MAOA基因rs6323多态性与同伴关系对男青少年早期抑郁的影响*
2015-02-02王美萍纪林芹张文新
王美萍 纪林芹 张文新
(山东师范大学心理学院,济南 250014)
1 问题提出
人类MAOA
基因位于X染色体p11.23-11.4区,编码单胺氧化酶 A。单胺氧化酶包括 MAOA和MAOB两种亚型,其中MAOA与单胺类神经递质,尤其是 5-羟色胺(5-hydroxytryptamine,5-HT)的亲和力高于MAOB,是5-HT的主要代谢酶。应用单胺氧化酶抑制剂(monoamine oxidase inhibitor,MAOI)能够明显提高中枢5-HT含量。精神药理学和神经行为研究结果则进一步表明,中枢 5-HT功能异常与抑郁症的发病相关(薛坤喜,2010)。因此,MAOA
基因成为抑郁研究的重要候选基因之一。现有资料表明,MAOA
基因与环境因素对个体抑郁存在显著交互作用。譬如,Beach等(2010)的研究显示,携带高活性MAOA-uVNTR (
variable number tandem repeat polymorphism)等位基因,且儿童期遭受过虐待的个体更容易抑郁。Nikulina,Widom和Brzustowicz (2012)的研究表明,那些经历过童年期虐待并携带MAOA-uVNTR
高活性等位基因的女性,抑郁得分较高。Cicchetti等(2007)的研究也发现MAOA-uVNTR
等位基因与抑郁的关系受到个体受虐经历的调节。然而,正如已有关于MAOA
基因与抑郁关系的综述(曹丛,陈光辉,王美萍,曹衍淼,张文新,2014)所提及的那样,既有相关研究环境指标的选取较为单一,主要关注的是受虐经历、压力性事件(stressful events)的影响,而忽视了对其他因素,譬如同伴关系的考察。作为影响个体发展的微系统,同伴关系对个体的社会适应具有直接和深远的影响(Mendle,Harden,Brooks-Gunn,&Graber,2012;Wentzel,2005;周宗奎,赵冬梅,孙晓军,定险峰,2006)。进入青少年早期后,个体与同伴共度的时间显著增加,对同伴文化的遵从日益增强(Rubin,Bukowski,&Parker,2006)。有资料显示(LaFontana &Cillessen,2010),早期青少年对同伴地位的重视程度达到高峰,因而考察同伴关系对青少年期早期个体抑郁的影响显得尤为必要。同伴接纳与同伴拒绝是同伴关系的两个常用测量指标。同伴接纳指团体对某一个体的喜欢程度,代表了同伴关系的积极维度,同伴拒绝则指团体对某一个体的不喜欢程度,代表了同伴关系的消极维度(张静,田录梅,张文新,2013)。既有研究显示(Nolan,Flynn,&Garber,2003;Prinstein &Aikins,2004;Stewart et al.,1999;Zimmer–Gembeck,Hunter,&Pronk,2007),同伴接纳和同伴拒绝均可以显著预测青少年期,包括青少年早期抑郁。根据新近兴起的关于基因‒环境交互作用形式的“不同易感性模型” (differential susceptibility model,Belsky&Pluess,2009),同一基因型不仅能够使某些个体比其他个体更容易受到消极环境的影响而出现问题或障碍,也能够令这些个体比其他个体更容易受到积极环境的影响而表现良好或优秀,因而要深入地揭示基因与环境对青少年早期抑郁的作用机制,有必要同时考察青少年成长的积极与消极环境的影响。鉴于此,本研究目的之一是以同伴接纳和拒绝为环境指标,探查同伴关系与MAOA
基因对青少年早期抑郁的交互作用机制。此外,现有相关研究主要考察的是位于启动子区上游的可变串联重复序列(variable number tandem repeat polymorphism,uVNTR)与抑郁的关系。MAOA-uVNTR
这一位点通常包含2R (2 repeat,通常缩写为2R,表示2次重复序列)、3R、3.5R、4R、5R和6R几种等位基因。然而,在有关这些等位基因的活性的界定上,已有研究标准并不一致。例如,有的研究(Sabol,Hu,&Hamer,1998)认为3.5R和4R的转录活性是其他等位基因的 2.4~9.6倍,因此这二者属于高活性等位基因,其他的则为低活性等位基因;有的研究(Cicchetti et al.,2007;Voltas,Aparicio,Arija,&Canals,2015)将重复序列次数大于3.5以上的等位基因视为高活性等位基因,而2R和3R为低活性等位基因;有的研究(Xie et al.,2014)则认为2R和5R等位基因的活性不确定,所以统计分析时将其剔除了。这一方面直接削弱了相关研究的可比性,另一方面可能是导致既有关于MAOA
基因活性与抑郁关系的研究结论存在分歧的原因之一。鉴于此,本研究并没有采纳MAOA-uVNTR
这一多态性位点作为候选基因指标,而是选择了另外一个在亚洲人群中也较为常见的多态性位点:位于外显子8的 rs6323(T941G)。该位点的 T/G颠换可改变MAOA活性的表达,突变后的 G等位基因致使MAOA的活性升高,相对于T等位基因,G等位基因能够提高MAOA活性多达75% (Jansson et al.,2005)。有资料显示,rs6323多态性与男性精神分裂症患者的情感失调(Kim et al.,2014)、个体对抗抑郁药的反应(Xu et al.,2011)等相关。然而,迄今尚未见到有关 rs6323多态性与环境因素对抑郁的交互作用的研究报道。因此,本研究拟采用rs6323多态性作为候选基因指标,并考察其与同伴接纳和拒绝对青少年早期抑郁的交互效应。需要说明的是,鉴于在女性群体中,MAOA
