人Grn E单克隆抗体的制备和鉴定
2014-12-31李卫玲张娅玲李彦青张丽娟夏海滨
李卫玲,张娅玲,李彦青,张丽娟,夏海滨
(陕西师范大学 生命科学学院,秦巴山区可持续发展协同创新中心,陕西 西安 710119)
颗粒蛋白前体(Progranulin,PGRN)是一种具有多种生物学功能的生长因子,除参与调节生长发育[1]、组织损伤修复和抗炎症等生理过程外[2],还与肿瘤及糖尿病的发生有着密切的关系[3-4].近年来研究表明,PGRN基因突变引起的单倍剂量不足(表达或分泌降低)是导致具有泛素阳性包涵体特征的额颞叶变性发生的主要因素之一[5].PGRN在结构上包含七个半结构相似的结构域,以P—G—F—B—A—C—D—E的顺序排列,其中P为半个结构域.每个结构域都是由四个β-发夹结构以旋转的梯状结构堆积而成,含有12个半胱氨酸并形成六对二硫键,这些结构域之间以短的多肽连接,弹性蛋白酶在连接处可将颗粒蛋白前体水解为大小不等的片段,这些片段很可能具有不同的生物学功能[6].目前对PGRN研究大多都集中于其生物学功能,对PGRN的各结构域功能及其分子机制研究较少.
Sortilin分子是调节细胞外PGRN水平的主要决定因素.Sortilin N-末端β螺旋结构域可与PGRN C-末端Grn E结合,介导PGRN的内化及运输[7].TDP-43可通过上调Sortilin的表达来促进PGRN吸收,从而减少细胞外PGRN水平[8].PGRN能够促进神经细胞存活[9],有研究表明PGRN的神经营养因子功能是通过 Grn E 结构域介导的[10],但PGRN在发挥神经营养因子功能方面并不依赖膜受体Sortilin[11].
单克隆抗体在研究蛋白质的结构和功能方面有重要的作用.目前还没有针对人Grn E单抗的报道,为此,本文将原核表达GST-Grn E融合蛋白作为免疫原免疫Balb/c小鼠,经过融合及初步筛选,成功获得5株针对Grn E的单克隆抗体细胞株,并对此进行初步鉴定.针对人Grn E单克隆抗体的成功制备为探索PGRN及Grn E的生物学功能及其分子机制奠定重要基础.
1 材料与方法
1.1 细胞株、菌株及载体
SP2/0和 HEK293 购自美国 ATCC,E.coli DH5α及BL21(DE3)菌株 均由本实验室保存.pGEM-T easy克隆载体购自Promega公司;pRSET-B原核表达载体购自Invitrogen公司;pGEX-4T-3原核表达载体购自Amersham公司;pOTB7/hPGRN质粒购自美国ATCC公司.4~6周龄Balb/c雌性小鼠购自第四军医大学.
1.2 主要试剂
改良型RPMI-1640干粉购自Hyclone公司;新生牛血清购自杭州四季青生物工程材料有限公司;8AG、PEG3350、IL-6、石 蜡 油 购 自 Sigma;100×HT、50×HAT购自 Gibco;BSA 购自 Hyclone;ABTS购自Gibco;Ni-NTA柱购自Novagen公司;限制性内切酶Prime STAR,dNTP mix,Solution I购自日本TaKaRa公司;小提中量试剂盒购自TIANGEN公司;DNA Marker购自 Hybigen公司;Anti-His antibody购自天根生物技术公司;辣根酶标记山羊抗小鼠IgG二抗购自中杉金桥公司;ECL免疫印迹底物购自美国Thermo公司;DAPI购自美国Invitrogen;小鼠单克隆抗体亚型鉴定Elisa kit购自Sigma公司.
1.3 Grn E蛋白的表达
1.3.1 人Grn E片段基因的克隆 Grn E位于人PGRN基因的1555~1782bp,共228bp.以实验室保存的hPGRN基因为模板,设计Grn E引物,5’-ClaI for:ATCGATgtggagtgtggggaaggaca,3’-SpeI no stop back:ACTAGTcagcagctgtctcaaggctg.所用引物由北京华大基因公司合成.
PCR产物连接pGEMT-easy载体,转化至DH5α感受态细菌,小提质粒经EcoRI酶切初步鉴定为阳性克隆后送华大基因公司进行测序,正确的克隆命名为pGEM-T easy/Grn E,并将之用于后续实验.
1.3.2 人Grn E原核表达载体的构建及表达纯化pGEM-T easy/Grn E由ClaI和SpeI双切后,回收片段.分别连入6His-tag原核表达载体pRSET-B和 GST-tag原核表达载体pGEX-4T-3中,经 KpnI酶单切鉴定切不动的为疑似阳性质粒.然后再由ClaI和SpeI双酶切进一步验证,切出300bp条带的阳性克隆分别命名为pRSET-B/Grn E和pGEX-4T-3/Grn E,并将之用于后续原核表达.
