小麦产量构成因素的基因效应和杂种优势分析
2015-01-01姚金保周淼平马鸿翔杨学明
姚金保, 杨 丹, 周淼平, 马鸿翔, 杨学明
(江苏省农业科学院/江苏省农业生物学重点实验室,江苏 南京 210014)
小麦籽粒产量由单位面积穗数、每穗粒数和千 粒质量3个因素构成[1-3]。育种实践表明,在保证足够穗数的基础上,增加穗粒数和稳定提高千粒质量是今后小麦产量突破的重要途径[4-5]。近年来,许多学者采用不同方法对小麦每穗粒数和千粒质量的遗传特性和杂种优势进行了研究。一般认为小麦每穗粒数和千粒质量的遗传符合加性-显性模型,基因作用方式以加性效应为主[6-10],也有以显性效应为主的报道[11-14]。也有研究者认为小麦每穗粒数或千粒质量的遗传机制复杂,可能存在上位性效应[15-17]。数量遗传研究结果表明,小麦的每穗粒数和千粒质量是质量-数量性状,由主基因和微效基因共同控制[18]。关于小麦每穗粒数和千粒质量杂种优势的研究结论也不完全一致。多数研究结果表明,小麦每穗粒数和千粒质量的平均杂种优势在F1代普遍存在一定的正向优势[19-22]。Inamullah等[23]、Al-Falahy[24]研究发现,每穗粒数和千粒质量的平均杂种优势变异幅度较大,其优势强弱因组合而异。许永财等[25]研究认为,千粒质量具有较高的正向超亲优势,但每穗粒数表现负向中亲优势。赵鹏等[26]、史秀秀等[27]的研究结果也证实了每穗粒数表现为负向中亲优势。但 Abdullah等[28]、Ilker等[29]认为,每穗粒数存在明显的正向超亲优势。由此可见,对于小麦产量构成因素的遗传特性和杂种优势还需进一步研究。本试验以7个产量构成因素差异较大的小麦品种为亲本,按n(n-1)/2双列杂交法配制21个组合,在2种环境下研究每穗粒数和千粒质量2个小麦产量构成因素的基因效应和杂种优势,旨在为小麦高产育种提供理论依据。
1 材料与方法
1.1 材料
2012年春选用综合农艺性状优良、每穗粒数和千粒质量有一定差异的7个冬小麦品种作为亲本,7个品种为徐麦25(P1)、矮抗58(P2)、淮麦20(P3)、淮麦33(P4)、烟 2801(P5)、济麦 22(P6),徐麦 31(P7)。
1.2 试验设计
按Griffing双列杂交模型II,配制成21个杂交组合,2013年秋分别在宿迁和南京种植F1及亲本,共28个基因型。田间按随机区组排列,重复3次,每小区2行区,行长1.5 m,行距25 cm,株距4 cm,田间管理措施同大田栽培。
1.3 性状调查
2014年在小麦成熟前,每小区随机取30穗,调查每穗粒数;成熟时按小区收获,脱粒后扬净、风干,分小区统计千粒质量。
1.4 数据统计分析
杂种优势测定:平均优势(%)=[(F1-MP)/MP]×100%,超高亲优势(%)=[(F1-HP)/HP]×100%,超低亲优势(%)=[(F1-LP)/LP]×100%,其中Fl为杂种一代性状平均值,MP、HP和LP分别表示双亲均值、高值亲本值和低值亲本值。利用Hayman双列杂交方法,对2个产量构成因素分试点进行遗传模型分析并估算遗传参数[30]。
