不同氮磷比条件下结缕草克隆整合作用的生长代价与收益
2015-01-01陈惠娟李德志赖苏雯贠小涛陈青青黄瑶瑶樊治华欧阳义王明祖
陈惠娟, 李德志,2,3, 赖苏雯, 贠小涛, 耿 松, 陈青青, 黄瑶瑶, 樊治华,欧阳义, 王明祖
(1.华东师范大学生态与环境科学学院,上海 200241;2.华东师范大学上海市城市化生态过程与生态恢复重点实验室,上海200241;3.华东师范大学浙江天童森林生态系统国家野外科学观测研究站,上海 200241)
克隆整合或生理整合是克隆植物适应复杂生境的重要生态对策,主要是指碳水化合物、矿质养分、水分、激素、酶和病原菌等资源或物质通过连接物或间隔子在克隆分株之间转移与分享的过程[1-3]。大量研究结果显示,克隆整合有利于克隆幼小分株的建立[4],缓解干旱[5]、水淹[6]、风蚀[7]、沙埋[8]、高盐[9]、采食[10]、低养和弱光[11]等不利环境因素对克隆植株的干扰或胁迫,增强克隆植株对异质性生境的适应和资源利用能力[8-9,12],从而提高克隆植株的生长收益。然而,资源的转移和匍匐茎的生长需要消耗能量,并且资源稀释容易导致较高的死亡风险,克隆植株为维持克隆整合必然要付出代价[13]。但这一代价的大小和相关影响因素,以及在不同环境条件下,整合时间对克隆植物生长代价的影响如何、通过什么方式可以达到对于整合代价的最佳释放,迄今尚未有人进行过深入的研究和揭示。此外,研究克隆植物的整合作用所产生的收益和代价,对深入理解克隆整合机制和剖析克隆植物复杂生态行为具有重要科学价值。
结缕草(Zoysia japonica Steud.)属于禾本科,结缕草属,多年生匍匐茎型克隆植物[14-15]。结缕草的抗逆性和环境适应能力强,常用于城市各类草坪的种植,同时也是优质牧草。因而,结缕草具有较高的经济和生态价值,其克隆生长习性和克隆整合特性也日益受到生态学家们的关注[16-17]。近年来,许多学者采用化学示踪方法对结缕草克隆植物体内的水分、碳素和氮素等养分资源的传输方向、整合格局与过程等方面进行了研究,进一步证实了结缕草克隆分株之间存在着广泛的生理整合作用[3,18-19]。然而,结缕草克隆分株之间进行养分资源整合过程中是否同时存在收益与代价,目前尚未见报道。
克隆植株的克隆整合特性和生长状况与生境中的养分水平及主要营养元素的比例密切相关[16,20]。李立科等[19]通过对结缕草进行15N 示踪研究发现,在高浓度的15N营养生境中植株倾向将更多的15N分配给匍匐茎以便进行远距离传输,在低浓度的15N营养生境中植株则优先满足A、B分株的生长需要。32P在大、小叶两种基因型的白三叶植株体内的传输策略及分配格局也受到生境中32P营养状况的影响[21]。在不同土壤氮磷肥力条件下,短穗兔儿草的克隆生长特性及生长状况表现出明显的差异[22]。此外,氮和磷也是植物生长发育过程中需求量较高的主要营养元素,参与植物体内多种生理代谢过程,也常常是植物生长发育的重要限制因子[23-24]。因此,在氮、磷比例不同的生境中,结缕草的生长状况和克隆整合作用所产生的代价与收益可能存在差异。本试验设置氮磷比不同的3种养分水平的土壤条件,并对结缕草克隆植株匍匐茎实施4种切断处理,以此研究在不同氮磷比土壤条件下,不同时间长度的克隆整合作用对结缕草植株生长和生物量积累的影响,并进行生长代价与收益的分析,以深入了解结缕草克隆整合机制及其对土壤养分资源适应策略,为结缕草草坪培育、维护和养分管理提供科学依据。