基因呈现出X染色体的随机失活(Benjamin,van Bakel,&Craig,2000),MAOA
基因在女性中的作用模式可能存在着极大的不确定性,所以为了确保研究结果的科学性和可靠性,参照诸多同类研究(Caspi et al.,2002;Marmorstein&Hart,2010)的做法,本研究只选取了男青少年作为被试。由于男性仅携带一条X染色体,所以男性在rs6323位点仅表现为T等位基因和G等位基因两种多态性,而非TT、TG和GG三种基因型。就检验基因‒环境交互作用的方法而言,目前常用的方法是传统的回归分析,如果发现存在基因‒环境交互效应,则进一步进行分组回归或者简单斜率检验以探明交互作用的具体表现形式。若结果显示携带某基因型的个体更容易受消极环境条件的影响而表现出更多的心理或行为问题,在积极环境条件下,不同基因型个体的心理或行为表现无显著差异,则表明交互作用形式符合素质‒压力模型,若结果显示那些在消极环境条件下表现出更多问题的个体,在积极环境条件下则会表现出更少的问题或者发展更为良好,则说明符合不同易感性模型(Belsky et al.,2009)。然而问题是,当所测量的环境指标的变异或者取值范围有限时,采用传统方法所发现的素质‒压力交互作用模式可能实际上是属于不同易感性交互作用模型的一部分,所以为了更科学地判断所考察的基因‒环境交互作用形式,近期有研究者颇具创意地提出了检验素质‒压力模型与不同易感性模型的新方法,其中较受推崇的是“显著性区域法” (Roisman et al.,2012)。显著性区域(region of significance,RoS)指的是自变量显著预测因变量时调节变量的取值范围(Roisman et al.,2012)。与简单斜率法相比,显著性区域分析方法考察的是自变量与因变量关联显著时调节变量近乎所有的取值[‒2 ~ +2],而不是只分析了两个点值(M
±1SD
),其优势显而易见。当某基因与因变量在环境变量的低分端区域(消极环境),而不是高分端区域(消极环境缺失或者积极环境)显著关联时,则该基因‒环境交互作用支持素质—压力模型,如果在两端区域均显著关联,则支持不同易感性模型。为了增强研究结果的可靠性和稳定性,本研究将首先运用传统的回归分析法,初步探查MAOA
基因与同伴关系对抑郁的交互作用情况,而后再采用“显著性区域法”对研究结果进行验证,若交互作用依然显著,则进一步探明其显著的区域范围。综上所述,本研究拟采用MAOA
基因 rs6323多态性为基因指标,同伴接纳与同伴拒绝为环境指标,综合运用传统的回归分析和新近兴起的“显著性区域”法,考察基因与环境对青少年早期抑郁的交互作用及其机制。2 研究方法
2.1 样本
选取山东省济南市11所中学初一的1555名学生作为被试,连续追踪测评3年。本研究采用了第一个时间点和最后一个时间点的数据,剔除因被试流失或未提供基因信息而缺失的数据后共获有效被试1323名,其中男生683人,女生640人。如前所述(见引言部分),本研究只选取了 683名男生作为被试。初次测评时男生被试平均年龄为13.35岁(SD
=0.51)。母亲和父亲平均年龄分别为40.12±2.35岁和42.04±3.04岁。母亲/父亲受教育水平为初中及其以下者占 13.30%/8.90%,高中或中专毕业者24.90%/20.00%,大专或大学本科毕业者 56.50%/59.50%,研究生毕业者5.30%/11.60%。家庭月收入1000元以下的占3.90%,1000~3000元的占26.30%,3000~6000元的占45.40%,6000元以上的占24.40%。式中,E—林木资源资产经济价值量;K—调整林分质量的系数;AI—盛产期年平均净收益;AJ—寿命期末木材净收益;Ai—初产期各年净收益;u—经营周期;n—林分年龄;n1—盛产期开始时的林分年龄;P—投资收益率。
2.2 研究工具、仪器与材料
2.2.1 青少年抑郁
采用国内学者俞大维和李旭修订的由美国心理学家 Kovacs编制(Children’s Depression Inventory,CDI;Kovacs,1992)的儿童抑郁量表。该量表已在国内外诸多研究中得到了广泛的应用,共包括 27个项目,内容涉及抑郁的一系列症状(例如,“不高兴”、“社交减少”、“感到疲倦”、“睡眠障碍”等),适用于年龄在 7~17岁之间的儿童,由青少年自我报告。采用 3 点记分——0(偶尔)、1(经常)、2(总是),以所有项目的总分代表抑郁状况,得分越高,抑郁水平越高。两次测量中,量表的克隆巴赫系数分别为0.87和0.88。
2.2.2 同伴拒绝与同伴接纳
采用同伴提名法测量儿童的同伴接纳与同伴拒绝。要求被试写出班里最不喜欢的三位同学(消极提名)和最喜欢的三位同学(积极提名)。对每个被试的被提名次数进行班级内标准化,以便于不同班级儿童提名分数的比较。以消极提名的Z分数作为同伴拒绝的指标,积极提名的Z分数作为同伴接纳的指标。在两个时间点的测量中,同伴接纳和同伴拒绝均呈显著负相关,相关系数分别为‒0.35和‒0.32。该方法被认为是测量同伴关系的经典方法(陈欣银,李正云,李伯黍,1994)。
2.2.3 DNA样本采集、提取、分型仪器与试剂
仪器
