腐植酸对小麦扬花期水分利用效率及灌浆进程的影响
2015-01-01张卓亚王晓琳许晓明
张卓亚, 王晓琳, 许晓明, 李 贵
(1.南京农业大学生命科学学院,江苏 南京 210095;2.江苏省农业科学院植物保护研究所,江苏 南京 210014)
腐植酸(Humic acid,HA)是动、植物的遗骸经过微生物的分解和转化以及在自然或人工物理化学反应作用下形成的一类成分复杂的天然有机高分子聚合物[1-3]。腐植酸中含有吲哚酸、赤霉素、萘乙酸、水杨酸等典型植物生长素[4,对植物生长具有促进作用[5]。而且土壤施用腐植酸类物质可以增加小麦对氮的吸收,叶面施用腐植酸可增加小麦对磷、钾、镁、铜、钙、锌的吸收[6]。在重金属污染土壤中施腐植酸肥料还可以明显减轻重金属元素对小麦的生理伤害[7],而腐植酸浸种还可以缓解盐胁迫下小麦幼苗的质膜损伤[8]。腐植酸还可以提高叶片的SPAD值和光合效率[9]。另外,腐植酸分子中的多元酚结构可作为氧的活化剂和氢的接受体,提高植物体内的氧化还原势,增强植物的呼吸作用[1]。总之,已有的研究结果表明腐植酸直接或者间接地影响了植物代谢、营养、呼吸等生理功能,具有调节植物的生理功能和改善植物生长发育的作用。在目前的农业生产中,腐植酸主要作为土壤改良剂、植物生长调节剂、肥效增进剂和农产品品质改良剂[10]用于农作物叶面喷施及滴灌。有研究结果表明腐植酸的生长调节作用可以贯穿作物生长的始终,尤其生长后期能发挥更大的作用[11]。
小麦开花后是产量和品质形成的关键时期[12],花后新合成的碳水化合物占籽粒干物质的70%~90%[13],对籽粒产量形成有重要作用。同时,冠层叶片的功能状况决定着籽粒灌浆物质的生产与积累[14]。在生产实践中,延缓叶片衰老,延长叶片功能期有利于提高籽粒质量和作物的最终产量[15]。适当增施肥料不仅能显著提高作物光合速率和叶绿素含量,延缓叶片衰老,而且有利于延长作物叶片功能期,在生育后期保持一定的光合面积和光合时间,促进籽粒灌浆[12]。由于冬小麦在拔节期后根系活力逐渐降低[16],尤其是进入生殖生长阶段,地下部营养会集中向顶部运输,根系逐渐衰老[17],影响水分利用与物质运输。而与根部土壤施肥相比,叶面施肥养分吸收快,利用率高,用量少,可以及时补充生长后期由于根部不发达或根系功能衰退而导致的养分吸收不足,起到增产作用[18]。因此在小麦抽穗开花期喷施叶面肥效果最好[19]。
小麦是需水较多的主要粮食作物,干旱缺水是影响小麦增产最重要的非生物限制因素[20]。近年来,中国以华北为中心的冬麦区冬春气象干旱呈加剧趋势,而冬春降水的减少直接导致农业尤其是冬小麦灌溉需水量的增加[21]。在小麦的不同生育时期,灌浆期的干旱胁迫对小麦生长发育及产量影响最大[22]。与传统耕作相比,少免耕可以提高地下水的利用效率[23]。同时旱作条件下进行少免耕有利于旗叶在灌浆后期保持较高的光合能力[24],并提高小麦的籽粒产量[25]。但多年少免耕会导致土壤坚实,土壤容质量增大,影响作物根系对土壤养分和水分的吸收[26]。因此提高浅耕条件下小麦生长后期的水分利用效率不仅有助于改善小麦对有限水资源的高效利用[27],而且有助于协调根冠生长状况,扩大库容量,提高小麦生长潜力和产量。