2 结果与分析
2.1 每穗粒数和千粒质量的遗传模型分析
2.1.1 方差分析 7个小麦品种及其双列杂交组配的21个杂交组合的2个产量构成因素在宿迁和南京2个试点上的测定值列于表1。组合间穗粒数在宿迁和南京试点间的变化范围分别为32.07~47.87粒和 28.63~46.40粒,变异系数分别为11.38%和10.51%;组合间千粒质量在宿迁和南京试点间的变化范围分别为 38.03~50.33 g和38.07~48.03 g,变异系数分别为6.87%和6.27%,表明所分析的性状具有较大差异。方差分析结果(表2)表明,每穗粒数和千粒质量基因型间方差均达极显著水平,而重复间差异均不显著,说明基因型效应存在显著差异,即供试材料间的遗传基础存在显著差异,因此可以做进一步的遗传分析。
2.1.2 加性-显性模型测验 每穗粒数系列协方差(Wr)/方差(Vr)回归分析结果(表3)表明,其直线回归系数b与0的差异达显著(宿迁)和极显著(南京),与1的差异均不显著,由此可见每穗粒数的遗传符合加性-显性模型。由回归截距α的测验结果可知,每穗粒数的回归截距α<0,但与0差异不显著,说明该性状属超显性遗传。对南京点试验的千粒质量Wr、Vr进行回归分析,得α=0.048,b=0.541。由回归斜率b的检验结果(表3)可以看出,b与0、1间都存在显著差异,这表明控制千粒质量的基因不仅具有加性、显性作用,还可能存在基因位点间的互作(上位性效应)。应作Wr-Vr和Wr+Vr方差分析,以进一步确定是否存在上位性效应。由表4可知,千粒质量的Wr+Vr、Wr-Vr值在公共亲本间和公共亲本内差异均未达到显著水平,表明不存在上位性效应。宿迁点千粒质量的分析结果(表3)则表明,b与0存在显著差异,而与1差异不显著。千粒质量的基因作用方式为加性、显性作用。综合2个地点的分析结果,认为千粒质量的遗传符合加性-显性模型,不存在上位性效应。
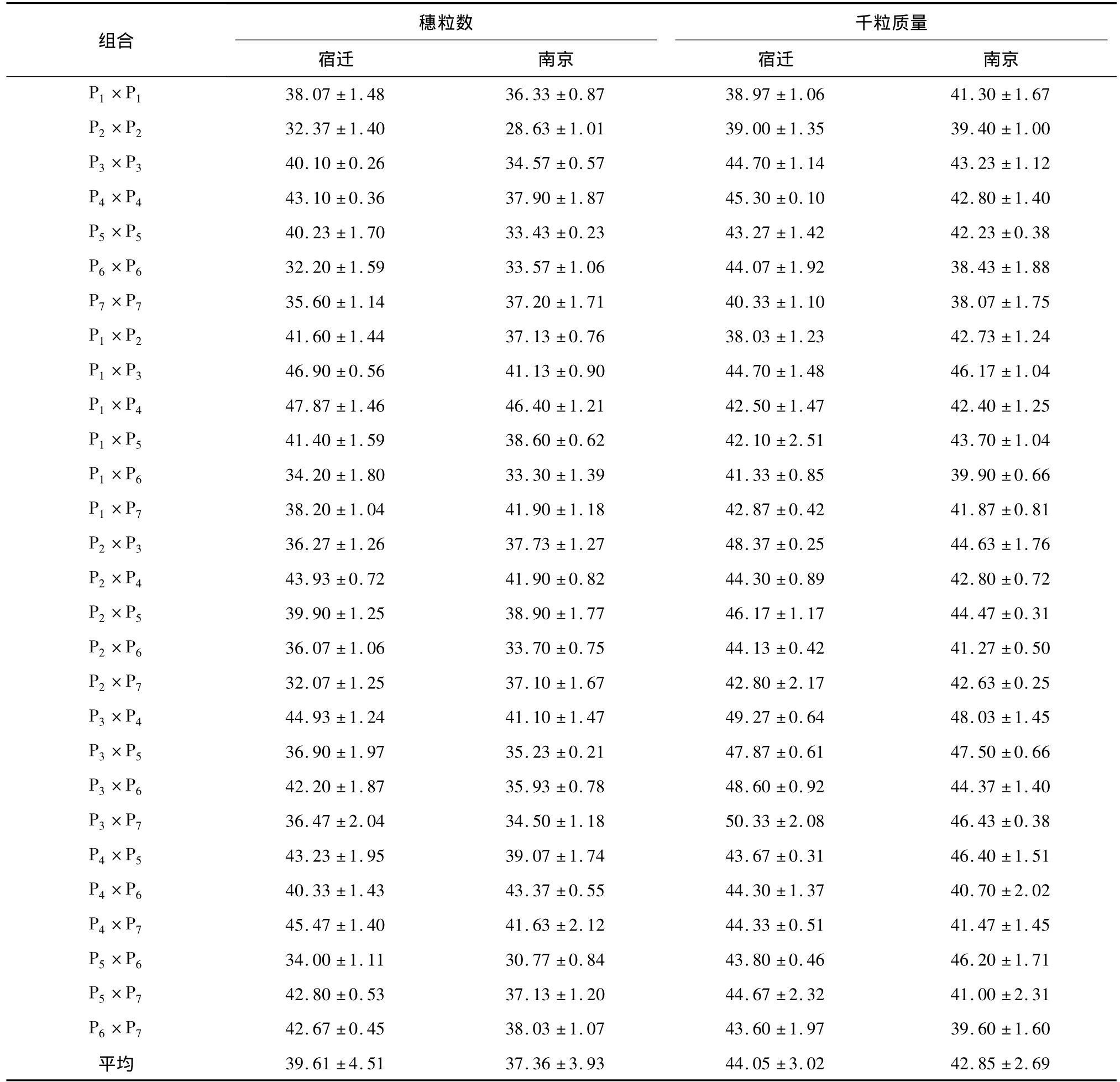
表1 不同组合每穗粒数和千粒质量在2个试点的平均值Table 1 Averages for number of kernels per spike and thousand-grain weight in different crosses at two sites
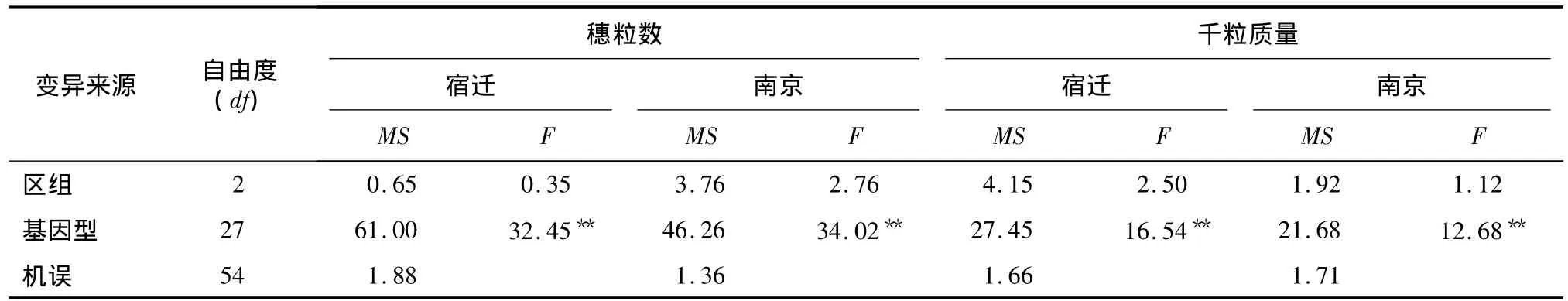
表2 2个试点每穗粒数和千粒质量的方差分析Table 2 ANOVA for kernel number per spike and thousand-grain weight at two sites

