雷公藤红素诱导肿瘤细胞凋亡机制的研究进展
2015-01-16宋书中伍春莲
宋书中, 伍春莲,2*
(1.西华师范大学生命科学学院,西南野生动植物资源保护教育部重点实验室,四川南充637009;2.西南大学,三峡库区生态环境与生物资源省部共建国家重点实验室,重庆400715)
雷公藤红素诱导肿瘤细胞凋亡机制的研究进展
宋书中1, 伍春莲1,2*
(1.西华师范大学生命科学学院,西南野生动植物资源保护教育部重点实验室,四川南充637009;2.西南大学,三峡库区生态环境与生物资源省部共建国家重点实验室,重庆400715)
雷公藤红素 (celastrol)是一种从传统中药雷公藤根皮中提取出的三萜单体,具有显著的抗癌活性。它作为一种蛋白酶体抑制剂,能影响肿瘤细胞内多种信号通路,如抑制蛋白酶体、IKKα/β激酶、HSP90等,并能激活caspase3/7。本文就雷公藤红素诱导肿瘤细胞凋亡及其机制做一系统综述。
雷公藤红素;蛋白酶体;NF-κB;HSP90;凋亡
雷公藤Tripterygium wilfordii Hook.F.,又名黄藤,是我国传统中草药,在治疗风湿性关节炎、肺结核以及其他慢性疾病时发挥着重要作用。雷公藤红素(celastrol,CSL)是从雷公藤根皮中提取出的一种三萜单体,具有抗炎、治疗神经性退化疾病等多种药理活性[1-3]。近年来,它的抗肿瘤作用也备受关注,其分子机制较为复杂,不过越来越多的实验表明,雷公藤红素在诱导肿瘤细胞凋亡方面表现出巨大的潜力。
1 雷公藤红素的理化性质
雷公藤红素又名南蛇藤素或南蛇藤醇,分子式C29H38O4,红色针状结晶,mp 185~200℃,在425 nm处有最强吸收,易溶有机溶剂,不溶于水。从结构上看,它属于五环三萜类化合物,其芳香酮基上的C2和C6有很强的亲核活性,从而具有抗肿瘤、抗微生物、抗氧化、细胞毒性和抗疟疾等作用,结构见图1。
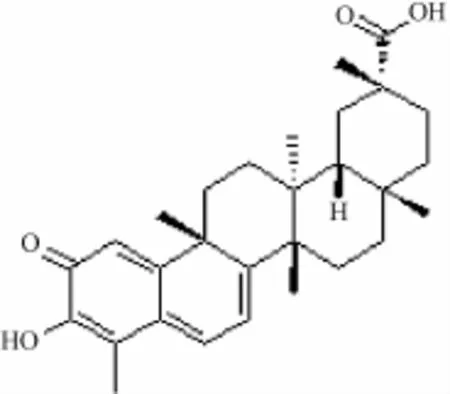
图1 雷公藤红素化学结构式
2 雷公藤红素诱导肿瘤细胞凋亡
近年来的实验表明,低剂量下的雷公藤红素在多种肿瘤细胞凋亡体外实验中都表现出较理想的抑制效果,如乳腺癌细胞、肺癌细胞、胰腺癌细胞、神经胶质瘤细胞、黑色素瘤细胞和前列腺癌细胞等[4-9]。雷公藤红素作为一种天然的蛋白酶体抑制剂[10-11],可影响细胞内多种信号通路及蛋白发挥作用,如NF-κB、Hsp90、蛋白酶体、c-Jun、AKT/mTOR、VEGFR、caspase等[1,4,11-12]。
2.1 作用于β1亚基,抑制蛋白酶体活性 蛋白酶体又称26S蛋白酶体 (沉降系数为26S),是一种催化复合蛋白体,可降解细胞内约80%以上的蛋白质,如周期蛋白、转录因子、IκB-α、促凋亡蛋白Bax以及错误折叠蛋白等。蛋白酶体与细胞中多种信号通路及蛋白发挥作用密切相关,例如NF-κB、Bcl-2家族蛋白等[13-14]。
雷公藤红素作为一种天然蛋白酶体抑制剂,可以诱导多种肿瘤细胞凋亡。Yang等[8]用雷公藤红素 (1~5 mol/L)处理前列腺癌细胞(PC-3和LNCaP),结果发现类胰凝乳蛋白酶的活性明显受到抑制,泛素化的蛋白及蛋白酶体底物 (IκB-α、Bax、p27)大量堆积,由此开始了对雷公藤红素作为蛋白酶体抑制剂的研究。随后其抑制蛋白酶体进而诱导肿瘤细胞凋亡这一作用被Wang等[15]在人宫颈癌细胞Hela中得到确认,但是它是否直接作用于蛋白酶体仍不明确。