1 材料与方法
1.1 材料培养
试验材料为华东师范大学上海市城市化生态过程与生态恢复重点实验室培育的结缕草。其基本组成单位是复合节(图1),A、B分株或分蘖,交错着生在复合节的两侧[3,17]。选择形态完整的结缕草克隆片段,切断匍匐茎,把得到的单个分株,种植在花盆中培育。约30 d后,大部分结缕草分株会形成第1个复合节。从中挑选大小、根系和形态特征基本一致,且长出第1个复合节的植株作为试验材料。把试验材料移植到塑料管(直径为2.5 cm、高15.0 cm)中,作为试验母株。在新形成的主匍匐茎每个复合节下方,设置另一个相同型号的塑料管,让新复合节在其中扎根,并形成第一个分株。在试验过程中,每长出一个复合节就增设一个塑料管。培养基质均为用清水漂洗过的河沙。

图1 结缕草克隆植株的典型形态结构示意图Fig.1 The diagram of typical morphology of Zoysia japonica cloned plant
1.2 试验设计
设置3种氮磷配比的营养液。在Hoagland溶液配方的基础上,通过修改磷酸二氢钾含量,配制3种氮磷比的营养溶液,减少的钾用氯化钾补充。3种氮磷配比的营养液为:(1)低氮磷比营养液(稀释10%的Hoagland溶液,N∶P2O5=7∶1);(2)中氮磷比营养液(N∶P2O5=14∶1);(3)高氮磷比营养液(N∶P2O5=21∶1)。使用时,所有营养液均稀释10倍。在分株生长过程中,对每个复合节已扎根的塑料管每7 d施加6 ml相应氮磷配比的营养液。
在每种养分水平的生境条件下,对匍匐茎实施4种处理(即4种整合水平):连接(不切断匍匐茎)、轻度切断、中度切断和重度切断。每种处理3个重复。3种切断处理的方法:分别在结缕草克隆植株长出12、8和4个复合节后,切断母株与第1个复合节之间的匍匐茎,之后每长出1个新复合节,就依次将位于基部的复合节与其他复合节间的连接关系切断,使匍匐茎上连接的复合节分别保持12、8和4个。
试验从2013年7月持续至2013年11月。所有试验材料均放在光照培养箱(GXZ智能型)中培养,设置白天12 h(光照度为12 000 lx)、28℃;夜晚12 h(无光)、20℃。试验期间正常浇水管理,并观察记录分株生长情况。
1.3 数据收集和处理
试验结束后,收获植株。收获时,用清水洗去塑料管中的沙粒,避免破坏根系、叶和匍匐茎的完整性。用毛巾吸干整个分株上的水珠,测量匍匐茎每个节间的长度,并记录一级和二级A、B分株或分枝数量。然后,将植株放入烘箱,于70℃下烘干至恒质量。分别称量植株的A分枝、B分枝、匍匐茎和根的干质量。以切断程度、氮磷比为因子,测定的各生长指标为因变量,采用SPSS 20.0软件,进行多因素方差分析;在方差分析结果达到显著时,采用LSD、Duncan法对不同处理下的数据进行多重比较。
2 结果与分析
2.1 氮磷比和切断处理条件下结缕草克隆植株的形态指标变化
随着对匍匐茎切断程度的加大,整个克隆植株的主匍匐茎总长度、主匍匐茎复合节数和一级A分株数呈下降趋势,且匍匐茎连接处理与其他3种切断处理间差异显著(表1、图2)。低氮磷比养分条件下生长的克隆植株主匍匐茎总长度显著高于中、高氮磷比养分条件,但主匍匐茎复合节数和一级A分株数在各氮磷比养分处理间差异不显著(表1)。
