs for future studies.International Journal of Neuropsychopharmacology,11
(5),701–721.Dhingra,V.,Magnay,J.L.,O'Brien,P.M.S.,Chapman,G.,Fryer,A.A.,&Ismail,K.M.(2007).Serotonin receptor 1A C (-1019)G polymorphism associated with premenstrual dysphoric disorder.Obstetrics &Gynecology,110
(4),788–792.Ellis,B.J.,Boyce,W.T.,Belsky,J.,Bakermans-Kranenburg,M.J.,&van Ijzendoorn,M.H.(2011).Differential susceptibility to the environment:An evolutionaryneurodevelopmental theory.Development and Psychopathology,23
(1),7–28.England,M.J.,&Sim,L.J.(Eds.).(2009).Depression in parents,parenting,and children:Opportunities to improve identification,treatment,and prevention
.Washington:National Academies Press.Feng,Z.Z.(2002).A study on social information processing of middle school students with depressive symptoms
(Unpublished doctoral dissertation).The Xi’nan Normal University,Chongqing.[冯正直.(2002).中学生抑郁症状的社会信息加工方式研究
(博士学位论文).西南师范大学,重庆.]Gu,H.,Liu,C.,Chen,M.,Zhang,Q.,Zhai,J.,Wang,K.,...Chen,C.(2013).The combined effects of the 5-HTTLPR and HTR1A rs6295 polymorphisms modulate decision making in schizophrenia patients.Genes,Brain and Behavior,12
(1),133–139.Hayden,E.P,Klein,D.N.,Dougherty,L.R.,Olino,T.M.,Laptook,R.S.,Dyson,M.W.,...Singsh,S.M.(2010).The dopamine D2 receptor gene and depressive and anxious symptoms in childhood:Associations and evidence for gene-environment correlation and gene-environment interaction.Psychiatric Genetics,20
(6),304–310.Hong,C.J.,Chen,T.J.,Yu,Y.W.,&Tsai,S.J.(2006).Response to fluoxetine and serotonin 1A receptor (C-1019G)polymorphism in Taiwan Chinese major depressive disorder.The Pharmacogenomics Journal,6
(1),27–33.Kishi,T.,Tsunoka,T.,Ikeda,M.,Kawashima,K.,Okochi,T.,Kitajima,T.,… Iwata,N.(2009).Serotonin 1A receptor gene and major depressive disorder:An association study and meta-analysis.Journal of Human Genetics,54
(11),629–633.Klauke,B.,Deckert,J.,Reif,A.,Pauli,P.,Zwanzger,P.,Baumann,C.,…Domschke,K.(2011).Serotonin transporter gene and childhood trauma—A G×E effect on anxiety sensitivSerotonin transporter gene and childhood trauma—A G×E effect on anxiety sensitivity.Depressed and Anxiety,28
(12),1048–1057.Kovacs,M.(1992).Children depression inventory (CDI)manual
.Toronto:Multi-Health Systems Inc.Le François,B.,Czesak,M.,Steubl,D.,&Albert,P.R.(2008).Transcriptional regulation at a HTR1A polymorphism associated with mental illness.Neuropharmacology,55
(6),977–985.Lee,S.G.,Joo,Y.,Kim,B.,Chung,S.,Kim,H.L.,Lee,I.,…Song,K.(2005).Association of Ala72Ser polymorphism with COMT enzyme activity and the risk of schizophrenia in Koreans.Human Genetics,116
(4),319–328.Lewinsohn,P.M.,Solomon,A.,Seeley,J.R.,&Zeiss,A.(2000).Clinical implications of “subthreshold” depressive symptoms.Journal of Abnormal Psychology,109
(2),345–351.Li,Q.,Wichems,C.,Heils,A.,Lesch,K.P.,&Murphy,D.L.(2000).Reduction in the density and expression,but not G-protein coupling,of serotonin receptors (5-HT1A) in 5-HT transporter knock-out mice:Gender and brain region differences.The Journal of Neuroscience,20
(21),7888–7895.Li,D.P.,Zhang,W.,Li,D.L.,Wang,Y.H.,&Zhen,S.J.(2012).The effects of parenting styles and temperament on adolescent aggression:Examining unique,differential,and mediation effects.Acta Psychologica Sinica,44
(2),211–225.[李董平,张卫,李丹黎,王艳辉,甄霜菊.(2012).教养方式、气质对青少年攻击的影响:独特、差别与中介效应.心理学报,44
(2),211–225.]Lindstedt,F.,Karshikoff,B.,Schalling,M.,Olgart Höglund,C.,Ingvar,M.,Lekander,M.,&Kosek,E.(2013).Serotonin-1A receptor polymorphism (rs6295) associated with thermal pain perception.Plos One,7
(8),e43221.McGinn,L.K.,Cukor,D.,&Sanderson,W.C.(2005).The relationship between parenting style,cognitive style,and anxiety and depression:Does increased early adversity influence symptom severity through the mediating role of cognitive style?Cognitive Therapy and Research,29
(2),219–242.Nyman,E.S.,Sulkava,S.,Soronen,P.,Miettunen,J.,Loukola,A.,Leppä,V.,...Paunio,T.(2011).Interaction of early environment,gender and genes of monoamine neurotransmission in the aetiology of depression in a large population-based Finnish birth cohort.British Medical Journal Open,1
(1),1–13.Pernhorst,K.,van Loo,K.M.,von Lehe,M.,Priebe,L.,Cichon,S.,Herms,S.,...Becker,A.J.(2013).Rs6295 promoter variants of the serotonin type 1A receptor are differentially activated by c-Jun in vitro and correlate to transcript levels in human epileptic brain tissue.Brain Research,1499
,136–144.Pine,D.S.,Cohen,E.,Cohen,P.,&Brook,J.(1999).Adolescent depressive symptoms as predictors of adult depression:Moodiness or mood disorder?American Journal of Psychiatry,156
(1),133–135.Plomin,R.,DeFries,J.C.,McClearn,G.E.,&McGuffin,P.(2008).Behavioral genetics
(Wen,N.,Wang,X.H.,Yang,Y.P.,&Liu,X.L.Trans).