Feng等[16]发现,雷公藤红素在体外能分别抑制肽基谷氨酰肽水解酶样(peptidyl glutamyl-like)、胰蛋白酶样(trypsin-like)、糜蛋白酶样(chymotrypsin-like)这3类蛋白酶的活性,分别对应β1、β2、β5亚基的催化。但是在以上3种催化亚基中,只有β1的活性可被雷公藤红素抑制,即使在较低的0.2μmol/L浓度下也同样如此。然后,他们将人的蛋白酶体β1亚基在大肠杆菌中克隆表达,发现雷公藤红素能明显地抑制其活性,进一步证实了它是直接抑制蛋白酶体的活性。
蛋白酶体抑制剂可以通过内质网应激,从而诱导肿瘤细胞的凋亡[17]。硼替佐米 (bortezomib)作为唯一一种上市的蛋白酶体抑制剂,能通过内质网应激抑制蛋白酶体降解蛋白,并促进细胞的凋亡[18]。实验证明,在多种肿瘤细胞中,雷公藤红素也可通过抑制蛋白酶体活性,引发内质网应激反应和未折叠蛋白反应[17-20]。Wang等[15]通过Western blot实验发现,雷公藤红素可明显上调HeLa细胞内PERK的表达,同时内质网应激的其他标志物Bip和IREI也得到上调。Feng等[16]也证实,它能激活内质网应激PERK通路的标志物eIF2α和CHOP。另外,Fribley等[21]报道,雷公藤红素还能激活肿瘤细胞eIF2α激酶的活性。
2.2 抑制IκB蛋白降解,抑制NF-κB通路活化 NF-κB(nuclear factor kappa B)即核因子κB,是由Rel蛋白家族中的成员以同源或异源二聚体形式形成的一组转录因子,参与炎症、免疫、应激、细胞的增殖和细胞凋亡等多种生理病理过程的基因调节。大量实验表明,持续性激活NF-κB可以引发人体中大量的恶性肿瘤[22]。
硼替佐米是一种蛋白酶体抑制剂,能抑制NF-κB通路的活化[23],而且雷公藤红素也有同样的抑制作用[24]。NF-κB通路的激发需要IκB蛋白的降解,而其前提是IκB激酶(IKK)的活化。2006年,Lee等[4]报道雷公藤红素能够抑制IKKα和IKKβ激酶的活化。Dai等[10]发现,在雄性激素非依赖型前列腺癌(AIPC)中,它可迅速抑制胞浆中IκBα的降解,阻止RelA进入细胞核。Sethi等[24]报道,雷公藤红素能够抑制诱导型和构成型的NF-κB活化,以及肿瘤坏死因子(TNF-α)诱导IκBα激酶的活化及降解。TAK1在TNF激活IKK过程中发挥着重要作用,而雷公藤红素能够抑制TAK1,Freuddsperger等[25]也进一步证实了这一点。
TMPRSS2/ERG(T/E)基因能促进前列腺癌细胞的增殖和侵袭[26],而NF-κB在此过程中发挥重要作用。Wang等[27]研究表明,NF-κB能通过 p65第 536位丝氨酸(p536)的磷酸化来激活肿瘤细胞T/E的表达。Shao等[28]报道,雷公藤红素是 p536的抑制剂,在前列腺癌细胞(PCa)中通过NF-κB抑制T/E基因的表达。
2.3 作用HSP90 C-末端,抑制HSR活化 热休克反应(heat shock response,HSR)在促进蛋白质折叠过程中起着关键作用,而在多种肿瘤细胞中,这条通路是持续和激活过度的[29]。研究显示,雷公藤红素能在人乳腺癌细胞系MCF-7和BT474、人非小细胞性肺癌细胞系H157、人神经母细胞瘤细胞系SH-SY5Y及HeLa细胞中抑制热休克反应[30]。