离心机(Eppendorf公司)、D-37520台式离心机(Thermo公司)、PCR仪(GeneAmp PCR System 9700,Applied Biosystems公司)、MassArray TM Nanodispenser (SAMSUNG 公司)、MassARRAY compact System (SEQUENOM 公司)、G384+10 Spectrochip TM (SEQUENOM 公司)、移液枪(Eppendorf公司)。试剂
HotStarTaq DNA Polymerase (1000 U)(包括 4×250 units HotStarTaq DNA Polymerase、10×PCR Buffer、25 mM MgCl2,Qiagen 公司)、iPLEX TM Reagent Kit (包括 10×SAP Buffer、1 U/uL SAP enzyme、10×iPlex Buffer、iPlex Termination mix、iPlex enzyme,SEQUENOM 公司)、Clean Resin(SEQUENOM 公司)、dNTP Mixture (TaKaRa 公司)。2.3 研究程序
2.3.1 问卷调查程序
以学校为单位,选取山东省济南市 11所中学的 1555名初一学生作为测查对象,并向所测查学生及其监护人发放参与本项目研究的同意信。在取得他们同意的前提下,以班级为单位,采用问卷法进行第一次(2010年)抑郁量表测查,之后连续追踪测评3年(2010~2012年),每年测评一次,剔除因转学等流失被试后,最终共获得有效被试1323名。主试均由经过严格培训的心理学专业教师、研究生和大四本科生担任。
2.3.2 DNA样本采集与分型程序
在征得学生被试及其监护人同意的前提下,以班级为单位采集学生被试的唾液样本,每人采集2~5 ml,最终共获取有效样本1323个。而后将唾液样本交付上海某生物科技有限公司,由该公司专业技术人员进行DNA的提取、纯化与分型。DNA分型利用美国Sequenom公司的MassARRAY系统完成。MAOA
基因引物为F: ACGTTGGATGTGCACTTAAATGACAGTCCC,R: ACGTTGGATGGATTCACTTCAGACCAGAGC。PCR 反应条件:94°C 15 min;94°C 20s,56°C 30s,72°C 1min,共 45 个循环;最终72°C 3min。PCR扩增后,剩余的dNTP将被去磷酸消化掉,反应体系包括 1.53 μL 水、0.17 μL SAP 缓冲液、0.3单位碱性磷酸酶(Sequenom)。该反应在37°C进行40 min,然后85°C 5 min使酶失活。碱性磷酸酶处理后,针对 SNP的单碱基延伸引物在下列反应体系中进行:0.755 μL 水、0.2μL10×iPLEX缓冲液、0.2 μL终止混合物、0.041 μL iPLEX酶(Sequenom),0.804 μL 10uM的延伸引物。单碱基延伸反应在下列条件下进行:94°C 30 s;94°C 5 s,52°C 5 s,80°C 5 s 5 个循环,共 40 个循环;最后72°C 3 min。在终止反应物中加入6 mg阳离子交换树脂(Sequenom)脱盐,混合后加入25 μL水悬浮。使用 MassARRAY Nanodispenser (Sequenom)将最终的分型产物点样到一块 384孔的 spectroCHIP(Sequenom)上,并用基质辅助激光解吸电离飞行时间质谱进行分析。最终结果由MassARRAY RT软件系统(版本号 3.0.0.4)实时读取,并由 MassARRAY Typer软件系统(版本号3.4)完成基因分型分析。2.4 数据处理与分析
采用 Foxbase 2.0进行数据录入,应用 SPSS 19.0进行数据处理与分析。基于横断数据在解释变量间关系方面的局限性,本研究中同伴关系的测量采用的是第一个测量时间点(2010年)的数据,抑郁采用的是最后一个时间点(2012年)的数据,同时分析了基线水平(2010年)抑郁的影响。此外,本研究的主要统计方法为分层回归分析和显著性区域检验,其中后者的统计指标主要包括:(1)显著性区域上限(higher bound)和下限(lower bound),指基因与抑郁显著关联时,同伴关系的取值范围。(2)交互作用比例(the proportion of interaction,POI),指在交互作用的区域中与不同易感性模型相符合的比例。通常若 0.40≤PoI≤0.60,则说明基因‒环境交互作用符合典型的不同易感性模型,若 PoI趋近 0,则意味着完全符合素质‒压力模型。鉴于 PoI会随环境变量的取值范围的变化而变化,因此Roisman等人(2012)推荐使用M
±2SD,
而不是M
±1SD
的区间。因为前者几乎涵盖了95%的取值区域,而后者仅涵盖了 68%,(3)受影响比例指标(the proportion affected index,PA),指大于两条回归线的交叉点所对应的 X值的比例,即交叉点右侧区域所占的比例。若PA近似为0.50 (0.40~0.60),则交互作用模式支持不同易感性模型,若小于 16%,基本表明不符合不同易感性模型,若近似为0,则支持素质‒压力模型。(4) X和ZX:ROS应用的前提条件是环境变量与因变量的关系符合线性假设,因而在模型检验前需要考察X和ZX(Z指调节变量,在本研究中为基因型)是否对因变量具有显著预测作用,如有,则需要在控制其影响的前提下考察交互项是否依然显著。(5)交叉点(cross-over point),指两条回归线交叉点所对应的 X 值,若该点落在[‒2~+2]的区间范围内,则说明基因−环境交互作用为非按序型交互作用,支持不同易感性模型;若该点位于或接近于横坐标的中间(标准化后的取值为 0),则表明基因‒环境交互作用符合典型模式的不同易感性模型。3 研究结果