Shanghai,China:Huadong Normal University Press.(Original work published 2001).[Plomin,R.,DeFries,J.C.,McClearn,G.E.,&McGuffin,P.(2008).行为遗传学
(温暖,王小惠,杨彦平,刘晓陵译).上海:华东师范大学出版社.]Plomin,R.,Kennedy,J.K.J.,&Craig,I.W.(2006).The quest for quantitative trait loci associated with intelligence.Intelligence,34
(6),513–526.Pluess,M.,&Belsky,J.(2013).Vantage sensitivity:Individual differences in response to positive experiences.Psychological Bulletin,139
(4),901–916.Propper,C.,Willoughby,M.,Halpern,C.T.,Carbone,M.A.,&Cox,M.(2007).Parenting quality,DRD4,and the prediction of externalizing and internalizing behaviors in early childhood.Developmental Psychobiology,49
(6),619–632.Sjöberg,R.L.,Nilsson,K.W.,Nordquist,N.,Öhrvik,J.,Leppert,J.,Lindström,L.,&Oreland,L.(2006).Development of depression:Sex and the interaction between environment and a promoter polymorphism of the serotonin transporter gene.International Journal of Neuropsychopharmacology,9
(4),443–449.Stockmeier,C.A.,Shapiro,L.A.,Dilley,G.E.,Kolli,T.N.,Friedman,L.,&Rajkowska,G.(1998).Increase in serotonin-1A autoreceptors in the midbrain of suicide victims with major depression–postmortem evidence for decreased serotonin activity.The Journal of Neuroscience,18
(18),7394–7401.Szewczyk,B.,Albert,P.R.,Burns,A.M.,Czesak,M.,Overholser,J.C.,Jurjus,G.J.,...Austin,M.C.(2009).Gender-specific decrease in NUDR and 5-HT1A receptor proteins in the prefrontal cortex of subjects with major depressive disorder.The International Journal of Neuropsychopharmacology,12
(2),155–168.Thompson,R.J.,Parker,K.J.,Hallmayer,J.F.,Waugh,C.E.,&Gotlib,I.H.(2011).Oxytocin receptor gene polymorphism(rs2254298) interacts with familial risk for psychopathology to predict symptoms of depression and anxiety in adolescent girls.Psychoneuroendocrinology,36
(1),144–147.Twenge,J.M.,&Nolen-Hoeksema,S.(2002).Age,gender,race,socioeconomic status,and birth cohort difference on the children's depression inventory:A meta-analysis.Journal of Abnormal Psychology,111
(4),578–588.Wang,M.P.,&Zhang,W.X.(2010).The Association between rs1824024 polymorphism in the CHRM2 gene and early adolescents’ depression.Acta Psychologica Sinica,42
(8),853–861.[王美萍,张文新.(2010).CHRM2基因rs1824024多态性与青少年早期抑郁的关系.心理学报,42
(8),853–861.]Wang,W.(2000).The Prevention of depression in children and adolescents:An introduction to the resourceful adolescent program.Psychological Science,23
(4),506–507.[王卫.(2000).青少年抑郁的预防:青少年应变力辅导计划简介.心理科学,23
(4),506–507.]Wu,Y.,Xu,Y.,Sun,Y.,Wang,Y.F.,Li,X.,Lang,X.E.,...Zhang,K.R.(2008).Association between the serotonin 1A receptor C (-1019) G polymorphism and major depressive disorder in the northern Han ethnic group in China.Chinese Medical Journal (English Edition),121
(10),874–876.Xue,K.X.(2010).Association study between the polymorphism of 5-Hydroxytryptamine related genes and unipolar depression in a Chinese Han population
(Unpublished master’s thesis).The Shantou University.[薛坤喜.(2010).中国汉族人群5-羟色胺相关基因多态性与单相抑郁症的关联研究研究
(硕士学位论文).汕头大学.]Yu,Y.W.Y.,Tsai,S.J.,Liou,Y.J.,Hong,C.J.,&Chen,T.J.(2006).Association study of two serotonin 1A receptor gene polymorphisms and fluoxetine treatment response in Chinese major depressive disorders.European Neuropsychopharmacology,16
(7),498–503.Zhang,K.R.,Xu,Q.,Xu,Y.,Yang,H.,Luo,J.X.,Sun,Y.,...Shen,Y.(2009).The combined effects of the 5-HTTLPR and 5-HTR1A genes modulates the relationship between negative life events and major depressive disorder in a Chinese population.Journal of Affective Disorders,114
(3),224–231.