对结缕草匍匐茎实施切断处理、土壤氮磷比处理及两者间的交互作用对结缕草克隆植株的一级B分株数、和二级A、B分枝数均有极显著的影响(表1)。与匍匐茎保持连接相比,无论对匍匐茎实施何种程度的切断处理,结缕草克隆植株上一级B分株数和二级A、B分枝数都显著上升(图2)。低氮磷比条件下生长的结缕草克隆植株上一级B分株数显著高于中、高氮磷比养分条件,并且在低氮磷比养分条件下对匍匐茎实施轻度切断处理时,一级B分株数和二级A、B分枝数的值最大。总体而言,结缕草克隆植株在低氮磷比养分条件下具有明显的生长优势。长期的克隆整合有利于主匍匐茎的伸长、主匍匐茎复合节的形成和一级A分株的建立,但不利于一级B分株的建立和二级A、B分枝的形成。
2.2 氮磷比和切断处理条件下结缕草克隆植株的生长代价与收益分析
结缕草匍匐茎切断处理、土壤氮磷比水平处理及两者间的交互作用对结缕草克隆植株的母株生物量有极显著的影响(表1)。对匍匐茎实施不同程度的切断均使克隆植物的母株生物量显著增加,尤其在高氮磷比条件下,随着匍匐茎切断程度由轻到重,克隆植株的母株生物量呈显著的上升趋势;在中、低氮磷比养分条件下,分别对匍匐茎实施轻度切断和中度切断时,克隆植株的母株生物量显著高于同种氮磷比条件下的其他切断处理(图3)。
结缕草克隆植株在低氮磷比条件下积累的总生物量显著高于中、高氮磷比条件(表1、图3),进一步证明其在低氮磷比生境中生长更为适宜。对匍匐茎实施重度切断处理时,结缕草克隆植株的总生物量和根系生物量都显著低于其他3种处理(即连接、轻度和中度切断处理),且在3种氮磷比养分条件下均有相同的趋势。在高氮磷比养分条件下,随着对匍匐茎的切断程度从轻到重,克隆植株的根系生物量呈显著下降趋势,植株总生物量也表现出相似的规律;在中氮磷比养分条件下,与匍匐茎连接相比,克隆植株的总生物量和根系生物量反而在轻度切断处理下分别显著上升30.6%和23.9%;而在低氮磷比养分条件下,与匍匐茎连接相比,克隆植株的总生物量和根系生物量分别在中度和轻度切断处理时显著上升,呈现显著的交互效应(表1)。在任何氮磷比养分条件下,分别对主匍匐茎实施中度切断、轻度切断和不切断(连接)处理时,其克隆植株总生物量分别为3.65 g、2.90 g和2.94 g,均高于重度切断处理。可见,结缕草母株和克隆分株之间保持较长时间的较紧密的生理整合更有利于结缕草克隆植株根系生物量和总生物量积累。
低氮磷比养分条件中生长的结缕草克隆植株上主匍匐茎生物量和A分枝生物量都显著高于中、高氮磷比养分条件(表1、图3);然而,无论在何种氮磷比条件下,随着克隆植株匍匐茎切断程度由轻到重,主匍匐茎本身的生物量都呈显著下降趋势。这再次证明主匍匐茎的生长与克隆整合密切相关,而在不同生长阶段中断整合(切断)都损害匍匐茎的生物量积累。与匍匐茎保持连接相比,对匍匐茎实施轻度和中度切断处理时,A分枝生物量显著增加(表1、图3),而对匍匐茎实施重度切断处理时,A分枝生物量则显著下降。值得注意的是,从轻到重3种切断处理,均使B分枝生物量显著增加。但在中、低氮磷比养分条件下,重度切断处理下的B分枝生物量显著低于轻度和中度切断处理(图3)。可见,对匍匐茎实施适当的切断处理,使A、B分枝的生长显著受益,然而,过重切断匍匐茎也会损害A、B分枝的生长。在低氮磷比条件下,对匍匐茎实施中度切断处理时,结缕草克隆植株上A、B分枝生物量最高,表现出显著的交互效应(表1)。