综上所述,增强小麦生殖生长阶段的根系水分利用效率和功能叶片光合速率将有助于小麦生育后期根冠关系的相互协调,共同促进小麦增产。而腐植酸作为一种良好的植物生长调节物质,在大田作物上的应用相对较少,尤其是在浅耕条件下小麦生育后期腐植酸茎叶处理对水分利用、灌浆进程等生理功能的影响还未见报道。本研究通过考察在小麦扬花期叶面喷施腐植酸的效果,探讨腐植酸对小麦生育后期水分利用效率、灌浆进程、产量及其构成因子的影响,为小麦生育后期的生长调控技术积累理论依据。
1 材料与方法
1.1 试验材料
小麦品种扬麦16、宁麦13由江苏省农业科学院粮食作物研究所提供;腐植酸[黄腐酸≥50%,氮(N)≥2%,钾(K2O)≥12%,有机质≥65%,黄腐植酸盐(FAS)≥70%],新疆双龙腐植酸有限公司生产。
1.2 试验设计
试验于2013年在江苏省农业科学院植物保护研究所实验场进行。土壤为马肝土,pH值7.5,土壤有机质含量11.0 g/kg,有效氮(N)5.1 g/kg,全磷(P2O5)1.3 g/kg,全钾(K2O)6.6 g/kg。每小区2 m×2 m。小麦于2013年11月21日播种,播种密度为150 kg/hm2。2014年4月23日(小麦扬花期)分别用 0 g/L、4 g/L、8 g/L、12 g/L腐植酸叶面喷雾,喷液量为900 L/hm2。每个处理4次重复,随机区组排列。常规防治病虫。
1.3 测定方法
1.3.1 叶片光合气体交换参数及水分利用效率的测定 腐植酸处理后7 d,每小区选取3株长势均匀的小麦植株,用LI-6400型便携式光合系统分析仪(美国LI-COR公司产品)测定旗叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Cond)、胞间CO2浓度(Ci)等光合气体参数。测量条件为:自然CO2浓度,红蓝光源(LI-6400-02B LED),光有效辐射强度(PAR)为1 000 μmol/(m2·s),测定时间为晴天上午 9∶00-11∶30。水分利用率(WUE)=净光合速率(Pn)/蒸腾速率(Tr);气孔限制值(Ls)=1-胞间CO2浓度(Ci)/大气 CO2浓度(Ca)[28]。表观叶肉导度(AMC)= 净光合速率(Pn)/胞间 CO2浓度(Ci)[29]。
1.3.2 穗质量增加速率及源库增量比的测定 穗质量增加速率的测定参考翟虎渠等[30]的方法,分别于小麦喷施腐植酸后 0 d、7 d、14 d、21 d、28 d 测定。取长势均匀、穗型大小一致的单茎5个,测定叶面积后将穗与茎蘖(茎、叶和鞘)分开,放入105℃烘箱中杀青15 min,然后80℃烘48 h至恒质量,分别称质量。计算公式:穗质量增加速率=(G2-G1)/0.5×(S1+S2)×D,式中,G1为第1次平均穗质量(g),G2为第2次平均穗质量(g),S1为第1次平均单茎叶面积(m2),S2为第2次平均单茎叶面积(m2),D为第1次取样至第2次取样的天数。源库增量比的测定参考曹树青等[31]的方法。从每小区选取长势均匀的5个单茎,105℃烘箱中杀青15 min后80℃烘4 h至恒质量。源库增量比=(W2-W1)/(G2-G1),式中W1为抽穗后第28 d的平均单茎干质量(g),W2为抽穗后第35 d的平均单茎干质量(g),G1为抽穗后第28 d的平均穗质量(g),G2为抽穗后第35 d的平均穗质量(g)。
1.3.3 产量及产量构成因素的测定 2014年5月30日收获,在自然风干后,分别统计各小区小麦有效穗数、穗粒数、千粒质量、结实率等性状,并实测各小区小麦产量。
