杂合抗菌肽MgJ基因在毕赤酵母中表达条件的优化
2015-01-03尹佳
尹 佳
杂合抗菌肽MgJ基因在毕赤酵母中表达条件的优化
尹 佳
(吉林化工学院生物与食品工程学院,吉林 吉林 132022)
为确定杂合抗菌肽MgJ基因的最佳诱导表达条件,将构建好的重组表达载体pPICZαA-MgJ经SacⅠ线性化处理,电转入巴斯德毕赤酵母GS115,利用抗生素Zeocin筛选阳性克隆,聚合酶链式反应(polymerase chain reaction,PCR)扩增验证,甲醇诱导表达。优化杂合抗菌肽MgJ诱导剂浓度、诱导时间、诱导温度、初始菌体浓度表达条件。结果表明杂合抗菌肽MgJ的最佳表达条件为:28℃、250 r/min诱导培养,每24 h添加体积分数0.5%的甲醇,诱导时间120 h,MgJ表达量可达11.9 mg/L。纯化后获得的表达产物MgJ对S. aureusATCC 29213有较好的抑菌活性,最小抑菌浓度(minimal inhibitory concentration,MIC)为10 ☒g/mL。
杂合抗菌肽MgJ;毕赤酵母;表达条件优化
抗菌肽(antimicrobial peptides,AMPs)是生物界中广泛存在的一类生物活性小肽,具有抗细菌或真菌作用,有些还具有抗原虫、病毒或肿瘤细胞的功能[1-2]。抗菌肽具有广谱的抗菌作用,对革兰氏阳性菌、革兰氏阴性菌、真菌具有抑制作用;可以杀伤动物体内的肿瘤细胞,却又不破坏动物体内的正常细胞[3]。在合成机制、氨基酸组成和作用机制等方面不同于传统的微生物(包括细菌、真菌和链霉菌等)产生的多肽类抗生素[4-5]。因此,抗菌活性肽的开发与应用研究已成为国内外昆虫学、植物学、医药学研究的热点。目前国内外已有采用基因工程的方法制备抗菌肽的报道,由于抗菌肽通常带有阳离子正电荷,与带有阴离子的DNA可能发生相互作用,具有一定的细胞毒性,因此通常采用融合蛋白的形式进行表达[6-7]。
本实验将抗菌肽Magainin-Ⅱ和JaponicinⅡ两个基因进行杂合得到的重组酵母表达载体pPICZαA-MgJ采用电转法转入巴斯德毕赤酵母GS115,对阳性克隆子进行诱导表达,获取杂合抗菌肽MgJ,并进行诱导剂浓度、诱导时间、诱导温度、初始菌体浓度表达条件的优化,获得高效表达的蛋白,以期为生产新型食品生物防腐剂提供参考。
1 材料与方法
1.1菌种与培养基
菌种:宿主菌毕赤酵母GS115,工程菌pPICZαA-MgJ由吉林农业大学食品工程实验室保存。
YPD培养基:酵母提取物10 g/L、胰蛋白胨20 g/L、葡萄糖20 g/L。BMGY培养基:酵母提取物10 g/L、胰蛋白胨20 g/L、K3PO4100 mmol/L、无氨基酵母氮源(yeast nitrogen base without amino acids,YNB)13.4 g/L、生物素4×10-5g/L、甘油10 g/L,pH 6.0。BMMY培养基:5 mL/L过滤除菌的甲醇替代BMGY的10 g/L甘油。YPDS培养基:胰化蛋白胨20 g/L、酵母提取物10 g/L、葡萄糖20 g/L、山梨醇182.2 g/L、琼脂粉20 g/L。水解酪蛋白(Mueller-Hinton)培养基:牛肉粉2 g/L、可溶性淀粉1.5 g/L、酸水解酪蛋白17.5 g/L。
1.2方法
1.2.1毕赤酵母的转化以及筛选
毕赤酵母转化采用电转化法[8]。重组质粒pPICZαAMgJ经SacⅠ线性化处理电转化至感受态细胞毕赤酵母GS115。以空质粒pPICZαA作阴性对照。电转化参数为:电压2 000 V、脉冲5 ms、温度0℃。电击结束后,立即加入1 mL预冷的1 mol/L山梨醇。将混合物转移到灭菌离心管中,30 ℃静置培养1.5 h,取转化物均匀涂布于含100 ☒g/mL Zeocin的YPDS选择平板上[9]。