人类胞浆中含有两种HSP90亚型,分别为HSP90α和HSP90β。雷公藤红素能够作用于HSP90-Cdc37复合物[6],抑制HSP90 ATP酶活性[31],Cdc-37和ATP结合域位于HSP90 N-末端。Zhang等[32]认为,雷公藤红素可作用于HSP90 C-末端,进而影响HSP90与Cdc37的相互作用。Zamphorlin等[33]通过HSP90α缺失变异体C-HSP90α(只剩C-末端)的SEC-MALS实验,证实雷公藤红素是作用于C-末端结合域从而影响HSP90α的功能。Sreeramulu等[34]报道,它不仅能直接作用于HSP90复合物,而且会将Cdc37作为靶标,从而影响HSP90-Cdc37复合物的形成。Peng等[31]认为,雷公藤红素可能是以共价键的方式与Cdc37氨基端的3个半胱氨酸残基相连,进而影响蛋白激酶的相互作用。此外,它不仅能诱导HSP90分子伴侣 p23的寡聚化[35],而且还是热休克转录因子 (HSF1)的抑制剂[34]。
HSP90维持着细胞内多种激酶、转录因子、受体等的稳定性和活性,如Raf-1、EGFR和AKT[36-38]。雷公藤红素不仅能促进H1650和H1975细胞内EGFR及AKT的降解[39],同时还可促进H460细胞EGFR、ErbB2和凋亡抑制蛋白survivin降解,并提高细胞内p53蛋白表达水平[40]。Huang等[41]发现,无论在常氧或缺氧条件下,雷公藤红素都能够下调HIF-1αmRNA水平,降低核HIF-1α蛋白累积。
2.4 升高Caspase-3/7表达水平 Caspase凋亡途径是细胞凋亡不可或缺的通路。其中,Caspase-3/7被认为是细胞凋亡的执行者[42],一旦被激活就会导致级联的介导细胞凋亡。PARP(poly ADP-ribose polymerase)为DNA修复酶,同时也是Caspase的切割底物[43],常被看作是细胞凋亡的标记物。
实验显示,雷公藤红素能够明显提高多种肿瘤细胞内Caspase3/7蛋白表达水平[39,44-45],同时PARP的剪切带也得到不同程度的增强[39,46-47]。Fan等[39]通过Western blot实验发现,在雷公藤浓度为2μmol/L和4μmol/L时,H1650细胞Caspase3/7蛋白浓度明显升高,PARP也得以激活。Jiang等[44]报道,雷公藤红素联合肝细胞生长因子受体(C-Met)抑制剂PHA-665752(PHA)作用于Huh7细胞,在高浓度(大于1.0μmol/L)时,它能使细胞停滞在G2/M时期并激活Caspase-3/7。另外,Mou等[48]发现,在非小细胞肺癌A549细胞中,雷公藤红素可以促进细胞色素C释放,上调Bax蛋白表达,同时下调Bcl-2蛋白表达。Yang等[45]报道,它可剂量依赖性地诱导乳腺癌细胞凋亡,在MCF-7细胞中,雷公藤红素显著促进了细胞色素C及凋亡诱导因子AIF释放,并降低Bcl-2蛋白表达,上调Bax蛋白表达。
2.5 其他转导途径 在细胞有条不紊地生命代谢的稳态中,稍微出现一个或几个小环节的细小差错,就会诱发整个细胞代谢的紊乱,可能将导致肿瘤细胞的发生。
马永岚等[49]报道,在人早幼粒白血病细胞HL-60中,雷公藤红素能抑制Akt的磷酸化,同时下调CyclinD1蛋白的表达水平,并呈显著的剂量和时间依赖性来诱导细胞凋亡。周培等[50]报道,它能显著抑制PCa细胞株生长,同时降低SENP1基因在mRNA水平表达,表明雷公藤红素可能是通过SENP1相关信号通路来诱导前列腺癌细胞凋亡的。雌激素受体α(ERα)作为一种核转录因子,是治疗乳腺癌的靶点。Jang等[51]发现,在人乳腺癌细胞 MCF7和T47D中,雷公藤红素能降低ERα在mRNA及蛋白水平表达,揭示它可降低MCF7和T47D细胞中ERα的转录活性,对于治疗人乳腺癌有着重大的意义。