1.4 数据分析
采用DPS7.05及Origin8.5软件对试验数据进行统计分析和作图,并使用Duncan’s新复极差法对平均数进行多重比较。
2 结果与分析
2.1 腐植酸叶面处理对小麦叶片光合特性的影响
腐植酸处理对小麦光合参数的影响如表1所示,扬麦16旗叶的净光合速率(Pn)、蒸腾速率(Tr)和气孔限制值(Ls)随着腐植酸浓度的增加呈现上升趋势。其中4~8 g/L腐植酸叶面处理,扬麦16的气孔限制值增加了18.75%~31.25%,而气孔导度(Cond)和胞间CO2浓度(Ci)分别降低了17.69% ~24.04%和5.78%~7.36%,且与对照有显著差异。宁麦13旗叶的Pn和Cond随腐植酸浓度增加呈先增加后降低的趋势,8 g/L腐植酸叶面处理下分别比对照显著增加了20.83%和29.98%。说明腐植酸叶面处理能显著促进小麦的光合作用。
田间自然条件下(未使用腐植酸处理),宁麦13旗叶的Pn、Tr和Ls显著高于扬麦16,而Ci显著低于扬麦16。4 g/L腐植酸叶面处理后,宁麦13的Cond、Ci和Tr显著高于同浓度处理下的扬麦16,而Ls显著低于扬麦16。说明腐植酸可能通过降低扬麦16的Cond和Ci,以及提高Ls,影响了扬麦16和宁麦13光合速率的差异显著性。
2.2 腐植酸叶面处理对小麦旗叶水分利用效率和表观叶肉导度的影响
腐植酸叶面处理后,扬麦16和宁麦13旗叶的水分利用效率(WUE)和表观叶肉导度(AMC)随腐植酸浓度的增加都呈先上升后降低的趋势(图1)。与对照相比,4~8 g/L腐植酸处理后,扬麦16的WUE显著增加了14.90%~26.92%。在腐植酸浓度4 g/L时,扬麦16旗叶WUE和AMC增加最为显著,分别增加了26.92%和13.64%。而在腐植酸浓度为8 g/L时,宁麦13的WUE和AMC增加最显著,分别增加了15.94%和20.37%。无腐植酸处理(空白对照)时,宁麦13旗叶的 WUE和AMC显著高于扬麦16,这可能与小麦不同的遗传背景有关。表观叶肉导度是用于通过净光合速率与胞间CO2浓度的关系估测非气孔限制作用,从而间接估计RuBP羧化酶的活性[29]。上述结果说明低浓度的腐植酸可以促进小麦的WUE和AMC,而高浓度腐植酸会抑制小麦的WUE和AMC,且不同小麦品种对腐植酸的敏感度不同。

表1 腐植酸叶面处理对小麦光合特性的影响Table 1 Effects of foliar spraying humic acid on photosynthetic characteristics of wheat
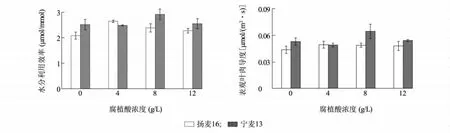
图1 腐植酸叶面处理对小麦水分利用效率和表观叶肉导度的影响Fig.1 The effect of foliar spraying humic acid on water use efficiency and apparent mesophyll conductance of wheat
2.3 腐植酸叶面处理对小麦灌浆进程的影响