图2 3种氮磷比水平和4种切断处理条件下结缕草的主匍匐茎总长度(A)、主匍匐茎复合节数(B)、一级A分株数(C)、一级B分株数(D)、二级A分枝数(E)和二级B分枝数(F)的变化Fig.2 Total primary stolon length(A),number of primary multiple nodes(B),number of primary A-ramets(C),number of primary B-ramets(D),number of secondary A-branches(E)and number of secondary B-branches(F)of Z.japonica plants in the treatments of stolon severings and N∶P application ratios
2.3 几种氮磷比和切断处理条件下结缕草克隆植株的生物量分配变化特征
与匍匐茎连接相比,无论采取何种切断处理,结缕草克隆植株对A、B分枝生物量的分配都显著增加,而对匍匐茎生物量的分配却显著下降,对根系的生物量分配则变化不显著,且在3种氮磷比养分生境中均有相似的规律(表1、图4)。值得注意的是,结缕草克隆植株对A分枝的投资 (25.7%~47.6%)始终高于对B分枝的投资 (3.9%~16.5%)。然而,随着生境中氮磷比的升高,结缕草克隆植株对根系生物量的分配提高(图4),即低氮磷比养分生境下根系生物量分配比 (31.9%~33.5%)<中氮磷比养分生境下根系生物量分配比(33.2%~36.8%)<高氮磷比养分生境下根系生物量分配比(34.8% ~38.3%),且低氮磷比与高氮磷比处理间差异显著(P<0.05)(表1)。
3 讨论
本研究发现,无论在何种氮磷比生境,当匍匐茎保持完全连接状态时,长期的克隆整合显著降低了母株生物量。这可能是由于母株通过匍匐茎向年幼分株传输养分时,资源输出导致内部资源稀释,因而通常要承担一定程度的生长代价[13]。
大量研究结果表明,克隆整合对克隆新生分株的建立和生长有显著的促进作用[11,25-26];也有研究者认为,匍匐茎的断开能够促进克隆植株的生长、繁殖和扩散[16,27]。本研究发现,随着匍匐茎切断程度由轻到重,结缕草克隆植株主匍匐茎总长度及其生物量、主匍匐茎复合节数和一级A分株数呈下降趋势,这可能与阻断养分资源向顶端运输密切有关,这表明克隆整合对结缕草的向顶生长具有重要作用;然而,对匍匐茎实施不同程度的切断处理,显著增加结缕草克隆植株的一级B分株数、二级A、B分枝数和B分枝生物量。由于在结缕草克隆植株的同一复合节上,A、B分株交互生长,A分株靠近克隆植株的基端,而B分株更靠近顶端[17],因此,B分株的生长可能更易受到匍匐茎顶端优势的影响。而对匍匐茎实施切断处理后,从匍匐茎顶端向被切断部分的信号传导被阻断[28],结缕草克隆植株的顶端优势在一定程度被释放,这有利于原本受到抑制的克隆分株的迅速生长和发育,B分株的表现尤为突出,不仅生长迅速,有的甚至可以成为一级B分枝。同时,匍匐茎的连接是克隆植物进行生理整合的前提,切断匍匐茎就会中断养分资源在母株和克隆分株之间的转移或分享,匍匐茎对资源的消耗也相应减少。在此情况下,克隆分株可能将自身吸收或合成的资源用来投资生长二级A、B分株或分枝,而增加二级分枝强度有利于克隆植株实现生境空间的拓展和占据,以此也有利于克隆植株寻找新的生机。
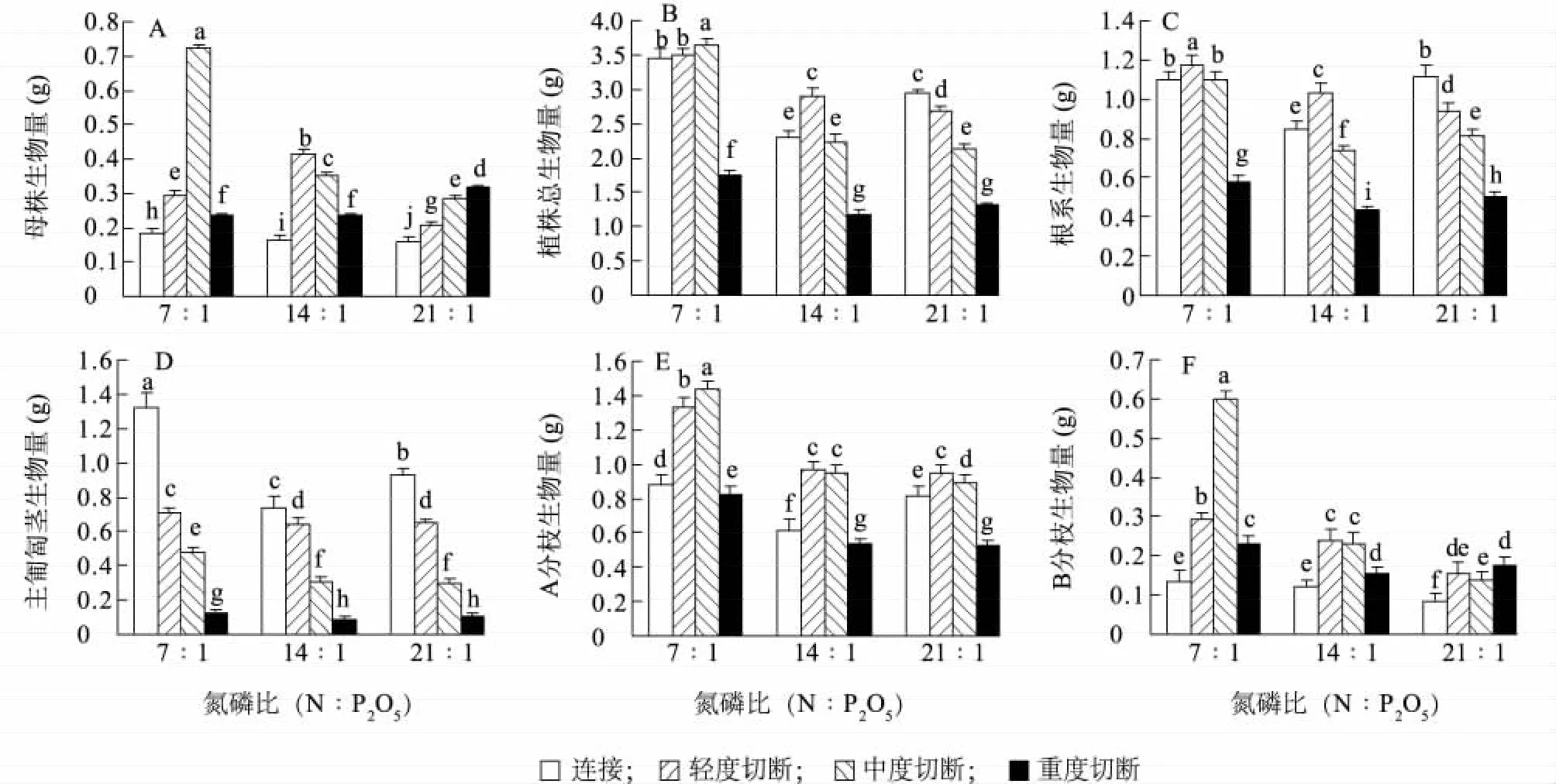