1.2.2酵母重组子的聚合酶链式反应(polymerase chain reaction,PCR)鉴定
用5’-AOX1引物(5’-GACTGGTTCCAATT GACAAGC-3’)和3’-AOX1引物(5’-GCAAATGGCA TTCTGACATCC-3’)分别作为上、下游引物,进行PCR扩增反应,以鉴定阳性克隆。PCR反应程序为:94℃预变性1 min;94℃变性1 min、56℃退火1 min、72℃延伸1 min,共30个循环;72℃延伸10 min。2%琼脂糖凝胶电泳检测PCR产物。
1.2.3重组酵母GpMgJ的生长动力学
发酵液稀释后于600 nm波长处以去离子水为对照进行比色测定。本实验中的OD600nm值为OD600nm读数与稀释倍数的乘积。从GpMgJ接种到BMGY培养基起开始计时,OD600nm= 6时转入BMMY,每6 h取样测定OD600nm值,绘制其生长曲线。
1.2.4重组杂合肽的表达
将GS115-pPICZαA-MgJ重组菌和对照重组菌GS115-pPICZαA分别接种于2 mL YPD液体培养基中,30℃、250 r/min摇床培养过夜,再以1%的量分别接种于100 mL BMGY培养基中,30℃、250 r/min摇床培养至OD600nm= 2~6。4 000 r/min离心5 min收集菌体,用100 mL BMMY培养基重悬细胞后转移至500 mL摇瓶中,于28℃、250 r/min诱导培养7 d,期间每隔24 h补加100%甲醇使其终体积分数达0.5%~1%,分别在0、24、48、72、96、120、144 h取样,10 000 r/min、4℃离心10 min,取上清液进行Tricine-十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfonate-polyacrylamide gel electrophoresis,SDS-PAGE)检测。
1.2.5表达产物的纯化
取1.2.4节中上清液加等体积无水乙醇,置于-20℃环境2 h,再于4℃、10 000 r/min离心15 min,取上清装入截留相对分子质量3 000的透析膜中,0.05 mol/L pH 6.5的磷酸盐缓冲液(phosphate buffered saline,PBS)于4℃过夜透析除盐脱色。再用25%的聚乙二醇(polyethylene glycol,PEG)6000反渗透浓缩至10 mL。浓缩液经Sephadex G-50层析和CM52阳离子交换层析纯化后浓缩至1 mg/mL,进行发酵上清液中蛋白含量的检测和Tricine-SDS-PAGE检测。
1.2.6重组菌发酵液中蛋白含量的测定
以牛血清白蛋白(bovine serum albumin,BSA)为标准蛋白,采用Bradford蛋白质定量法[10-11]。用1.0 mg/mL的标准蛋白质溶液给各试管分别加入0、0.01、0.02、0.04、0.06、0.08、0.1 mL,然后用无离子水补充到0.1 mL。最后各试管中分别加入5.0 mL考马斯亮蓝G-250试剂,2~5 min后,即可开始用比色皿,在分光光度计上测定各样品在595 nm波长处的吸光度(A595nm),以标准蛋白质量(mg)为横坐标,以吸光度(A595nm)为纵坐标,绘制标准曲线,得曲线方程y=0.197 9x+0.124 5(R2=0.995 1)。由此测出抗菌肽的A595nm值,即可查出未知样品的蛋白质含量。0.5 mg/mL BSA溶液的A595nm约为0.50。
1.2.7重组抗菌肽表达条件的优化
1.2.7.1甲醇体积分数