随着灌浆进程的推进,扬麦16和宁麦13的穗质量增加速率呈先增加后降低的趋势,且都在叶面喷施腐植酸后21 d达到最大,然后逐渐降低。腐植酸叶面处理后7 d,腐植酸显著增加了扬麦16的穗质量增加速率。其中8 g/L腐植酸处理的扬麦16和宁麦13在第21 d时穗质量增加速率达到最大,同时扬麦16的最大穗质量增加速率显著低于宁麦13(图2)。源库增量比反映了抽穗后叶片光合作用与籽粒灌浆的切合程度[31]。与对照相比,扬麦16的源库增量比显著降低,且随着腐植酸浓度的增加而呈先降低后增加的趋势;而宁麦13的源库增量比随腐植酸浓度的增加而呈上升趋势(图3)。上述结果表明,腐植酸叶面处理显著促进了宁麦13的灌浆进程,且在灌浆后期作用更为明显。
2.4 腐植酸叶面处理对小麦产量及产量构成因子的影响
产量及构成因子分析结果(表2)表明,随着腐植酸浓度的增加,扬麦16的千粒质量、穗粒数和产量有增加的趋势,其中8~12 g/L腐植酸叶面处理后,扬麦16千粒质量增加3.9% ~7.2%,穗粒数增加 4.4%~8.9%,产量显著增加 17.34%~23.42%;宁麦13的千粒质量随腐植酸浓度的增加呈上升趋势,而穗粒数和产量呈先增加后降低趋势,其中,8~12 g/L腐植酸叶面处理后,宁麦13的千粒质量增加16.29% ~20.90%,穗粒数增加3.53% ~12.68%,产量显著增加 17.44%~28.18%。说明腐植酸叶面处理能有效促进小麦的千粒质量和穗粒数,而高浓度腐植酸可能对宁麦13有一定的抑制作用。无腐植酸处理时,扬麦16与宁麦13的千粒质量、穗粒数、有效穗数和产量之间没有显著差异;当4~8 g/L腐植酸叶面处理后,宁麦13的千粒质量和产量显著高于同浓度处理下的扬麦16,而穗粒数没有显著差异,可见宁麦13较扬麦16产量的提高可能是由于腐植酸处理对宁麦13千粒质量的促进效果显著大于扬麦16。
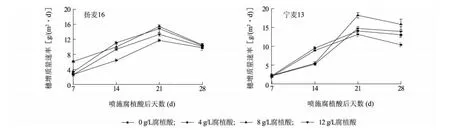
图2 灌浆进程中腐植酸对小麦穗增质量速率的影响Fig.2 The effect of humic acid on grain filling rate of wheat during grain filling
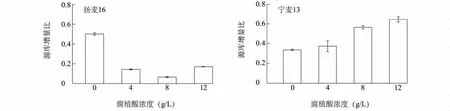
图3 腐植酸对小麦源库增量比的影响Fig.3 The effect of humic acid on the incremental ratio of source to sink of wheat
3 讨论
叶面肥可促进作物体内养分代谢平稳,增强作物抗逆性,提高产量,改善品质[32]。小麦生育后期增施叶面肥能提高冬小麦根系活力,增进旗叶及茎秆的养分更多地向籽粒运输[33]。目前研究结果表明,腐植酸类叶面肥在小麦[34]和水稻[35]等作物上使用,不仅能显著提高作物产量,而且有刺激生长、改善品质和提高抗逆能力等作用[36]。本研究通过田间试验探讨了浅耕条件下腐植酸对小麦生育后期叶片光合性能、水分利用效率和灌浆进程等的影响,结论是腐植酸叶面处理可以提高小麦的光合速率,改善水分利用效率并促进灌浆进程。
光合作用是植物最基本的生理过程[37],小麦产量的90%~95%来自光合作用,尤其是生育后期功能叶光合产物对籽粒的贡献可达80%,其中30%左右来自旗叶[38]。刘建辉等[39]的研究结果表明小麦开花后较高的光合能力及较长的光合持续期是提高千粒质量,进而提高产量的重要生理基础。可见,生育后期旗叶的光合功能对小麦产量的提高有重要作用。1,5-二磷酸核酮糖羧化酶(Rubisco)是碳同化的限速酶,也是限制作物光合速率的重要因素[40]。许多研究证明[41-42],叶片光合能力的提高与RuBisco活性提高和含量增加有关。并且Rubisco的含量及活性与光合速率呈正相关[43]。而Rubisco活性可以用表观叶肉导度来间接估测[29]。本试验结果表明,随着腐植酸浓度的增加,扬麦16和宁麦13的净光合速率基本呈上升趋势,且与表观叶肉导度变化趋势基本一致,说明腐植酸叶面处理下小麦叶片净光合速率的增加,可能与腐植酸叶面处理下Rubisco活性或含量的增加有关。