3.1 变量的描述统计量与相关分析结果
如表1所示,rs6323多态性(T等位基因携带者274人,G等位基因携带者404人)的分布与同伴接纳、同伴拒绝相关均不显著,说明可以排除基因‒环境相关的可能性。rs6323多态性与抑郁相关亦不显著。同伴接纳与同伴拒绝呈显著负相关,因而为避免多元共线性的问题,在后续的分析中分别对同伴接纳和同伴拒绝建构模型进行相应的统计分析。同伴接纳与两个时间点的抑郁均呈显著负相关,同伴拒绝则与两个时间点的抑郁均呈显著正相关。两次抑郁得分之间存在显著正相关,因此为了更细致地考察rs6323多态性与同伴关系,以及二者的交互作用对抑郁(2012)的预测作用,后继分析中,我们同时呈现了控制基线水平抑郁(2010)前、后的结果。
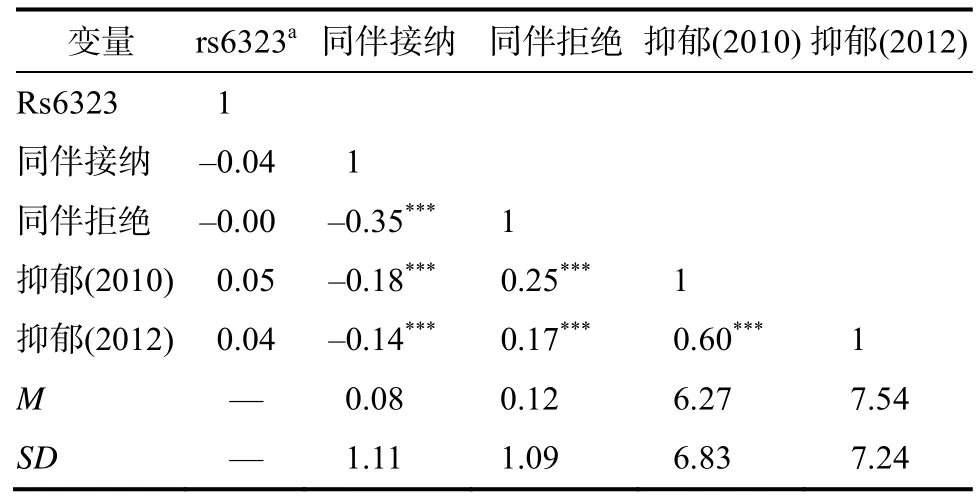
表1 rs6323多态性、同伴关系与抑郁的描述统计量及相关分析结果
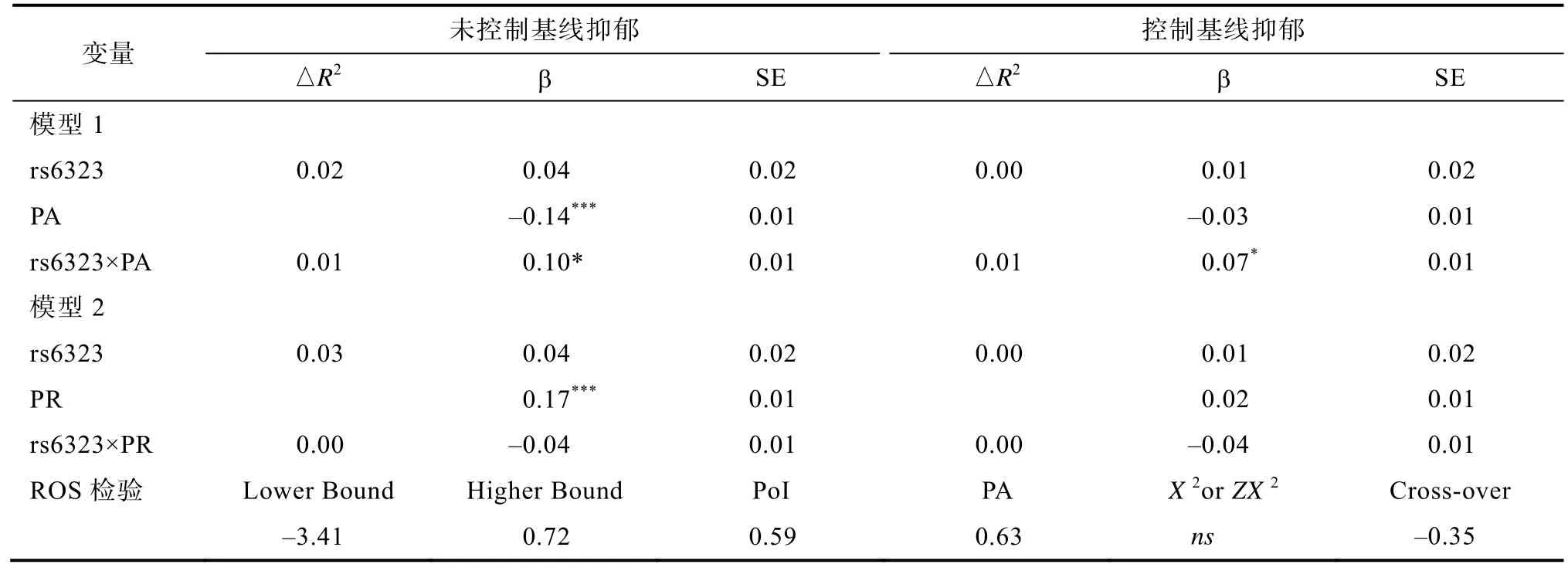
表2 抑郁对青少年的MAOA基因rs6323多态性与同伴关系的分层回归分析
3.2 MAOA基因rs6323多态性与同伴关系对青少年早期抑郁的影响
以2012年时间点的抑郁为因变量,rs6323多态性、同伴接纳或者同伴拒绝,以及二因素的交互项为预测变量进行两大组分层回归分析,第一组未控制基线水平的抑郁(2010),第二组则进行了控制,结果显示(见表2),rs6323多态性对青少年早期抑郁均无显著主效应;同伴接纳则能够显著负向预测青少年的抑郁,同伴拒绝能够显著正向预测青少年抑郁,但在控制基线水平的抑郁后,其预测作用变得不显著;rs6323多态性与同伴接纳对青少年的抑郁具有显著交互作用,且在排除基线水平抑郁的影响后依然显著;rs6323与同伴拒绝的交互作用在控制基线水平抑郁前后均不显著。
为了便于对 rs6323多态性与同伴接纳的交互作用进行简单效应分析,依据目前较为通用的方法,我们将高于平均分 1个标准差的划分为高分组(G等位基因:116人,T等位基因:73人),低于平均分1个标准差的划分为低分组(G等位基因:119人,T等位基因:91人)。简单效应分析结果显示(见图1 左),当同伴接纳(β
=‒0.10,t
=‒1.32,p
>0.05)水平较低时,G等位基因携带者的抑郁水平表现出高于T等位基因携带者的趋势,但差异未达到显著性水平;当同伴接纳水平较高时,G等位基因携带者的抑郁水平显著低于T等位基因携带者(β=0.19,t
=2.61,p
<0.05)。另一方向的简单效应分析进一步显示(见图1右),当rs6323位点为G等位基因时,同伴接纳(β=‒0.08,t
=‒2.00,p
<0.05)可以显著负向预测青少年早期抑郁,当rs6323位点为T等位基因时,同伴接纳(β=0.05,t
=0.95,p
>0.05)对青少年早期抑郁无显著预测作用。
图1 rs6323多态性与同伴接纳对男青少年早期抑郁交互作用
为了进一步探明交互作用的形式,我们又进行了 ROS检验,结果显示(见表2),回归线交叉点(cross-over point)值为‒0.35,基本位于X轴(区间范围为‒2 ~ +2)中间位置;当同伴接纳取值低于‒3.41或者高于0.79时(见图2),rs6323位点不同等位基因携带者的抑郁存在显著差异;POI为 0.59,PA为0.63,且同伴接纳与抑郁不存在非线性关系。这些数据信息表明,同伴接纳与rs6323多态性的确对青少年抑郁存在交互作用,但其作用形式不支持素质‒压力模型,而是基本符合不同易感性模型。

图2 rs6323多态性与同伴接纳对男青少年早期抑郁交互作用的显著性区域
4 讨论
本研究同时采用传统的分层回归与新兴的“显著性区域”检验两种方法,深入考察了MAOA
基因rs6323多态性与同伴关系对男青少年早期抑郁是否存在交互作用,若存在,其显著区域范围是多少,以及作用形式是符合素质‒压力模型,还是符合不同易感性模型的问题。结果发现,rs6323多态性与同伴接纳对男青少年早期抑郁存在显著交互作用,但是该位点多态性与同伴拒绝的交互作用不显著。对 rs6323多态性与同伴接纳对男青少年早期抑郁交互效应的进一步分析显示,当同伴接纳水平较低时,G等位基因携带者的抑郁水平表现出高于T等位基因携带者的趋势。G等位基因编码高活性单胺氧化酶A,尽管既有关于MAOA
基因活性与抑郁关联的研究结果存在分歧,但是 Fan等人(2010)的一项元分析表明,亚洲男性高活性MAOA
等位基因携带者更易罹患重性抑郁。这与本研究结果基本一致。然而,本研究的另一新发现是,当同伴接纳水平较高时,G等位基因携带者的抑郁水平显著低于T等位基因携带者,即在高、低同伴接纳两种条件下,rs6323位点G等位基因与抑郁的关联表现出相反的方向。而且进一步的分析显示,同伴接纳可以显著预测G等位基因携带者的抑郁,但对T等位基因携带者的抑郁无显著预测作用。这提示我们,G等位基因可能并非是风险基因(vulnerability gene),而是对外界环境更为敏感的“可塑基因” (plasticity gene)。该研究结果为既有关于MAOA
基因与抑郁关系的研究结论存在分歧提供了一种可能的解释,即没有考察被试所处环境因素的影响,或者不同研究中被试生活经历的差异,是导致现有关于MAOA