将GpMgJ重组菌转入25 mL BMMY(pH 6.0)培养液中,于28℃、250 r/min诱导培养,诱导24 h后按0.0%、0.5%、1.0%、1.5%、2.0%的终体积分数追加甲醇;诱导120 h后各取2 mL样品,10 000 r/min离心10 min,收集表达上清纯化,进行蛋白质含量测定。
1.2.7.2诱导时间
将GpMgJ重组菌转入25 mL BMMY(pH 6.0)培养液后,于28℃、250 r/min诱导培养,每24 h按0.5%的终体积分数追加甲醇;分别于24、48、72、96、120、144 h各取2 mL样品,10 000 r/min离心10 min,收集表达上清纯化,进行蛋白质含量测定。
1.2.7.3诱导温度
将GpMgJ重组菌转入25 mL BMMY(pH 6.0)培养液中,分别在24、26、28、30、32℃进行诱导表达,250 r/min诱导培养,每24 h按0.5%的终体积分数追加甲醇;诱导120 h后各取2 mL样品,10 000 r/min离心10 min,收集表达上清纯化,进行蛋白质含量测定。
1.2.7.4初始菌体浓度
将GpMgJ重组菌接种到5mL BMGY培养基中,30℃、250 r/min摇床培养至OD600nm= 5,4℃、4 000 r/min离心10 min,收集菌体;将适量菌体接入25 mL诱导培养基BMMY(pH 6.0)中,使得起始诱导表达浓度分别为OD600nm= 5、10、20、30、40、50。培养120 h,每24 h按0.5%的终体积分数追加甲醇;各取2 mL样品,10 000 r/min离心10 min,收集表达上清液纯化,进行蛋白质含量测定。
1.2.8重组抗菌肽抗菌活性的测定
采用美国国家临床实验室标准委员会(National Committee for Clinical Laboratory Standards,NCCLS)规定的微量稀释法[12]。将供试菌(S. aureusATCC 25923)接种到水解酪蛋白培养基内,37℃培养12~18 h,使细菌处于对数生长期。取1.2.5节纯化的抗菌肽MgJ加入水解酪蛋白培养基中,使其终质量浓度分别为1、5、10、15 ☒g/mL,各加0.1 mL到无菌微量反应板各孔中,每梯度质量浓度重复2孔。同样方法,用100 ☒g/mL的氨苄西林(ampicillin,AMP)作阳性对照,空载体转化酵母表达蛋白为阴性对照。
2 结果与分析
2.1重组酵母菌株的PCR鉴定

图1 重组酵母菌株的PCR鉴定Fig.1 Identification of pPICZαA-MgJ by PCR
由图1可知,6个重组酵母的质粒DNA均扩增出约700 bp的明显条带,而空载体在500~750 bp之间扩增出约580 bp的条带,结果正确。
2.2重组杂合肽的表达及纯化
重组菌GS115-MgJ表达上清液经进行Tricine-SDS-PAGE检测,结果见图2。在发酵96 h后,有约5.3 kD的蛋白条带出现,与推测的MgJ杂合抗菌肽分子质量相同,而pPICZαA转化的GS115空载体对照的诱导上清液无此蛋白条带出现。表明重组酵母菌GS115-MgJ特异表达了MgJ。重组菌离心后上清中杂蛋白含量较多(图2),用Sephadex G-50层析和CM52阳离子交换层析纯化后得到了相对纯度较高的融合蛋白(图3)。

图2 诱导不同时间表达产物的Tricine-SDS-PAGE图Fig.2 Tricine-SDS-PAGE of expressed products
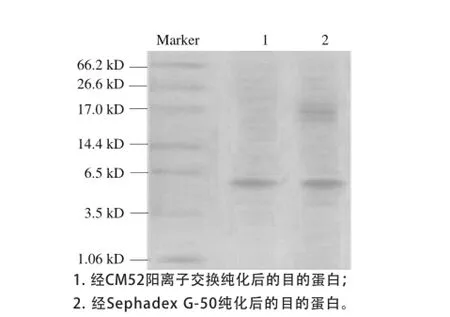
图3 蛋白分离纯化后的Tricine-SDS-PAGGEE图Fig.3 Tricine-SDS-PAGE of the purified antibacterial proteins