灌浆期是小麦产量形成的重要时期,而较高的光合生产力是作物获取较高籽粒产量的物质基础。冠层叶片的功能状况决定着籽粒灌浆物质的生产与积累,其早衰会导致小麦产量下降[44]。而小麦灌浆期间适宜的水分状况,不仅能减缓高温下由于缺水引起的早衰,还有利于光合产物的形成和向籽粒的转移、贮藏,并有效提高粒质量[45]。水分利用效率可以体现出作物对水分利用的综合效应,而不同作物种间和同一作物不同品种间水分利用效率都存在明显差异,其中小麦品种间差异可达50%以上[46]。在小麦灌浆期,叶片水分利用效率与光合速率呈显著正相关,与蒸腾速率和胞间CO2浓度呈负相关,且达到极显著水平[47]。本试验结果表明,8 g/L腐植酸叶面处理后,宁麦13的净光合速率显著高于对照,而蒸腾速率和胞间CO2浓度与对照无显著差异,而扬麦16的净光合速率和蒸腾速率与对照无显著差异,但胞间CO2浓度显著低于对照。可见,8 g/L腐植酸叶面处理可以显著提高小麦的水分利用效率。同时,在田间自然条件下,宁麦13的水分利用效率显著高于扬麦16。
小麦籽粒的灌浆速率主要受遗传控制[48],且小麦的灌浆速率整体表现为慢快慢,但不同品种的灌浆进程差异较大。而生育后期叶片的光合同化产物能否同步满足籽粒灌浆的需求是影响高产小麦品种产量潜力发挥的重要因素之一[49]。本试验结果表明,8 g/L腐植酸处理可以显著增加灌浆中后期扬麦16和宁麦13的穗质量增加速率,促进小麦光合产物的积累,有助于小麦千粒质量的增加。在产量及产量构成因子方面,叶面喷施腐植酸可以显著增加千粒质量和产量等[35]。本试验中,腐植酸叶面处理明显增加小麦的千粒质量和穗粒数,从而增加了产量。
综上所述,腐植酸叶面处理下小麦叶片净光合速率的增加,可能与腐植酸叶面处理下Rubisco活性或含量的增加有关,而小麦旗叶净光合速率的增加有助于开花后小麦光合产物的积累,促进源的增加。同时,水分利用效率的提高有利于提高小麦的库容量,提高小麦的产量。另外,腐植酸叶面处理对扬麦16和宁麦13光合生理指标和灌浆进程的影响不同可能是由于有不同的遗传背景导致。总之,腐植酸叶面处理可以作为提高小麦水分利用效率和促进灌浆进程的一种有效措施。
[1] 郑 平.煤炭腐殖酸的生产和应用[M].北京:化学工业出版社,1991:217-280.
[2] 张盛华,郑凯琪,薛红波,等.城市污泥堆肥过程中腐殖酸及重金属形态的变化[J].江苏农业学报,2014,30(6):1350-1354.
[3] 罗晓辉,唐旺全,邱孟斌.添加不同组合生物菌、γ-聚谷氨酸、腐殖酸的新型肥料在小麦生产上的应用[J].江苏农业科学,2014,42(12):96-98.
[4] 周霞萍,邓 林,韩媛媛,等.腐植酸天然植物生长素调节剂[J].中国生物防治,2006,22(增刊):163-170.
[5] 李仲谨,李铭杰,王海峰,等.腐植酸类物质应用研究进展[J].化学研究,2009,20(4):103-107.