质粒电转化至大肠杆菌BL21(DE3),经终浓度1mmol/L IPTG诱导(37℃,225r/min,4h)后,收集菌体,低温超声裂解,以10%SDS-PAGE检测目的蛋白的表达.当所构建的His-Grn E和GSTGrn E蛋白表达良好且分子量与预期大小一致时,即进行大量表达纯化.分别挑取表达量较好的pRSET-B/Grn E 克隆4和pGEX-4T-3/Grn E 克隆2菌液划板后挑取单克隆于200mL培养基,至OD600nm为0.6,IPTG诱导表达,以8mmol/L尿素变性方法纯化过柱,分别以20mmol/L、40 mmol/L、60mmol/L和80mmol/L咪唑洗脱杂蛋白,最后以250mmol/L咪唑洗脱目的蛋白.
1.4 单克隆抗体的制备
1.4.1 免疫动物 免疫4只6周龄Balb/c雌性小鼠,GST-Grn E原核蛋白免疫量为40μg/只.将200 μg免疫原稀释于2mL经生理盐水中,然后加等体积褔氏完全佐剂充分研磨,皮下多点注射.间隔3周,第二次免疫生理盐水稀释后加褔氏不完全佐剂研磨.间隔3周后,第三次免疫生理盐水稀释至80 μg/mL腹腔注射每只0.5mL,2周后断尾取血测定免疫效价.选取效价好的小鼠加强免疫,并用于后续实验.
1.4.2 骨髓瘤细胞和饲养细胞的准备 复苏SP2/0细胞,8AG终浓度20μg/mL筛选稳定后扩大培养,细胞总数约为5×107.铺板2只正常小鼠腹腔巨噬细胞作为饲养层细胞.
麻醉脱颈处死4号免疫鼠后,75%酒精消毒5 min,制备脾细胞总数约为2×108,调整骨髓瘤细胞和脾细胞的比例为1∶5.取37℃预热融合剂1 mL,30s内加完,室温静置90s.加终止液无血清1640培养基,9min内加完25mL.800r/min,4 ℃离心6min.重悬至42mL铺于饲养细胞的96孔板.
1.4.3 杂交瘤细胞筛选和克隆培养 融合24h后开始HAT/HT选择性培养液筛选.7~10d后当杂交瘤细胞长至合适大小时补充换液,采用Elisa方法检测细胞上清中的抗体,将阳性培养孔中的杂交瘤细胞株通过限稀释法,以每孔分别含0.5、1和2个细胞接种于96孔培养板中.经过3~4次克隆化检测为全阳性后,选择抗体效价高、呈单个克隆生长、形态良好的细胞孔,进行转孔扩大培养和冻存.
1.4.4 抗体腹水的制备 从筛选得到的5株针对Grn E阳性杂交瘤细胞株中选取效价较好的3株制备腹水.选取6周龄Balb/c雌性小鼠12只,腹腔注射石蜡油0.8mL/只.7~14d后,1×PBS重悬的杂交瘤细胞调整细胞浓度2×106细胞/mL,腹腔注射0.5mL/只.1周左右收获含有抗体的腹水,分装冻存,用于后续实验.
1.5 针对Grn E单克隆抗体的鉴定
1.5.1 Western blot 变性后的样品经10%SDSPAGE凝胶电泳后,半干转至PVDF膜,5%脱脂牛奶封闭2h,一抗室温孵育1h,再经山羊抗小鼠IgG HRP标记的二抗(1∶10 000)室温孵育1h,ECL发光,暗室显影.
1.5.2 免疫荧光染色 细胞经4%甲醇室温固定15min,1×PBS洗涤后再由0.5%Triton X-100通透20min,一抗室温孵育2h,再经山羊抗小鼠IgG TRITC标记的二抗(1∶100)室温1h,DAPI(1∶3 000)染色10min,荧光倒置显微镜下观察照相.
1.5.3 免疫沉淀 将携带PGRN的真核表达载体转染生长于一个60mm培养皿的HEK293细胞中,48h后制备获得500μL细胞裂解液,其中取50 μL细胞裂解液加入等体积的2×SDS,变性后取20 μL作为input用于western blot检测;分别取ProteinA/G珠子20μL和抗体2μL于500μL 3%BSA中,4℃,10r/min封闭结合1h,经完全洗涤后,将剩余的450μL细胞裂解液全部加入上述珠子继续结合1h,珠子经1×PBS洗涤后,加入等体积约20μL的2×SDS,变性后全部上样用于western blot检测.