表3 每穗粒数和千粒质量系列协方差(Wr)对方差(Vr)的回归分析Table 3 Regression analyses of covariance(Wr)on variance(Vr)for kernel number per spike and thousand-grain weight at two sites
2.1.3 亲本显隐性基因的分布及其作用方向分析 由图1可知,在每穗粒数的遗传上,各亲本具有不等的显隐性等位基因。其中淮麦33、烟2801距原点最近,含有最多的显性基因;徐麦25、矮抗58距原点最远,表明含隐性基因最多。由图2可知,淮麦20具有较多控制千粒质量遗传的显性基因,而其他6个亲本在2个试点的试验中所含显隐性基因比例不完全一致。由表5可知,每穗粒数和千粒质量的亲本均值(Yr)与Wr+Vr的相关系数均呈负相关,表明每穗粒数和千粒质量的遗传表现为高值受显性基因控制,低值受隐性基因控制,显性等位基因对每穗粒数和千粒质量起增效作用,隐性基因则起减效作用。
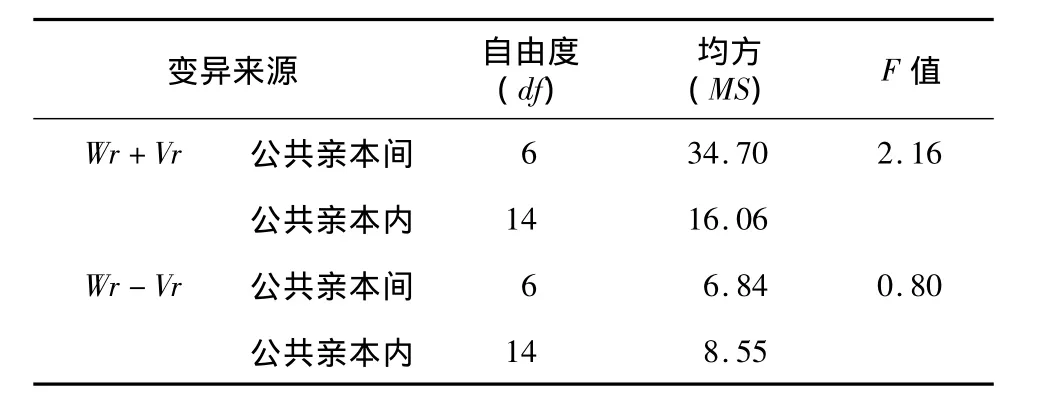
表4 千粒质量Wr+Vr和Wr-Vr的方差分析(南京点)Table 4 ANOVA for Wr+Vrand Wr-Vrof thousand-grain weight in Nanjing experiment

图1 宿迁(A)、南京(B)试点每穗粒数Wr依Vr的回归直线Fig.1 Regression of Wron Vrfor kernel number per spike in Suqian(A)and Nanjing(B)

图2 宿迁(A)、南京(B)试点千粒质量Wr依Vr的回归直线Fig.2 Regression of Wron Vrfor thousand-grain weight in Suqian(A)and Nanjing(B)
2.1.4 遗传参数的估算 表5结果表明,每穗粒数和千粒质量的加性效应方差D和显性效应方差H1、H2均达极显著水平,说明每穗粒数和千粒质量的遗传受加性效应和显性效应共同作用;由于H1远大于D,表明在每穗粒数和千粒质量的遗传中,基因的显性作用较加性作用更重要。每穗粒数的平均显性度分别为1.715(宿迁点)和2.047(南京点),千粒质量的平均显性度分别为1.576(宿迁点)和1.787(南京点),显性程度均为超显性。每穗粒数和千粒质量的H2/4H1值均<0.25,表明亲本中每穗粒数和千粒质量的增、减效等位基因分布不对称;由显性和隐性基因之比[(4DH1)1/2+F]/[(4DH1)1/2-F]均为<1可知,控制亲本的隐性基因频率大于显性基因频率。每穗粒数和千粒质量在2个试点的平均狭义遗传力分别为54.00%和63.17%。
2.2 每穗粒数和千粒质量的杂种优势