图3 3种氮磷比水平和4种切断处理条件下结缕草的母株生物量(A)、植株总生物量(B)、根系生物量(C)、匍匐茎生物量(D)、A分枝生物量(E)和B分枝生物量(F)的变化Fig.3 Biomass of parent ramet(A),total biomass of plant(B),root biomass(C),stolon biomass(D),A-branch biomass(E)and B-branch biomass(F)of Z.japonica in the treatments of stolon severings and N∶P application ratios
然而,在重度切断处理条件下的一级B分株数、二级A、B分枝数、A、B分枝生物量和植株总生物量均显著低于轻度切断处理或中度切断处理。由于过早中断克隆整合,结缕草植株向顶生长严重受限,在一定程度上可能也会影响克隆植株的侧向生长。因此,在结缕草克隆生长过程中,宜对匍匐茎采取适当的切断处理,有利于结缕草克隆植株释放生长潜能和增加二级分枝数量,从而能够高效积累生物量。
克隆整合显著改变生物量分配格局,这在很多研究中得到证实[25-26]。与匍匐茎连接相比,结缕草克隆植株对A、B分枝生物量分配在3种切断处理下显著增加,而对匍匐茎的生物量分配却恰好相反。这可能也是适当切断能够增加结缕草克隆植株分枝数量和生物量的重要因素。由于切断匍匐茎减少了其对资源的消耗[13],同时也阻断了养分的传输,克隆植株将更多的资源分配至其地上的分枝部分,以获取充足的光照和形成更多的光合同化物。值得注意的是,不论在何种切断处理条件下,A分枝生物量分配始终大于B分枝生物量分配,结缕草对水分资源的分配也有相似的现象[3],这进一步验证了结缕草同一复合节上A分枝的生长速率始终大于并优先于B分枝的趋势的不变性[17]。
与中、高氮磷比养分生境相比,结缕草克隆植株在低氮磷比养分生境中长势最好,这可能受磷含量下降和氮磷比例的影响。在本试验中,3种营养液中磷含量随着氮磷比的升高而下降。生境中的氮磷配比可能也会影响植物对氮、磷的吸收效率而影响生长[29]。户桂敏等[30]发现,生境中磷水平的限制会影响五爪金龙植物对氮的吸收。然而,随着生境土壤氮磷比的升高,结缕草克隆植株对根系生物量的分配增加,这可能是由于在养分资源贫瘠或利用效率低的生境中,克隆植物倾向把更多的养分资源分配给根系,做出积极的补偿,保证其能够获取生长所需的养分或水分[31]。同时,随着生境氮磷比的升高,结缕草母株和克隆分株之间保持较长时间的较紧密的生理整合更有利于结缕草克隆植株生长和生物量积累。可见,结缕草克隆整合策略与生境氮磷比密切相关。
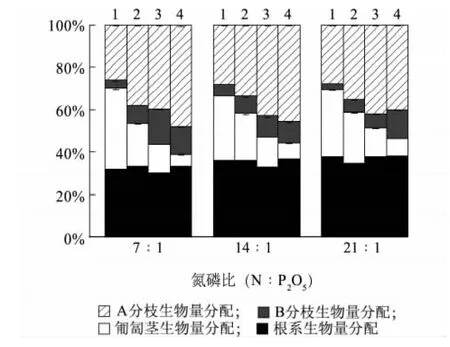
图4 3种氮磷比水平和4种切断处理下结缕草植株对A分枝、B分枝、匍匐茎和根系的生物量分配Fig.4 Proportional biomass allocation to A-branches,B-branches,stolons and roots of Z.japonica plants in the treatments of stolon severings and N∶P application ratios
综上所述,结缕草克隆植株克隆整合的代价与收益同时存在。结缕草在低氮磷比条件下生长最好,适时中断克隆整合可以释放结缕草克隆植株的生长潜能,减少匍匐茎对植物体内养分资源的消耗,增加二级A、B分枝数量和对A、B分枝生物量分配,从而有利于克隆植株实现对生境空间的占据,以获取更多养分资源,提高自身生长收益。因此,在低氮磷比条件下,对匍匐茎实施中度切断处理时,结缕草植株总生物量收益最好。本研究结果反映了结缕草适应不同氮磷比生境的复杂生态策略,对于结缕草草坪的科学管理也具有一定的应用价值。