3 雷公藤红素的药物作用强度
作为从传统中草药中提取出的单体,雷公藤红素的抗炎、治疗神经退行性疾病及抗癌作用已被广泛报道。作为一种具有巨大潜在价值的抗肿瘤新药,它无论是在肿瘤细胞,还是在肿瘤组织模型方面都表现出较强的抑制肿瘤增殖的特性,并具剂量和时间依赖性。
雷公藤红素针对不同类型的体外培养的肿瘤细胞,其半数抑制浓度 (IC50)也不同,见表1。Feng等[16]用雷公藤红素处理Hela细胞48 h,MTT细胞活性实验检测出其IC50值为(0.79±0.22)μmol/L。
利用肿瘤异体移植模型(Tumor xenograftmodel),Pang等[54]以雷公藤红素2 mg/(kg·d)的量连续腹腔注射16 d,结果发现裸鼠PC-3肿瘤组织体积明显减小,从 (129.36± 37.35)mm3降至 (35.49±9.71)mm3,而对照组肿瘤组织则明显增大,从 (130.63±37.62)mm3升至 (330.74± 72.20)mm3。采用同样的肿瘤异体移植模型,Zheng等[55]以雷公藤红素每2天注射一次裸鼠腹腔,连续21 d,发现随着浓度的增大,它能明显的抑制肺癌细胞95-D组织体积的增大,见表2。
在临床实验中,Zhang等[56]报道,按100μg/kg的剂量,通过静脉注射,雷公藤红素峰浓度Cmax为 (38.83± 12.83)μg/L,血药浓度为 (79.35±19.85)μg/(h·L),半衰期T1/2β为 (8.33±0.84)h,而片剂的生物利用度为17.06%。由此推测,体内代谢和组织分布可能是生物利用度较低的原因。

表1 雷公藤红素作用不同类型细胞系IC50值
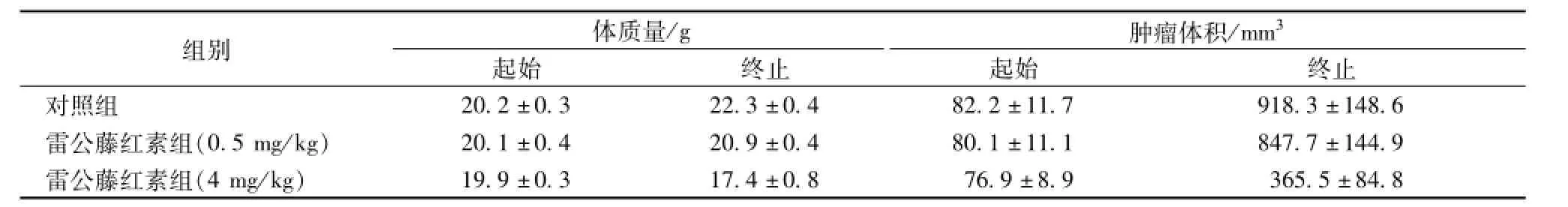
表2 雷公藤红素作用人肺癌95-D细胞裸鼠模型
4 展望
雷公藤作为一种常用药材,在消炎止痛、祛风通络、杀虫解毒方面发挥着重要作用。而来源于其根皮的三萜单体雷公藤红素,近年来在抗肿瘤、抗炎等方面也显示出巨大的潜在治疗价值。另外在低浓度下,雷公藤红素不易产生毒副作用,而其在诱导肿瘤细胞凋亡方面的显著活性正在不断得到实验的证实。
作为一种蛋白酶体抑制剂,雷公藤红素可作用于蛋白酶体β1亚基,从而抑制其活性。它可通过抑制IκB蛋白降解,从而抑制NF-κB信号途径活化。而且,雷公藤红素还能通过作用于HSP90 C-末端结构域,进而影响HSP90与其底物蛋白的相互作用。本课题组认为,雷公藤红素分别通过直接作用于蛋白酶体和HSP90,引起蛋白酶体多种底物蛋白降解异常,以及HSP90多种分子伴侣功能异常。受到抑制的蛋白酶体,通过抑制IκB蛋白的降解,进而抑制NF-κB信号通路的活化,使错误折叠蛋白累积,引发内质网应激反应,再通过激活线粒体凋亡信号途径,进一步诱导肿瘤细胞的凋亡。作为一种提取于我国传统中草药的抗肿瘤药物,雷公藤红素高效低毒,其药用价值正在被不断地发掘,期望能像青蒿素一样为人类的医药事业做出更大的贡献。
[1]Salminen A,Lehtonen M,Paimela T,etal.Celastrol:molecular targets of Thunder God Vine[J].Biochem Biophys Res Commun,2010,394(3):439-442.