由表6可见,尽管每穗粒数和千粒质量在2试点间都出现了中亲、超高亲和超低亲优势为负值的一些组合,但两性状的超低亲优势、平均中亲优势和超高亲优势均为正值,这表明每穗粒数和千粒质量主要表现正向杂种优势,且表现为趋高亲遗传。每穗粒数和千粒质量的3种杂种优势变异幅度都较大,并在组合间、地点间以及基因型与地点互作间均存在极显著差异(表7)。因此,每穗粒数和千粒质量的杂种优势表现受基因型、环境和基因型与环境互作等因素的共同影响。
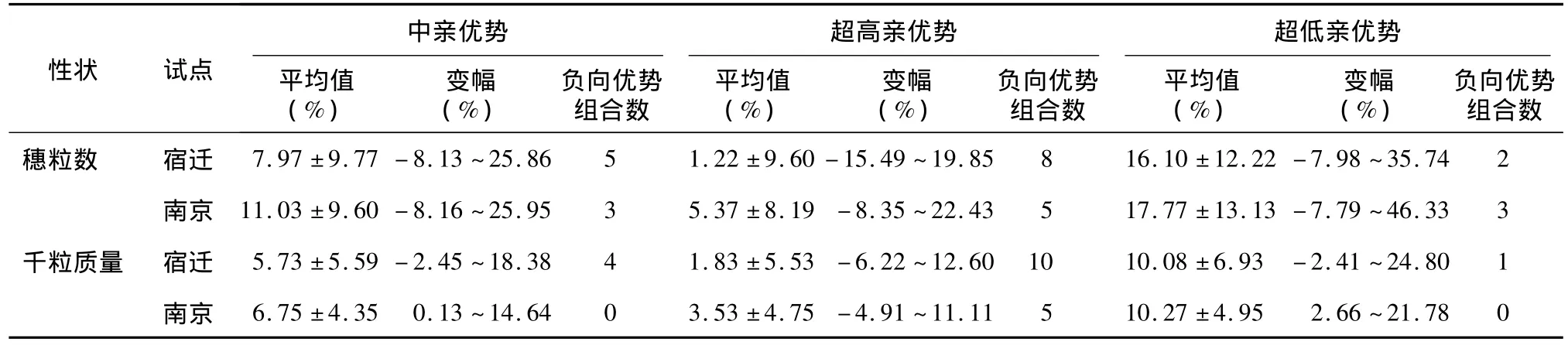
表6 每穗粒数和千粒质量的杂种优势Table 6 Heterosis of kernel number per spike and thousand-grain weight

表7 每穗粒数和千粒质量超高亲优势联合方差分析Table 7 Pooled analysis of variance of super-parent heterosis for kernel number per spike and thousand-grain weight
3 讨论
3.1 关于小麦穗粒数和千粒质量的遗传特点
不少学者对小麦穗粒数和千粒质量的遗传规律、基因作用方式等进行了研究。姚金保等[31]早期的研究结果表明,小麦每穗粒数和千粒质量遗传控制由加性和非加性效应共同起作用,但以加性为主。Inamullah等[8]的研究结果也得到相似的结论。本试验结果表明,小麦每穗粒数和千粒质量受加性效应和显性效应2种遗传效应的共同作用,显性作用相对较大,加性作用相对较小,这与张利华等[11]、Hussain 等[12]、Akram 等[13]的研究结果相符。总之对小麦每穗粒数和千粒质量的遗传研究中,关于加性和显性共同起作用这一观点是基本一致的,但在加性与显性的相对重要性方面尚有分歧。这些分歧可能与试验环境、供试材料的不同有关。
3.2 关于环境对小麦穗粒数和千粒质量的影响
小麦产量性状是复杂的数量性状,除受控于基因型外,还在一定程度上受环境因子以及基因型与环境因子互作等的影响[32-33]。本研究中,有关试验结果在2个地点间存在一定差异,宿迁点的每穗粒数和千粒质量分别比南京点高2.25粒和1.20 g。这种差异主要是由于种植地点的气候条件(温度、光照、降水)、土壤以及栽培措施的不同所引起。Reynolds等[34]认为每穗粒数的减少与二棱期至开花的时间缩短有关。Ishag等[35]研究认为,千粒质量与灌浆期间的平均气温呈负相关,且温度每上升1℃,千粒质量下降4 mg。Borner等[36]检测到2个与每穗粒数相关的QTL以及3个与千粒质量相关的QTL;Heidari等[37]检测到2个控制每穗粒数的主效QTL和6个控制千粒质量的主效QTL,具有基因型-环境互作效应。因此,在小麦高产育种实践中,应在不同生态区对育种材料进行多点鉴定试验,筛选在不同环境条件下表现均稳定的优良基因型。在推广高产小麦品种时,还需要采用配套栽培技术,按照品种的适应范围进行区域化种植和生产。
3.3 关于小麦穗粒数和千粒质量的遗传改良