基因与抑郁关系的结论存在分歧的可能原因之一。虽然有关rs6323多态性G等位基因(高活性等位基因)对环境影响更为敏感的内在机理仍有待科学研究的揭示,但是有资料显示,与低活性MAOA
等位基因携带者相比,那些携带高活性MAOA
等位基因个体的右眶额皮质、左丘脑(眶额皮质和左丘脑是人类愉快、悲伤等情绪产生的重要神经机制)对环境刺激表现出更高的激活反应(Sebastian et al.,2010),在识别消极情绪时其大脑右侧前额叶中回与左侧前额叶下回激活程度显著增高(Guo et al.,2014)。这表明在外界刺激作用下,MAOA基因可能通过激活与情绪相关的脑区,进而影响抑郁的发生。高活性等位基因携带者的相关脑区的激活程度更高,因而对外界刺激表现出更高的反应性。该推理能够在一定程度上为本研究结果提供解释,也与当前较为流行的“基因‒脑‒行为”模型(Eisenberger,Way,Taylor,Welch,&Lieberman,2007)观点相吻合。需要说明的是,显著性区域检验的结果进一步表明,rs6323多态性与同伴关系的交互作用模式虽然不支持“素质‒压力模型”,但是只有当同伴接纳的取值区间为[‒∞,‒3.41]或者[0.79,+∞]时 rs6323多态性与抑郁的关联才达到显著。这说明本研究结果只获得了部分支持不同易感性模型观点的证据,即只验证了其关于携带某种基因型的个体在积极成长环境(高同伴接纳水平)下会表现出更低的抑郁水平的假设,而没有支持其携带该基因型个体在消极成长环境(同伴拒绝和低同伴接纳)下也更容易患抑郁的观点。这可能是由于本研究的被试均为正常群体青少年,所报告的同伴拒绝和低同伴接纳的严重程度较低,其影响不足以揭示出消极同伴关系与rs6323多态性的交互作用。正如Roisman等(2012)所指出的那样,没有任何一种有关基因‒环境交互作用的检验方法可以完全不受取样的影响。因此,未来研究可增加处境不利儿童青少年或临床抑郁患者样本,对该结果继续进行考证。
本研究是首项考察同伴关系对MAOA
基因与抑郁关系的调节作用的研究,也是首项考察MAOA
基因rs6323多态性与环境对抑郁交互作用的研究。所以本研究结果有待后续研究的验证。尽管如此,本研究同时采用了传统回归和新兴的显著性区域检验两种方法对基因‒环境交互作用进行分析,所获得的研究结果也经过了 Benjamini和 Hochberg(1995)程序的校正,而且在控制基线水平抑郁的影响后,所发现的同伴接纳与rs6323多态性交互作用依然显著。另外,由于本研究中抑郁和同伴拒绝水平均呈偏态分布,所以我们也尝试了对抑郁和同伴拒绝得分进行平方根转换后重新进行统计分析,结果无实质性改变。这表明了本研究结果的稳定性。本研究的不足之处是采用了单基因‒环境设计,因而与现有同类研究相似(Beach et al.,2010;Klauke et al.,2011),本研究中没有发现MAOA
基因的主效应,所发现的基因‒环境交互效应也较小(仅为 1%)。有研究显示,MAOA
基因与受虐经历的关联受到5-羟色胺转运体(serotonin-transporter-linked polymorphic region,5-HTTLPR
)基因的调节(Cicchetti et al.,2007),MAOA
基因与儿茶酚胺‒氧位‒甲基转移酶 (catechol-O-methyl transferase,COMT
)基因对个体抑郁亦存在显著交互作用(Comasco,Sylvén,Papadopoulos,Sundström-Poromaa,Oreland,&Skalkidou,2011)。因而,未来研究有必要采用多基因‒环境设计进一步探查rs6323多态性对青少年早期抑郁的影响。5 研究结论
本研究综合运用传统分层回归与新兴“显著性区域”检验两种方法,首次考察了MAOA
基因rs6323多态性与同伴关系对男青少年早期抑郁的交互作用,结果发现,同伴接纳可以显著预测G等位基因携带者的抑郁,但对T等位基因携带者的抑郁无显著预测作用;而且在高、低同伴接纳两种条件下,rs6323位点G等位基因与抑郁的关联表现出相反的方向;其交互作用形式基本符合不同易感性模型的观点。Beach,S.R.H.,Brody,G.H.,Gunter,T.D.,Packer,H.,Wernett,P.,&Philibert,R.A.(2010).Child maltreatment moderates the association of MAOA with symptoms of depression and antisocial personality disorder.Journal of Family Psychology,24
(1),12–20.Belsky,J.,&Pluess,M.(2009).Beyond diathesis stress:Differential susceptibility to environmental influences.Psychological Bulletin,135
(6),885–908.Benjamini,Y.,&Hochberg,Y.(1995).Controlling the false discovery rate: A practical and powerful approach to multiple testing.Journal of the Royal Statistical Society.Series B (Methodological),57