[1] LAM C K,BELANGER F C,WHITE J F,et al.Invertase activity in Epichloe acermonium fungal endophytes and its possible role in choke disease[J].Mycological Research,1995,99(7):867-873.
[2] ALPERT P.Clonal integration in Fragaria chiloensis differs between populations:ramets from grassland are selfish[J].Oecologia,1999,120(1):69-76.
[3] 程立丽,李德志,朱志玲,等.克隆植物结缕草的水分生理整合格局特征及其生态效应分析[J].西北植物学报,2013,33(7):1442-1451.
[4] ROILOA S R,ANTELO B,RETUERTO R.Physiological integration modifies δ15N in the clonal plant Fragaria vesca,suggesting preferential transport of nitrogen to water-stressed offspring[J].Annals of Botany,2014,114(2):399-411.
[5] ZHANG Y,ZHANG Q,LUO P,et al.Photosynthetic response of Fragaria orientalis in different water contrast clonal integration[J].Ecological Research,2009,24(3):617-625.
[6] XIAO Y,TANG J,QING H,et al.Clonal integration enhances flood tolerance of Spartina alterniflora daughter ramets[J].Aquatic Botany,2010,92(1):9-13.
[7] YU F H,WANG N,HE W M,et al.Adaptation of rhizome connections in drylands:increasing tolerance of clones to wind erosion[J].Annals of Botany,2008,102(4):571-577.
[8] CHEN J,LEI N,DONG M.Clonal integration improves the tolerance of Carex praeclara to sand burial by compensatory response[J].Acta Oecologica,2010,36(1):23-28.
[9] 廖咏梅,刘富俊,陈劲松.克隆整合对异质性盐分胁迫下积雪草生长的影响[J].植物分类与资源学报,2013,35(1):55-61.