对产量构成因素杂种优势的分析结果表明,每穗粒数和千粒质量均表现正向杂种优势,且表现为趋高亲遗传,而且优势变幅都较大,这与Ilker等[29]对每穗粒数和Inamullah等[23]对千粒质量的研究结果相似。因此,通过正确选择双亲,利用其正向优势来协调提高穗粒数和千粒质量进而筛选出产量潜力更高的小麦品种是可能的。本研究结果表明,小麦每穗粒数和千粒质量的遗传符合加性-显性模型,基因的显性作用较加性作用重要。高每穗粒数和千粒质量表现高值为显性,低值为隐性。有关研究结果对小麦产量构成因素的遗传改良可能有一定参考价值。在育种早代,由于高每穗粒数和千粒质量基因型含显性基因较多,且处于杂合分离状态,因此必须通过连续多代的定向选择才能获得理想的材料。在本研究中,淮麦33和烟2801是含高穗粒数显性基因较多的2个亲本,淮麦20是含高千粒质量显性基因较多的亲本,这3个亲本其他农艺性状和抗病性表现也较好,在小麦高产育种中,应进一步利用这3个优良亲本。
[1] 吴宏亚,蒋正宁,王摇玲,等.小麦籽粒硬度及其对面粉加工品质影响的研究进展[J].江苏农业学报,2014,30(2):437-441.
[2] 蒋小忠,封超年,郭文善.磷肥种类对弱筋小麦籽粒产量和蛋白质含量的影响[J].江苏农业科学,2013,41(12):63-66.
[3] 蔡金华,陈爱大,李东升.不同施氮条件下镇麦168籽粒产量与构成因素的相关性研究[J].江苏农业科学,2014,42(10):73-75.
[4] HE Z H,RAJARAM S,XIN Z Y,et al.A history of wheat breeding in China[M].Mexico DF:CIMMYT,2001.
[5] 田纪春,王延训.超级小麦的育种策略与实践[J].作物杂志,2005(4):67-68.
[6] TAHIR M S,ALAM K,CHOWDHRY M A,et al.Genetic analy-sis of some important economic traits in bread wheat(Triticum aestivum L.)[J].Pakistan Journal of Agricultural Sciences,1995,32:172-177.
[7] KHAN A S,KHAN M K R,KHAN T M.Genetic analysis of plant height,grain yield and other traits in wheat(Triticum aestivum L.)[J].International Journal of Agriculture&Biology,2000,2:129-132.
[8] INAMULLAH,AHMAD H,MOHAMMAD F,et al.Diallel analysis of the inheritance pattern of agronomic traits of bread wheat[J].Pakistan Journal of Botany,2006,38(4):1169-1175.