2.3重组菌发酵上清液中的蛋白含量

表1 不同时间重组菌GS115-MgJ分泌总蛋白和目的蛋白含量Table 1 Total secreted proteins and target protein in fermentation supernanant of GS115-MgJ
由表1可知,在发酵上清液中,重组菌GS115-MgJ分泌的总蛋白和目的蛋白量随时间的延长而增加。分泌目的蛋白在发酵120 h后达到峰值11.9 mg/L,随后随时间延长呈下降趋势。在发酵144 h时,分泌总蛋白含量达300.6 mg/L。

图4 GpMgJ生长曲线Fig.4 Growth curve of GpMgJ
2.4 重组酵母
GpMgJ菌体生长曲线见图4。18~36 h生长最旺盛,为对数生长期;36~48 h生长变的相对缓慢;48 h时菌体密度(OD600nm)达到最高46.9,此后进入稳定期。
2.5重组杂合肽表达条件的优化
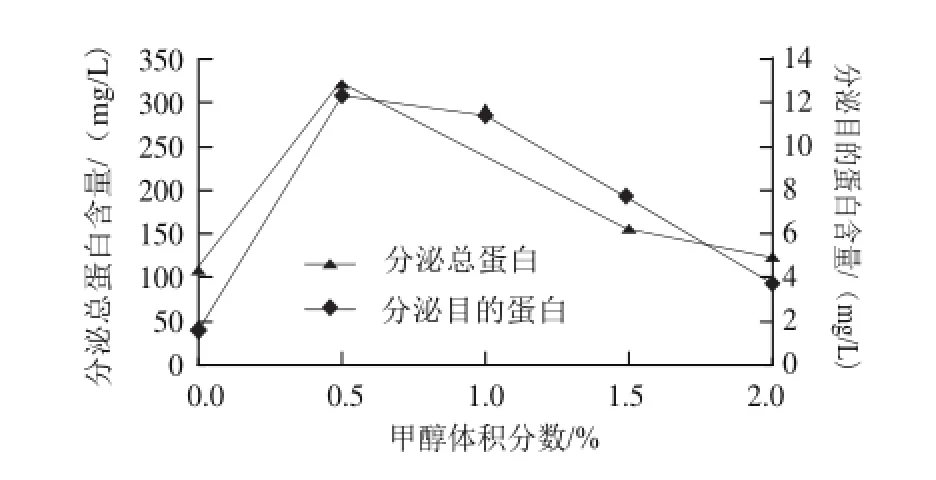
图5 甲醇体积分数对目的蛋白表达的影响Fig.5 Expression of target peptide at different concentrations of methanol
2.5.1甲醇体积分数
由图5可知,甲醇体积分数在0.5%时,分泌总蛋白含量和目的蛋白的表达量均达到最大值,因此,甲醇的最佳诱导体积分数为0.5%。
2.5.2诱导时间
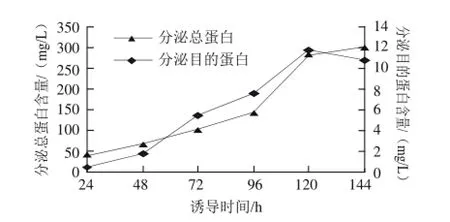
图6 诱导时间对目的蛋白表达的影响Fig.6 Expression of target peptide at different fermentation times
由图6可知,重组菌GpMgJ经过诱导后,分泌总蛋白含量和目的蛋白含量逐渐上升,诱导120 h时目的蛋白达到最高值11.9 mg/L。120 h后,总分泌蛋白量虽然继续上升,但目的蛋白分泌量下降。因此,最佳诱导表达时间为120 h。2.5.3诱导温度由图7可知,随着温度升高或降低,蛋白表达都会受到影响。在28℃诱导表达时,蛋白的表达量最高。因此,最佳表达温度为28℃。

图7 诱导温度对目的蛋白表达的影响Fig.7 Expression of target peptide at different temperatures
2.5.4初始菌体浓度

图8 初始菌体浓度对目的蛋白表达的影响Fig.8 Effect of initial cell concentration on the expression of target peptide
由图8可知,当OD600nm在5~40范围内,随着诱导起始菌体浓度的增加,目的蛋白表达量缓慢增加,当OD600nm超过40后,目的产物的表达量有所下降。
2.6重组杂合肽的抗菌活性

图9 金黄色葡萄球菌的MIC检测结果Fig.9 MIC of MgJ onS. aureusATCC 29213
由图9可知,有细菌生长者呈亮黄色,无细菌生长者为红色。自10 ☒g/mL质量浓度起,较高质量浓度的MgJ能显著抑制S. aureusATCC 29213生长,由此推断,MgJ对S. aureusATCC 29213的最小抑菌浓度(minimal inhibitory concentration,MIC)为10 ☒g/mL。在MIC孔及前后孔各取培养液10μL,转种MH平板,35℃培养24 h,在较低稀释梯度孔没有细菌生长;MIC孔无细菌生长;较高稀释梯度孔有细菌生长,证明结果判定正确。
3 结论与讨论