[6] BAR BULENT A K,MURAT A T,HAKAN E,et al.盐胁迫环境下腐植酸类物质对小麦生长及矿物营养吸收的影响[J].腐植酸,2009(6):30-35.
[7] 任学军,杜 彬,任艳军,等.腐植酸钠对镉胁迫小麦幼苗的生物效应[J].南方农业学报,2011,42(10):1233-1237.
[8] 郭 伟,于立河.腐植酸浸种对盐胁迫下小麦萌发种子及幼苗生理特性的影响[J].麦类作物学报,2012,32(1):90-96.
[9] 闫军营,裴瑞杰.喷施叶面肥对灌溉区小麦光合特性和产量的影响[J]. 安徽农业科学,2014,42(10):2872-2873,2876.
[10]陈玉玲.腐植酸对植物生理活动的影响[J].植物学通报,2000,17(1):11-16.
[11]白志平.腐植酸在农药中的应用及发展前景[J].生物灾害科学,2012(2):6.
[12]赵长星,马东辉,王月福,等.施氮量和花后土壤含水量对小麦旗叶衰老及粒重的影响[J].应用生态学报,2008,19(11):2388-2393.
[13]周 玲,王朝辉,李富翠,等.不同产量水平旱地冬小麦品种干物质累积和转移的差异分析[J].生态学报,2012,32(13):4123-4131.
[14]张 宾,冯佰利,韩媛芬,等.干旱条件下冷型小麦叶片衰老特性研究[J].干旱地区农业研究,2003,21(3):70-73.
[15]尚玉磊,李春喜.生长调节剂对小麦旗叶衰老和产量性状的影响[J].麦类作物学报,2001,21(2):72-75.
[16]周广生,梅方竹.冬小麦根系活力与产量性状关系的研究[J]. 华中农业大学学报,2001,20(6):531-534.
[17]周照留,赵 平,汤 利,等.小麦蚕豆间作对作物根系活力、蚕豆根瘤生长的影响[J].云南农业大学学报,2007,22(5):665-671.
[18]李燕婷,李秀英,肖 艳,等.叶面肥的营养机理及应用研究进展[J].中国农业科学,2009,42(1):162-172.
[19]周吉红,曹海军,朱青兰,等.不同类型叶面肥在不同时期喷施对小麦产量的影响[J].作物杂志,2012(5):140-145.
[20] SIDDIQUE M R B,HAMID A,ISLAM M S.Drought stress effects on photosynthetic rate and leaf gas exchange of wheat[J].Botanical Bulletin of Academia Sinica,1999,40:141-145.
[21]房世波,齐 月,韩国军,等.1961-2010年中国主要麦区冬春气象干旱趋势及其可能影响[J].中国农业科学,2014,47(9):1754-1763.
[22]李迎春,张超英,庞启华,等.干旱胁迫下小麦在不同生育时期的耐旱性研究[J].西南农业学报,2008,21(3):621-624.
[23] SU Z,ZHANG J,WU W,et al.Effects of conservation tillage practices on winter wheat water-use efficiency and crop yield on the Loess Plateau,China[J].Agricultural Water Management,2007,87(3):307-314.
[24] JIANG X D,WANG Y,HOU L T,et al.Effects of minimum tillage and no-tillage systems on photosynthetic characteristics at late growth stages of winter wheat[J].Trans CSAE,2006,22:66-69.
[25] LI Y J,WU J Z,HUANG M,et al.Effects of different tillage systems on photosynthesis characteristics of flag leaf and water use efficiency in winter wheat[J].Trans CSAE,2006,22:44-48.
[26] FABRIZZI K P,GARCIA F O,COSTA J L,et al.Soil water dynamics,physical properties and corn and wheat responses to minimum and no-tillage systems in the southern Pampas of Argentina[J].Soil and Tillage Research,2005,81(1):57-69.
[27]山 仑,陈培元.旱地农业生理生态基础[M].北京:科学出版社,1998.