[9] NAZEER W,ALI Z,ALI A,et al.Genetic behavior for some polygenic yield contributing traits in wheat(Triticum aestivum L.)[J].Journal of Agricultural Research,2010,48(3):267-276.
[10] CHEN D,ZHANG J P,WANG J S,et al.Inheritance and availability of high grain number per spike in two wheat germplasm lines[J].Journal of Integrative Agriculture,2012,11(9):1409-1416.
[11]张利华,许梅芬.小麦收获指数和其它几个农艺性状的基因效应分析[J].核农学报,1997,11(3):135-140.
[12] HUSSAIN F,SIAL R A,ASHRAF M.Genetic studies for yield and yield related traits in wheat under leaf rust attack[J].International Journal of Agriculture&Biology,2008,10:531-535.
[13] AKRAM Z,AJMAL S U,SHABBIR G,et al.Inheritance mechanism of some yield components in bread wheat Journal[J].Pakistan Journal of Agricultural Research,2009,22:1-8.
[14] RABBANI G,MAHMOOD A,SHABBIR G,et al.Gene action in some yield attributes of bread wheat under two water regimes[J].Pakistan Journal of Botany,2011,43(2):1141-1156.
[15] RAHMAN M A,SIDDQUIE N A,ALAM M R,et al.Genetic analysis of some yield contributing and quality characters in spring wheat(Triticum aestivum)[J].Asian Journal of Plant Sciences,2003,2(3):277-282.
[16]姚金保,任丽娟,张平平,等.小麦产量构成因素的双列杂交分析[J].核农学报,2011,25(4):633-638.