2 结果
2.1 His-Grn E及GST-Grn E原核表达表达和纯化
将pRSET-B/Grn E酶切鉴定正确的克隆电转至BLD21(DE3)菌株,随机挑取4个克隆,经IPTG诱导后,SDS-PAGE电泳发现,在相对分子质量(Mr)为17kD处与未诱导相比有较强特异性条带(图1a).细菌裂解液经Ni-NTA亲和层析柱纯化后,采用anti-His抗体经western blot检测,17kD处有His-Grn E目的条带 (图1b),表明所诱导表达的蛋白正确.Nano drop OD280nm吸光值测浓度分别为1.06mg/mL.

图1 pRSET-B/Grn E的原核表达纯化Fig.1 The prokaryotic expression and purification of pRSET-B/Grn E
将pGEX-4T-3/Grn E电转至BLD21(DE3)菌株,经IPTG诱导后,SDS-PAGE电泳发现,在相对分子质量为34kD处,细菌裂解液中有1条特异性条带,而未诱导处相应蛋白基本无表达(图2a).细菌裂解液经Ni-NTA亲和层析柱纯化后,western blot检测,34kD处有 GST-Grn E目的条带(图2b),测定浓度为0.43mg/mL.

图2 pGEX-4T-3/Grn E重组蛋白原核表达与纯化Fig.2 The expression and purification of pGEX-4T-3/Grn E
2.2Grn E单克隆抗体的制备
免疫小鼠断尾取血,经Elisa检测选取效价达到1∶128 000的4号免疫鼠,取脾脏经杂交瘤融合技术获取杂交瘤细胞株,通过3次有限稀释法克隆化和Elisa检测均全阳性的有5株,分别是1D5,1G5,2E1,3F7和4D4.选取其中效价较高的3株1D5、2E1和3F7扩大培养制备腹水,以备后期检测用.通过使用Sigma小鼠单克隆抗体亚型鉴定Elisa kit,测定1D5和3F7均为IgG1型,2E1为IgG2a(图3).

图3 杂交瘤分泌的抗体亚型测定Fig.3 The detection of the subtype of antibody from hybridoma cell lines
2.3 人Grn E单克隆抗的鉴定
2.3.1 Western blot鉴定 分别采用磷酸钙法转染将携带PGRN和Grn E的真核表达载体转染至HEK293细胞,72h裂解细胞收取蛋白,然后通过western blot鉴定所制备抗体.结果显示所制备的针对人Grn E的3株抗体(1D5、2E1及3F7)在稀释1∶1 000时能检测到与目的片段大小一致的特异条带(见图4).

图4 针对Grn E单克隆抗体的western blot鉴定Fig.4 The diagnosis of anti-Grn E monoclonal antibody by western blot
2.3.2 免疫荧光染色鉴定 为了检测所制备的单克隆抗体是否可以用于免疫荧光染色,将携带外源人PGRN和Grn E基因的真核表达载体分别转染至HEK293细胞,24h后4%甲醛固定细胞,经免疫荧光染色检测所制备的抗体.放大倍数为目镜是10X,物镜20X.阴性对照为anti-GST tag.结果表明,3株抗体可以检测出PGRN和Grn E的表达(见图5).
2.4 Grn E抗体用于免疫沉淀的鉴定
抗体在免疫沉淀实验过程具有重要的应用价值,为了验证所制备的针对人Grn E单克隆是否可以用于免疫沉淀实验,我们选取腹水效价较好的1D5和3F7两株单克隆抗体先与Protein A/G珠子进行结合,经过充分洗涤后再与转染有外源PGRN基因的细胞裂解液孵育,最后用于免疫印迹的抗体选用实验室以往制备的针对PGRN中Grn A结构域的单克隆抗体[12].免疫印迹所获得分子量与PGRN分子量相一致,该结果表明,两株针对人Grn E抗体可用均于免疫沉淀实验(见图6).

图5 针对人Grn E单克隆抗体的细胞免疫荧光染色鉴定Fig.5 Immunofluorescence staining of monoclonal antibodies against Grn E in HEK293
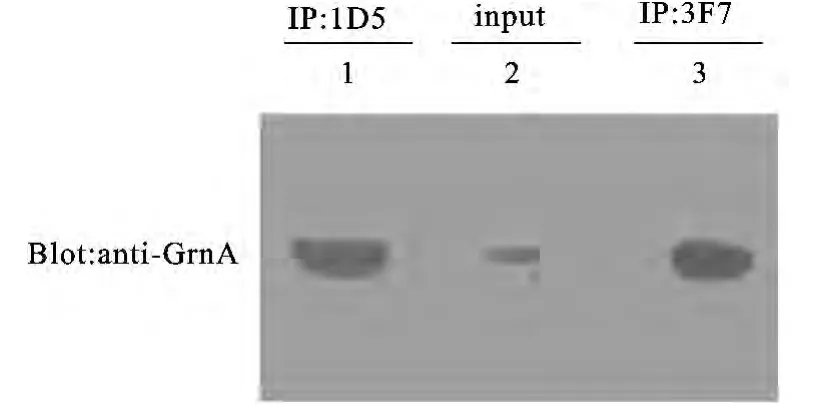
图6 Grn E单克隆抗体用于免疫沉淀淀检测Fig.6 The detection of anti-Grn E by immunoprecipitation assay
3 讨论
PGRN及其水解产生的Grn多肽均具有一定的生物学功能,在它们的功能研究中,抗体是一种有利的、必不可少的工具.针对PGRN的结构特点,制备能够用于识别不同结构域的抗体,对于PGRN的功能研究和临床应用有重要意义.