[2]Morita T.Celastrol:a new therapeutic potential of traditional Chinesemedicine[J].Am JHypertens,2010,23(8):821.
[3]Kannaiyan R,Shanmugam M K,Sethi G.Molecular targets of celastrol derived from Thunder of God Vine:potential role in the treatment of inflammatory disorders and cancer[J].Cancer Lett,2011,303(1):9-20.
[4]Lee JH,Koo T H,Yoon H,et al.Inhibition of NFkappa B activation through targeting I kappa B kinase by celastrol,a quinonemethide triterpenoid[J].Biochem Pharmacol,2006,72(10):1311-1321.
[5]Zhu H,Liu X W,Cai T Y,et al.Celastrol acts as a potent antimetastatic agent targeting beta1 integrin and inhibiting cellextracellularmatrix adhesion,in part via the p38 mitogen-activated protein kinase pathway[J].J Pharmacol Fxp Ther,2010,334(2):489-499.
[6]Zhang T,Hamza A,Cao X,etal.A novel Hsp90 inhibitor to disrupt Hsp90/Cdc37 complex against pancreatic cancer cells[J].Mol Cancer Ther,2008,7(1):162-170.
[7]Huang Y,Zhou Y,Fan Y,etal.Celastrol inhibits the growth of human glioma xenografts in nude mice through suppressing VEGFR expression[J].Cancer Lett,2008,264(1):101-106.
[8]Yang H,Chen D,Cui Q C,et al.Celastrol,a triterpene ex-tracted from the Chinese“Thunder of God Vine,“is a potent proteasome inhibitor and suppresses human prostate cancer growth in nude mice[J].Cancer Res,2006,66(9):4758-4765.
[9]Dai Y,Desano JT,Meng Y,et al.Celastrol potentiates radiotherapy by impairment of DNA damage processing in human prostate cancer[J].Int J Radiat Oncol Biol Phys,2009,74(4):1217-1225.