(1),289–300.Benjamin,D.,van Bakel,I.,&Craig,I.W.(2000).A novel expression based approach for assessing the inactivation status of human X-linked genes.European Journal of Human Genetics,8
(2),103–108.Cao,C.,Chen,G.H.,Wang,M.P.,Cao,Y.M.,&Zhang,W.X.(2014).Association between MAOA gene and depression.Advances in Psychological Science,22
(12),1899–1910.[曹丛,陈光辉,王美萍,曹衍淼,张文新.(2014).MAOA基因与抑郁的关系.心理科学进展,22(12),1899–1910.]
Caspi,A.,McClay,J.,Moffitt,T.E.,Mill,J.,Martin,J.,Craig,I.W.,… Poulton,R.(2002).Role of genotype in the cycle of violence in maltreated children.Science,297
,851–854.Chen,X.Y.,Li,Z.Y.,&Li,B.S.(1994).Peer relationships:An examination of sociometric status and socio-emotional characteristics.Psychological Science,17
(4),198–204.[陈欣银,李正云,李伯黍.(1994).同伴关系与社会行为:社会测量学分类方法在中国儿童中的适用性研究.心理科学,17(4),198–204.]
Cicchetti,D.,Rogosch,F.A.,&Sturge-Apple,M.L.(2007).Interactions of child maltreatment and serotonin transporter and monoamine oxidase A polymorphisms: Depressive symptomatology among adolescents from low socioeconomic status backgrounds.Development and Psychopathology,19
(4),1161–1180.Comasco,E.,Sylvén,S.M.,Papadopoulos,F.C.,Sundström-Poromaa,I.,Oreland,L.,&Skalkidou,A.(2011).Postpartum depression symptoms: A case-control study on monoaminergic functional polymorphisms and environmental stressors.Psychiatric Genetics,21
(1),19–28.Eisenberger,N.I.,Way,B.M.,Taylor,S.E.,Welch,W.T.,&Lieberman,M.D.(2007).Understanding genetic risk for aggression: Clues from the brain’s response to social exclusion.Biological Psychiatry,61
(9),1100–1108.Fan,M.,Liu,B.,Jiang,T.Z.,Jiang,X.P.,Zhao,H.Z.,&Zhang,J.(2010).Meta-analysis of the association between the monoamine oxidase-A gene and mood disorders.Psychiatric Genetics,20
(1),1–7.Guo,H.R.,Ren,Y.M.,Zhao,N.,Wang,Y.L.,Li,S.Y.,Cui,H.,… Zhang,J.H.(2014).Synergistic effect of 5-HT2A receptor gene and MAOA gene on the negative emotion of patients with depression.Clinical Physiology and Functional Imaging,34
(4),277–281.Jansson,M.,McCarthy,S.,Sullivan,P.F.,Dickman,P.,Andersson,B.,Oreland,L.,… Pedersen,N.L.(2005).MAOA haplotypes associated with thrombocyte-MAO activity.BMC Genetics,6
,46.Kim,S.K.,Park,H.J.,Seok,H.,Jeon,H.S.,Chung,J.H.,Kang,W.S.,… Shin,D.H.(2014).Association study between monoamine oxidase A (MAOA