毕赤酵母表达系统在表达异源蛋白上有诸多优点[13]。本实验对杂合抗菌肽基因表达的诱导剂体积分数、诱导时间、诱导温度、初始菌体浓度4个表达条件进行了优化。
当诱导剂甲醇体积分数高时不仅对目的蛋白的表达具有抑制作用,也会抑制细胞的生长甚至导致细胞的死亡;甲醇体积分数较低时,甲醇会成为限制性底物,限制蛋白表达。因此,过高或过低的甲醇体积分数对毕赤酵母表达外源蛋白都不利[14-16]。本研究结果显示甲醇的最佳诱导体积分数为0.5%。
外源目的蛋白MgJ在培养基中的积累是一个动态的过程,一方面毕赤酵母不断分泌表达MgJ,另一方面MgJ又不断被降解,因此,MgJ在培养基中的积累随时间的变化而变化[17]。本研究结果显示,在诱导120 h以后,培养基中的总分泌蛋白虽然继续增加,但目的蛋白分泌量下降。可能是由于目的蛋白的分泌减少,而胞外酶对目的蛋白水解作用加剧的结果。
毕赤酵母最适生长温度在28~30℃之间[18]。研究结果证实,在28℃发酵时,外源MgJ的表达量较高,高于或低于此范围都将显著影响其表达量。
在一般情况下,细胞光密度(OD600nm)值随着接种量的增加而呈上升趋势。随着诱导起始菌体浓度的增加,目的蛋白表达量也会缓慢增加,但也有研究证实,过大的初始菌体浓度反而会造成目的蛋白表达量的下降[19]。这可能与表达过程中培养基的溶氧有关。发酵过程中具有足够的溶氧,菌体浓度越大,目的蛋白的表达量也应越高。高密度发酵可以提高单位培养上清中分泌蛋白的含量,但随着重组菌生长稳定期的到来,菌体的繁殖速率下降,死亡速率逐步上升,虽然菌体的总数没有变化,可蛋白酶等代谢产物的分泌却逐步增多,使得外源蛋白被降解而影响表达量。因此,可以通过有效控制初始菌体浓度与表达时间的相互关系以获得最大表达量[20]。
由此可见,获得了杂合抗菌肽MgJ最佳表达条件且表达产物抗菌活性良好。重组表达的杂合抗菌肽MgJ在体外具有显著的抑制细菌生长的作用,可作为抗微生物活性物质应用于制备食品防腐剂[21],为开发新型抗菌剂提供了参考。
[1]王军,庞广昌.抗菌肽抗菌机理的研究现状及趋势[J].食品科学, 2005, 26(8): 526-529.
[2]胡功铃,陈国平,胡宗利,等.杂合抗菌肽HMCM的原核表达及其活性鉴定[J].药物生物技术, 2012, 19(1): 6-10.
[3] HOSKIN D W, RAMAMOORTHY A. Sudies on anticancer activities of antimicrobial peptides[J]. Biochimica et Biophysica Acta-Biomembranes, 2008, 1778: 357-375.
[4]张志强,黄庆洲,李英伦,等.抗菌肽重组表达体系及基因表达调控研究进展[J].中国兽医杂志, 2012, 49(7): 58-60.
[5] BOLINTINEANU D, HAZRATI E, DAVIS H T, et al. Antimicrobial mechanism of pore-forming protegrin peptides: 100 pores to killE. coli[J].Peptides, 2010, 31(1): 1-8.
[6] JIN Fengliang, XU Xiaoxia, WANG Liexi, et al. Expression of recombinant hybrid peptide cecropinA (1-8)-magainin2 (1-2) inPichia pastoris: purification and characterization[J]. Protein Expression and Purification, 2006, 50(2): 147-156.