[17] AHMAD F,KHAN S,AHMAD S Q,et al.Genetic analysis of some quantitative traits in bread wheat across environments[J].African Journal of Agricultural Research,2011,6(3):686-692.
[18]李斯深,陈茂学,王洪刚.利用重组自交系(RILs)群体进行质量-数量性状的遗传分析—遗传模型和小麦产量性状遗传[J].作物学报,2001,27(6):896-904.
[19] RASUL I,KHAN A S,ALI Z.Estimation of heterosis for yield and some yield components in bread wheat[J].International Journal of Agriculture&Biology,2002,4:214-216.
[20]冀天会,张灿军,杨子光,等.水旱生态型冬小麦品种杂种优势分析[J].麦类作物学,2003,23(4):19-22.
[21] BAO Y G,WANG S,WANG X Q,et al.Heterosis and combining ability for major yield traits of a new germplasm Shannong 0095 derived from Thinopyrum intermedium[J].Agricultural Sciences in China,2009,8(6):753-760.
[22]王玉斌,逯腊虎,武计萍,等.冬小麦主要农艺性状的杂种优势和配合力分析[J].山西农业科学,2014,42(7):653-656.
[23] INAMULLAH,AHMAD H,MOHAMMAD F,et al.Evaluation of the heterotic and heterobeltiotic potential of wheat genotypes for improved yield[J].Pakistan Journal of Botany,2006,38(4):1159-1167.
[24] AL-FALAHY M A H.Estimate of genetic parameters of grain yield and some agronomic traits in durum wheat using diallel crosses[J].Journal of Agricultural Science and Technology,2013,3:26-34.
[25]许永财,相吉山.春小麦主要农艺性状杂种优势与配合力研究[J]. 青海大学学报:自然科学版,2007,25(2):4-9.
[26]赵 鹏,彭惠茹,倪中福,等.穗分枝小麦与普通小麦杂交组合主要性状的杂种优势与配合力分析[J].麦类作物学报,2009,29(2):212-216.
[27]史秀秀,毕晓静,马守才,等.黄淮麦区杂交小麦亲本的杂种优势和配合力分析[J].麦类作物学报,2013,33(6):1111-1118.
[28] ABDULLAH G M,KHAN A S,ALI Z.Heterosis study of certain important traits in wheat[J].International Journal of Agriculture& Biology,2002,4(3):326-328.
[29] ILKER E,TONK F A,TOSUN M.Heterosis for yield and its components in bread wheat crosses among powdery mildew resistant and susceptible genotypes[J].Pakistan Journal of Botany,2010,42(1):513-522.
[30]刘来福,毛盛贤,黄远樟.作物数量遗传[M].北京:农业出版社,1984:125-149.
[31]姚金保,王书文,姚国才,等.小麦产量构成因素的遗传分析[J].上海农业学报,2004,20(1):45-48.
[32]常 磊,柴守玺,杨德龙,等.我国旱地春小麦产量及主要农艺指标的变异分析[J].应用生态学报,2010,21(11):2821-2829.
[33] SANCHEZ-GARCIAA M,ALVAROA F,MARTIN-SANCHEZA J A,et al.Breeding effects on the genotype×environment interaction for yield of bread wheat grown in Spain during the 20th century[J].Field Crops Research,2012,126:79-86.
[34] REYNOLDS M P,CALDERINI D F,CONDON A G,et al.Physiological basis of yield gains in wheat associated with the LR19 translocation from Agropyron elongatum[J].Euphytica,2001,119:139-144.
[35] ISHAG H M,MOHAMED B A.Phasic development of spring wheat and stability of yield and its components in hot environments[J].Field Crops Research,1996,46:169-176.
[36] BORNER A,SCHUMANN E,FURSTE A,et al.Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat(Triticum aestivum L.)[J].Theoretical and Applied Genetics,2002,105:921-936.
[37] HEIDARI B,SAYED-TABATABAEI B E,SAEIDI G,et al.Mapping QTL for grain yield,yield components,and spike features in a doubled haploid population of bread wheat[J].Genome,2011,54:517-527.