[10]Dai Y,Desano J,TangW,et al.Natural proteasome inhibitor celastrol suppresses androgen-independent prostate cancer progression by modulating apoptotic proteins and NF-kappaB[J]. PLoSOne,2010,5(12):e14153.
[11]Kim Y,Kang H,Jang SW,et al.Celastrol inhibits breast cancer cell invasion via suppression of NF-κB-mediated matrix metalloproteinase-9 expression[J].Cell Physiol Biochem,2011,28(2):175-184.
[12]Kannaiyan R,Manu K A,Chen L,et al.Celastrol inhibits tumor cell proliferation and promotes apoptosis through the activation of c-Jun N-terminalkinase and suppression of PI3 K/Akt signaling pathways[J].Apoptosis,2011,16(10):1028-1041.
[13]Li B,Dou Q P.Bax degradation by the ubiquitin/proteasomedependent pathway:involvement in tumor survival and progression[J].Proc Natl Acad Sci U S A,2000,97(8):3850-3855.
[14]Karnak D,Xu L.Chemosensitization of prostate cancer by modulating Bcl-2 family proteins[J].Curr Drug Targets,2010,11(6):699-707.
[15]Wang W B,Feng L X,Yue Q X,et al.Paraptosis accompanied by autophagy and apoptosis was induced by celastrol,a natural compound with influence on proteasome,ER stress and Hsp90[J].JCell Physiol,2012,227(5):2196-2206.
[16]Feng L,Zhang D,Fan C,et al.ER stress-mediated apoptosis induced by celastrol in cancer cells and important role of glycogen synthase kinase-3βin the signal network[J].Cell Death Dis,2013,4(7):e715.
[17]Fribley A,Wang C Y.Proteasome inhibitor induces apoptosis through induction of endoplasmic reticulum stress[J].Cancer Biol Ther,2006,5(7):745-748.
[18]Fels DR,Ye J,Segan A T,etal.Preferential cytotoxicity of bortezomib toward hypoxic tumor cells via overactivation of endop lasmic reticulum stress pathways[J].Cancer Res,2008,68(22):9323-9330.
[19]Bush K T,Goldberg A L,Nigam SK.Proteasome inhibition leads to a heat-shock response,induction ofendoplasmic reticulum chaperones,and thermotolerance[J].J Biol Chem,1997,272(14):9086-9092.
[20]Nawrocki S T,Carew JS,Dunner K J,et al.Bortezomib inhibits PKR-like endoplasmic reticulum(ER)kinase and induces apoptosis via ER stress in human pancreatic cancer cells[J].Cancer Res,2005,65(24):11510-11519.
[21]Fribley A M,Miller JR,Brownell A L,et al.Celastrol induces unfolded protein response-dependent cell death in head and neck cancer[J].Fxp Cell Res,2014,330(2):412-422.
[22]Habraken Y,Piette J.NF-kappaB activation by double-strand breaks[J].Biochem Pharmacol,2006,72(9):1132-1141.
[23]Ganten TM,Koschny R,Haas T L,etal.Proteasome inhibition sensitizes hepatocellular carcinoma cells,but not human hepatocytes,to TRAIL[J].Hepatology,2005,42(3):588-597.
[24]Sethi G,Ahn K S,Pandey M K,etal.Celastrol,anovel triterpene,potentiates TNF-induced apoptosis and suppresses invasion of tumor cells by inhibiting NF-kappaB-regulated gene productsand TAK1-mediated NF-kappaB activation[J].Blood,2007,109(7):2727-2735.
[25]Freudlsperger C,Bian Y,Contag Wise S,et al.TGF-βand NF-κB signal pathway cross-talk ismediated through TAK1 and SMAD7 in a subset of head and neck cancers[J].Oncogene,2013,32(12):1549-1559.
[26]Tom lins S A,Laxman B,Varambally S,et al.Role of the TMPRSS2-ERG gene fusion in prostate cancer[J].Neoplasia,2008,10(2):177-188.