本实验室曾经通过纯化的真核表达的PGRN-免疫Balb/c小鼠,仅获得针对Grn A、Grn B和Grn C三个结构域的单克隆抗体[12].鉴于Grn E结构域的重要性,为了获得针对人Grn E的单克隆抗体,我们通过采用原核载体单独表达Grn E结构域,并将之用于制备针对该结构域的单克隆抗体.经过Elisa初步筛选获得5株阳性杂交瘤细胞株,其中1G5和4D4效价相对较低,因此选取另外3株:1D5、2E1和3F7制备腹水和测定亚型,三株抗体的亚型均为IgG型.在后期western blot和免疫荧光染色鉴定中,1D5和3F7显示出良好的特异性及敏感性,其中2E1效果相对较差,可能与腹水质量不高有关;或者与杂交瘤细胞培养过程中出现染色体丢失而丧失抗体分泌能力有关.因此,后续可通过复苏该细胞株再次克隆化并制备腹水进行进一步的检测.此外,经免疫沉淀结果验证1D5和3F7可用于后续与PGRN相互作用蛋白的实验研究.
4 结论
本文制备并获得3株针对人Grn E的单克隆抗体.经ELISA测定证实1D5和3F7亚型均为IgG1型,2E1为IgG2a;所制备的针对Grn E的单克隆抗体均可以通过western blot和免疫荧光染色;通过免疫沉淀实验表明,所制备的人Grn E抗体可用于后续与PGRN相互作用蛋白的研究.针对人Grn E单抗的成功制备,为PGRN和Grn E功能研究及其分子机制研究奠定了重要基础.
[1]Zanocco-Marani T,Bateman A,Romano G,et al.Biological activities and signaling pathways of the granulin/epithelin precursor[J].Cancer Research,1999,59:5331-5340.
[2]Zhu Jing,Nathan C,Jin Wenwen,et al.Conversion of proepithelin to epithelins:roles of SLPI and elastase in host defense and wound repair[J].Cell,2002,111:867-878.
[3]Matsubara T,Mita A,Minami K,et al.PGRN is a key adipokine mediating high fat diet-induced insulin resistance and obesity through IL-6in adipose tissue[J].Cell Metabolism,2012,15:38-50.
[4]Bateman A,Bennett H P.The granulin gene family:from cancer to dementia[J].Bioessays,2009,31:1245-1254.
[5]Baker M,Mackenzie I R,Pickering-Brown S M,et al.Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17[J].Nature,2006,442:916-919.
[6]Bateman A,Bennett H P.Granulins:the structure and function of an emerging family of growth factors[J].Journal of Endocrinology,1998,158:145-151.
[7]Zheng Yanqiu,Brady O,Meng P,et al.C-terminus of progranulin interacts with the beta-propeller region of sortilin to regulate progranulin trafficking[J].PLOS ONE,2011,6:1-7.
[8]Prudencio M,Jansen-Westa K,Lee W,et al.Misregulation of human sortilin splicing leads to the generation of a nonfunctional progranulin receptor[J].Proceedings of the National Academy of Sciences of the United States of America,2012,109:21510-21515.
[9]Damme P,Hoecke A,Lambrechts D,et al.Progranulin functions as a neurotrophic factor to regulate neurite outgrowth and enhance neuronal survival[J].The Journal of Cell Biology,2008,181:37-41.
[10]Muynck L,Herdewyn S,Beel S,et al.The neurotrophic properties of progranulin depend on the granulin E domain but do not require sortilin binding[J].Neurobiology of Aging,2013,34:2541-2547.
[11]Gass J,Lee W,Cook C,et al.Progranulin regulates neuronal outgrowth independent of sortilin[J].Molecular Neurodegeneration,2012,733:1-13.
[12]Chai Lihong,Mao Qinwen,Liu Shihai,et al.Domainspecific monoclonal antibodies produced aganist human PGRN[J].Hybrodoma,2011,30:271-278.