) gene polymorphisms and schizophrenia: Lack of association with schizophrenia and possible association with affective disturbances of schizophrenia.Molecular Biology Reports,41
(5),3457–3464.Klauke,B.,Deckert,J.,Reif,A.,Pauli,P.,Zwanzger,P.,Baumann,C.,… Domschke,K.(2011).Serotonin transporter gene and childhood trauma—A G×E effect on anxiety sensitivity.Depressed and Anxiety,28
(12),1048–1057.Kovacs,M.(1992).Children’s depression inventory (CDI)
manual
.Toronto: Multi-Health Systems Inc.LaFontana,K.M.,&Cillessen,A.H.N.(2010).Developmental changes in the priority of perceived status in childhood and adolescence.Social Development,19
(1),130–147.Marmorstein,N.,&Hart,D.(2010).Interactions between MAOA genotype and receipt of public assistance: Predicting change in depressive symptoms and body mass index.Journal of Research on Adolescence,21
(3),619–630.Mendle,J.,Harden,K.P.,Brooks-Gunn,J.,&Graber,J.A.(2012).Peer relationships and depressive symptomatology in boys at puberty.Developmental Psychology,48
(2),429–435.Nikulina,V.,Widom,C.S.&Brzustowicz,L.M.(2012).Child abuse and neglect,MAOA,and mental health outcomes:A prospective examination.Biological Psychiatry,71
(4),350–357.Nolan,S.A.,Flynn,C.,&Garber,J.(2003).Prospective relations between rejection and depression in young adolescents.Journal of Personality and Social Psychology,85
(4),745–755.Prinstein,M.J.,&Aikins,J.W.(2004).Cognitive moderators of the longitudinal association between peer rejection and adolescent depressive symptoms.Journal of Abnormal Child Psychology,32
(2),147–158.Roisman,G.I.,Newman,D.A.,Fraley,R.C.,Haltigan,J.D.,Groh,A.M.,&Haydon,K.C.(2012).Distinguishing differential susceptibility from diathesis-stress:Recommendations for evaluating interaction effects.Development and Psychopathology,24
(2),389–409.Rubin,K.H.,Bukowski,W.,&Parker,J.(2006).Peer interactions,relationships,and groups.In N.Eisenberg(Ed.),Handbook of child psychology: Social,emotional,and personality development
(6th ed.,pp.571–645).New York: Wiley.Sabol,S.Z.,Hu,S.,&Hamer,D.(1998).A functional polymorphism in the monoamine oxidase A gene promoter.Human Genetics,103
(3),273–279.Sebastian,C.L.,Roiser,J.P.,Tan,G.C.,Viding,E.,Wood,N.E.,&Blakemore,S.J.(2010).Effects of age and MAOA genotype on the neural processing of social rejection.Genes,Brain and Behavior,9