[10] LIU H,YU F,HE W,et al.Clonal integration improves compensatory growth in heavily grazed ramet populations of two inlanddune grasses[J].Flora-Morphology,Distribution,Functional E-cology of Plants,2009,204(4):298-305.
[11] ROILOA S R.Responses of the clonal Fragaria vesca to microtopographic heterogeneity under different water and light conditions[J].Environmental and Experimental Botany,2007,61(1):1-9.
[12]罗 栋,钱永强,刘俊祥,等.克隆植物野牛草对异质营养的表型可塑性响应[J].草业学报,2014,23(3):104-109.
[13] PITELKA L F,ASHMUN J.Physiology and integration of ramets in clonal plants[M].New Haven:Yale University Press,1985:399-435.
[14]张 勇,吕爱芹,吴翠霞,等.15%硝磺草酮防除草坪(早熟禾)杂草效果及其安全性[J].杂草科学,2013,31(4):54-56.
[15]张绍明,张开龙,强 胜.江苏省主要农田杂草调查方法探讨[J]. 杂草科学,2014,32(4):24-27.
[16] LI D Z H,TAKAHASHI S,ZHI T C.Effects of soil nitrogen availability and clonal integration on the branching behaviors of Zoysia japonica[J].Acta Prataculturae Sinica,2006,15(5):135-146.
[17]李德志,申芳芳,王长爱,等.克隆植物结缕草在两种环境中的生长发育特征[J].华东师范大学学报:自然科学版,2006(6):117-124.
[18]何莹莹,李德志,徐 洁,等.不同光照条件下结缕草全株碳素生理整合格局及其生态效应[J].植物资源与环境学报,2013,22(4):1-10.
[19]李立科,李德志,赵鲁青,等.克隆植物结缕草主匍匐茎中氮素的传输特征[J].东北林业大学学报,2011,39(11):40-45.
[20] HE W M,YU F,ZHANG L.Physiological integration impacts nutrient use and stoichiometry in three clonal plants under heterogeneous habitats[J].Ecological Research,2010,25(5):967-972.
[21] LOTSCHER M,HAY M.Genotypic differences in physiological integration,morphological plasticity and utilization of phosphorus induced by variation in phosphate supply in Trifolium repens.[J].The Journal of Ecology,1997,85(3):341-350.
[22]杨 帆,周华坤,叶 鑫,等.不同氮磷组合对短穗兔儿草克隆繁殖特性的影响[J].草原与草坪,2010,30(4):64-67.
[23] GÜSEWELL S.N∶P ratios in terrestrial plants:variation and functional significance[J].New Phytologist,2004,164(2):243-266.
[24]宁运旺,马洪波,张 辉,等.氮、磷、钾对甘薯生长前期根系形态和植株内源激素含量的影响[J].江苏农业学报,2013,29(6):1326-1332.
[25]王 微.克隆整合对结缕草与狗牙根种间竞争的影响研究[J].湖北农业科学,2012,51(9):1833-1836.
[26] YOU W H,YU D,XIE D,et al.The invasive plant Alternanthera philoxeroides benefits from clonal integration in response to defoliation[J].Flora-Morphology,Distribution,Functional Ecology of Plants,2014,209(11):666-673.
[27]滕小华,洪锐民,乌云娜.克隆植物的裂断、空间遗传结构的研究进展[J].东北农业大学学报,2011,42(7):8-14.
[28] MASCHINSKIAND J,WHITHAM T G.The continuum of plant responses to herbivory:the Influence of plant association,nutrient availability,and timing[J].The American Naturalist,1989,134(1):1-9.
[29] GÜSEWELL S,BOLLENS U.Composition of plant species mixtures grown at various N∶P ratios and levels of nutrient supply[J].Basic and Applied Ecology,2003,4(5):453-466.
[30]户桂敏,王文天,彭少麟.不同氮磷比下入侵种五爪金龙和本地种鸭脚木的竞争表现[J].生态环境学报,2009,18(4):1449-1454.
[31]马俊改,石福臣.养分条件对互花米草表型可塑性的影响[J].生态学杂志,2011,30(3):459-463.