[27]Wang J,Cai Y,Shao L J,etal.Activation of NF-κB by TMPRSS2/ERG fusion isoforms through Toll-like receptor-4[J]. Cancer Res,2011,71(4):1325-1333.
[28]Shao L J,Zhou Z S,Cai Y,et al.Celastrol suppresses tumor cell growth through targeting an AR-ERG-NF-κB pathway in TMPRSS2/ERG fusion gene expressing prostate cancer[J]. PLoSOne,2013,8(3):e58391.
[29]Whitesell L,Lindquist S L.HSP90 and the chaperoning of cancer[J].Nat Rev Cancer,2005,5(10):761-772.
[30]Westerheide SD,Bosman JD,Mbadugha B N,et al.Celastrols as inducers of the heat shock response and cytoprotection[J].Biol Chem,2004,279(53):56053-56060.
[31]Peng B,Xu L,Gao F,etal.HSP90 inhibitor,celastrol,arrests human monocytic leukemia cell U937 at G0/G1 in thiolcontaining agents reversible way[J].Mol cancer,2010,9(1):79-91.
[32]Zhang T,Li Y,Yu Y,et al.Characterization of celastrol to inhibit hsp90 and cdc37 interaction[J].J Biol Chem,2009,284(51):35381-35389.
[33]Zanphorlin LM,Alves FR,Ramos CH.The effectof celastrol,a triterpene with antitumorigenic activity,on conformational and functional aspects of the human 90 kDa heat shock protein Hsp90α,a chaperone implicated in the stabilization of the tumor phenotype[J].Biochim Biophys Acta,2014,1840(10):3145-3152.
[34]Sreeramulu S,Gande S L,Göbel M,et al.Molecularmechanism of inhibition of the human protein complex Hsp90-Cdc37,a kinome chaperone-cochaperone,by triterpene celastrol[J]. Angew Chem Int Fd Fngl,2009,48(32):5853-5855.
[35]Chadli A,Felts S J,Wang Q,et al.Celastrol inhibits Hsp90 chaperoning of steroid receptors by inducing fibrillization of the Co-chaperone p23[J].J Biol Chem,2010,285(6):4224-4231.
[36]Basso A D,Solit D B,Chiosis G,et al.Akt forms an intracellular complex with heat shock protein 90(Hsp90)and Cdc37 and is destabilized by inhibitors of Hsp90 function[J].J Biol Chem,2002,277(42):39858-39866.
[37]Schulte TW,Blagosklonny M V,Ingui C,et al.Disruption of the Raf-1-Hsp90 molecular complex results in destabilization of Raf-1 and loss of Raf-1-Ras association[J].J Biol Chem,1995,270(41):24585-24588.
[38]Sawai A,Chandarlapaty S,Greulich H,et al.Inhibition of Hsp90 down-regulates mutant epidermal growth factor receptor(EGFR)expression and sensitizes EGFR mutant tumors to paclitaxel[J].Cancer Res,2008,68(2):589-596.
[39]Fan X X,Li N,Wu JL,et al.Celastrol induces apoptosis in gefitinib-resistant non-small cell lung cancer cells via caspasesdependent pathways and Hsp90 client protein degradation[J]. Molecules,2014,19(3):3508-3522.
[40]Lee JH,ChoiK J,SeoW D,etal.Enhancementof radiation sensitivity in lung cancer cells by celastrol ismediated by inhibition of Hsp90[J].Int JMol Med,2011,27(3):441-446.
[41]Huang L,Zhang Z,Zhang S,et al.Inhibitory action of celastrol on hypoxia-mediated angiogenesis and metastasis via the HIF-1αpathway[J].Int J Mol Med,2011,27(3):407-415.
[42]Lakhani SA,Masud A,Kuida K,et al.Caspases 3 and 7:keymediators ofmitochondrial events of apoptosis[J].Science,2006,311(5762):847-851.