[7] CEREGHINO J L, CREGG J M. Heterologous protein expression in the methylotrophic yeastPichia pastoris[J]. FEMS Microbiology Reviews, 2000, 24(1): 45-66.
[8]高炳淼,唐天乐,长孙东亭.毕赤酵母高效电转化条件的研究[J].中国海洋药物, 2010, 29(2): 1-5.
[9]张宇. RML脂肪酶基因在毕赤酵母中的高效表达与发酵优化[D].广州:华南理工大学, 2011.
[10]李增婷,谢力,冯顺利,等.杂合抗菌肽LPCB在毕赤酵母中的表达条件优化[J].应用与环境生物学报, 2012, 18(3): 65-69.
[11]郭娜.东北林蛙皮肤抗菌肽dybowskin-lsT毕赤酵母表达体系的构建[D].哈尔滨:东北林业大学, 2011.
[12]葛平,潘云华. 1998年美国NCCLS抗生素最小抑菌浓度(MIC)测定法规介绍[J].上海医学检验杂志, 1998(3): 13-18.
[13] MU Xupeng, KONG Ning, CHEN Weili, et al. High-level expression, purification, and characterization of recombinant human basic fibroblast growth factor inPichia pastoris[J]. Protein Expression and Purification, 2008, 59(2): 282-288.
[14]董娜,马清泉,单安山,等.β-发卡抗菌肽的全新设计及其生物学活性[J].生物工程学报, 2012, 28(2): 243-250.
[15]马青山,余占桥,韩冰,等.抗菌肽融合表达研究进展[J].生物工程学报, 2011, 27(10): 1408-1416.
[16]周宇荀,曹巍,魏东芝,等.抗菌肽Adenoregulin基因工程菌培养条件的优化及分批发酵研究[J].生物工程学报, 2005, 21(4): 615-621.
[17] 张兰廷,刘忠渊,张富春.抗菌肽结构与功能关系及分子改造研究进展[J].生物技术, 2008, 18(5): 89-92.
[18]方曙光,储炬,黄立,等.温度对巴斯德毕赤酵母表达瑞替普酶(reteplase)影响[J].工业微生物, 2007, 37(4): 13-14.
[19]李青,周晓宏.新型生物防腐剂:多聚阳离子抗菌肽在毕赤酵母中的表达[J].食品科学, 2013, 34(5): 161-166. doi: 10.7506/spkx1002-6630-201305034.
[20]尹娜,李鸿钧,彭梅.抗菌肽Cecropin D在毕赤酵母中的表达、纯化及活性鉴定[J].中国生物制品学杂志, 2008, 21(3): 185-189.
[21]王宜文,杜兵.应用于食品工业的天然肽类防腐剂[J].农技服务, 2007, 24(2): 105-106.
Optimization of Expression Conditions for Hybrid Antimicrobial Peptide MgJ in Pichia pastoris
YIN Jia
(College of Biology and Food Engineering, Jilin Institute of Chemical Technology, Jilin 132022, China)
The objective of this study was to determine the optimal induction conditions for the expression of hybrid antimicrobial peptideMgJinPichia pastoris. TheSacI-linearized recombinant expression vector pPICZαA-MgJ was transformed intoPichia pastorisGS115 by electroporation. The positive clones were screened with Zeocin and identified by PCR. The protein expression was induced by methanol and the expression conditions of hybridantimicrobial peptide MgJ were optimized. The results showed that the optimum expression conditions of MgJ were as follows: 28℃, 250 r/min, adding 0.5%(V/V) methanol to the culture every 24 h, and induced expression time 120 h. The amount of expressed MgJ could reach up to 11.9 mg/L. After purification, the expressed product MgJ had better antimicrobial activity onS. aureusATCC 29213 with a minimal inhibitory concentration (MIC) of 10μg/mL.
hybrid antimicrobial peptide MgJ;Pichia pastoris; optimization of expression conditions
Q78
1002-6630(2015)11-0162-05
10.7506/spkx1002-6630-201511031
2014-07-02
吉林化工学院科学技术研究项目(2012066)
尹佳(1983—),女,讲师,硕士,研究方向为食品生物技术与功能性食品。E-mail:yinjia612@163.com