(6),628–637.Shih,R.A.,Belmonte,P.L.,&Zandi,P.P.(2004).A review of the evidence from family,twin and adoption studies for a genetic contribution to adult psychiatric disorders.International Review of Psychiatry,16
(4),260–283.Stewart,S.M.,Betson,C.L.,Lam,T.H.,Chung,S.F.,Ho,H.H.,&Chung,T.C.F.(1999).The correlates of depressed mood in adolescents in Hong Kong.Journal of Adolescent Health,25
(1),27–34.Voltas,N.,Aparicio,E.Arija,V.,&Canals,J.(2015).Association study of monoamine oxidase-A gene promoter polymorphism (MAOA-uVNTR) with self-reported anxiety and other psychopathological symptoms in a community sample of early adolescents.Journal of Anxiety Disorders,31
,65–72.Wentzel,K.R.(2005).Peer relationships,motivation,and academic performance at school.In A.J.Elliot &C.S.Dweck (Eds.),Handbook of competence and motivation
(pp.279–296).New York,NY: Guilford.Xie,B.,Li,D.,London,S.J.,Palmer,P.H.,Johnshon,C.A.,Li,Y.,… Conti,D.V.(2014).Gender difference in interactions between MAOA promoter uVNTR polymorphism and negative familial stressors on body mass index among Chinese adolescents.Pediatric Obesity,9
,e80–e90.Xu,Z.,Zhang,Z.J.,Shi,Y.Y.,Pu,M.J.,Yuan,Y.G.,Zhang,X.G.,&Li,L.J.(2011).Influence and interaction of genetic polymorphisms in catecholamine neurotransmitter systems and early life stress on antidepressant drug response.Journal of Affective Disorders,133
(1-2),165–173.Xue,K.X.(2010).Association study between the polymorphism of 5-Hydroxytryptamine related genes and unipolar depression in a Chinese Han population
(Unpublished master’s thesis).The Shantou University.[薛坤喜.(2010).中国汉族人群5-羟色胺相关基因多态性与单相抑郁症的关联研究 (硕士学位论文).汕头大学.]
Zhang,J.,Tian,L.M.,&Zhang,W.X.(2013).Relations between peer rejection and early adolescents’ academic achievement: The moderating effects of peer acceptance and friendship support.Psychological Development and Education,29
(4),353–360.[张静,田录梅,张文新.(2013).同伴拒绝与早期青少年学业成绩的关系: 同伴接纳、友谊支持的调节作用.心理发展与教育,29(4),353–360.]
Zhou,Z.K.,Zhao,D.M.,Sun,X.J.,&Ding,X.F.(2006).Children’s experiences with peers and loneliness: A two-year longitudinal study.Acta Psychologica Sinica,38
(5),743–750.[周宗奎,赵冬梅,孙晓军,定险峰.(2006).儿童的同伴交往与孤独感: 一项2年纵向研究.心理学报,38(5),743–750.]
Zimmer–Gembeck,M.J.,Hunter,T.A.,&Pronk,R.(2007).A model of behaviors,peer relations and depression: Perceived social acceptance as a mediator and the divergence of perceptions.Journal of Social and Clinical Psychology,26
(3),273–302.