[43]Gong K,Li W.Shikonin,a Chinese plant-derived naphthoquinone,induces apoptosis in hepatocellular carcinoma cells through reactive oxygen species:A potential new treatment for hepatocellular carcinoma[J].Free Radic BiolMed,2011,51(12):2259-2271.
[44]Jiang H L,Jin JZ,Wu D,et al.Celastrol exerts synergistic effects with PHA-665752 and inhibits tumor growth of c-Met-deficient hepatocellular carcinoma in vivo[J].Mol Biol Rep,2013,40(7):4203-4209.
[45]Yang H S,Kim JY,Lee JH,et al.Celastrol isolated from Tripterygium regelii induces through both caspase-dependentand independent pathways in human breast cancer cells[J].Food Chem Toxicol,2011,49(2):527-532.
[46]杨 溦.ABT-737与雷公藤红素联合抗肿瘤活性的机制研究[D].杭州:浙江大学,2012.
[47]谢 勇,闫燕艳,尉杰忠,等.雷公藤红素诱导胃癌细胞株MGC803凋亡作用研究[J].中药药理与临床,2010,26(5):31-33.
[48]Mou H,Zheng Y,Zhao P,et al.Celastrol induces apoptosis in non-small-cell lung cancer A549 cells through activation of mitochondria-and Fas/FasL-mediated pathways[J].Toxicol In Vitro,2011,25(5):1027-1032.
[49]马永岚,易 虹,杨飞兰,等.雷公藤红素下调HLP-60细胞P-Akt与CyclinD1蛋白表达及其诱导细胞凋亡的效应[J].中国实验血液学杂志,2010,18(4):898-902.
[50]周 培,乔小霞,贺甜甜,等.雷公藤红素对人前列腺癌细胞凋亡及SENP1基因表达的影响[J].现代肿瘤医学,2011,19(11):2153-2157.
[51]Jang SY,Jang SW,Ko J.Celastrol inhibits the growth of estrogen positive human breast cancer cells through modulation of estrogen receptorα[J].Cancer Lett,2011,300(1):57-65.
[52]Ni HW,Zhao W Z,Kong X T,et al.NF-kappa B modulation is involved in celastrol induced human multiple myeloma cell apoptosis[J].PLoSOne,2014,9(4):e95846.
[53]Tozawa K,Sagawa M,Kizaki M.Quinone methide tripterine,celastrol,induces apoptosis in humanmyeloma cells via NF-κB pathway[J].Int JOncol,2011,39(5):1117-1122.
[54]Pang X,Yi Z,Zhang J,etal.Celastrol suppresses angiogenesis-mediated tumor growth through inhibition of AKT/mammalian target of rapamycin pathway[J].Cancer Res,2010,70(5):1951-1959.
[55]Zheng L,Fu Y,Zhuang L,et al.Simultaneous NF-κB inhibition and E-cadherin upregulation mediate mutually synergistic anticancer activity of celastrol and SAHA in vitro and in vivo[J].Int JCancer,2014,135(7):1721-1732.
[56]Zhang J,LiCY,Xu M J,etal.Oralbioavailability and gender-related pharmacokinetics of celastrol following administration of pure celastrol and its related tablets in rats[J].JFthnopharmacol,2012,144(1):195-200.
R966
:A
:1001-1528(2015)07-1548-05
10.3969/j.issn.1001-1528.2015.07.033
2014-11-12
中国博士后基金项目 (2013M540391);四川省教育厅重大培育项目 (13CZ0029);三峡库区生态环境与生物资源省部共建重点实验室开放课题基金项目(SKL-2011-05)
宋书中 (1988—),男,硕士,从事天然药物对肿瘤细胞凋亡作用的研究。
*通信作者:伍春莲 (1976—),女,博士,副教授,从事细胞和分子生物学的研究和教学。Tel:18990874796,E-mail:wcl_xj@ 163.com