[28]宗学凤,张建奎,王三根.蓝、紫粒小麦光合特性的研究[J].中国生态农业学报,2008,16(6):1423-1428.
[29]武志海,赵国臣,徐克章,等.吉林省过去47年来水稻品种遗传改良过程中叶片光合指标的变化[J].中国水稻科学,2009,23(2):165-171.
[30]翟虎渠,李良璧.超高产杂交稻灌浆期光合功能与产量的关系[J].中国科学:C 辑,2002,32(3):211-217.
[31]曹树青,杨国南.鉴定水稻灌浆期叶片光合作用与籽粒灌浆切合程度的新指标[J].作物学报,2002,28(4):516-520.
[32]党建友,姬虎太.冬小麦喷施叶面肥增产效应分析[J].小麦研究,2000,21(1):17-18.
[33]裴雪霞,王姣爱,党建友,等.后期喷肥对强筋小麦临汾138产量和品质的影响[J].麦类作物学报,2006,25(6):148-149.
[34] SHAABAN S H A,MANAL F M,AFIFI M H M.Humic acid foliar application to minimize soil applied fertilization of surface-irrigated wheat[J].World Journal of Agricultural Sciences,2009,5(2):207-210.
[35] SON N T,THANG M N,DAN N K,et al.The effect of spray humic acid on leaves to growth,development,yield and quality of rice of C70 variety[J].Nong Nghiep Va Phat Trien Nong Thon,2004,4:485-485,493.
[36]李安民,陈绍荣,卢燕林.腐植酸在作物生长发育化学控制中的作用及机理探讨[J].腐植酸,2007(4):15-22.
[37] WANG J L,YU G R,WANG B L,et al.Response of photosynthetic rate and stomatal conductance of rice to light intentity and CO2concentration in northern China[J].Chinese Journal of Plant Ecology,2005,29(1):16-25.
[38] FENG Z Z,WANG X K,DUAN X N,et al.Effect of nitrogen fertilization on diurnal changes of flag leaf photosynthetic rate in spring wheat[J].Chinese Journal of Ecology,2003,22(4):90-92.
[39]刘建辉,孙建云,戴廷波,等.不同小麦进化材料生育后期光合特性和产量[J].植物生态学报,2007,31(1):138-144.
[40]张 国,李 滨,邹 琦.小麦 Rubisco活化酶基因的克隆和表达特性[J].植物学通报,2005,22(3):313-319.
[41]许晓明,陆 巍,张荣铣,等.超高产水稻协优9308的高效光合功能[J].南京师大学报:自然科学版,2004,27(1):78-81.
[42] ZHANG D Y,CHEN G Y,GONG Z Y.Ribulose-1,5-bisphosphate regeneration limitation in rice leaf photosynthetic acclimation to elevated CO2[J].Plant Sci,2008,175:348-355.
[43]李卫芳,王秀海,王 忠.小麦旗叶Rubisco和Rubisco活化酶与光合作用日变化的关系[J].安徽农业大学学报,2006,33(1):30-34.
[44]张 宾,冯佰利,韩媛芬,等.干旱条件下冷型小麦叶片衰老特性研究[J].干旱地区农业研究,2003,21(1):70-73.
[45]山东农学院.作物栽培学(北方本,上册)[M].北京:农业出版社,1990:87-89,148.
[46] VALKOUN J.Wheat pre-breeding using wild progenitors[M].Netherlands:Springer,2001:699-707.
[47]赵 慧,张正斌,徐 萍.小麦叶片水分利用效率生理性状遗传相关分析[J].中国农业科学,2006,39(9):1796-1803.
[48]王瑞霞,张秀英,伍 玲,等.不同生态环境条件下小麦籽粒灌浆速率及千粒重QTL分析[J].作物学报,2008,34(10):1750-1756.
[49]曹树青,赵永强,温家立,等.高产小麦旗叶光合作用及与籽粒灌浆进程关系的研究[J].中国农业科学,2000,33(6):19-25.
